
Abstract
Abstract
Porcine pluripotent cells with the capacity to generate germ line chimeras have not been developed yet. The transcription factor Oct-4 is an important marker of undifferentiating status and a central regulator of pluripotency in cells. Establishment of an Oct-4 promoter-based reporter system, such as that used in mice, will be a useful tool for monitoring the differentiating statuses of porcine cells both in vivo and in vitro. In the present study, we constructed a vector, pOGN2, in which enhanced green fluorescent protein (EGFP) was driven by the porcine Oct-4 promoter. In pigs containing this vector, EGFP was expected to be specifically expressed in pluripotent cells. We delivered the vectors into porcine fetal fibroblasts (PEFs) using liposomes. After transfected PEFs were selected with G418, we established eight cell lines containing the pOGN2 vector. When transgenic cells were used as donor nuclei to make somatic cell nuclear transfer (SCNT) embryos, SCNT embryos derived from four transgenic cell lines expressed green fluorescence. When PEFs with pOGN2 vectors were infected with retroviral vectors encoding the four transcription factors (Oct-4, Sox2, Klf4, and c-Myc), EGFP-expressing iPS cell colonies were observed at day 20. This work lays a foundation that can be used to generate a pig strain with an Oct4-EGFP reporter system, which would be greatly helpful in studying the differentiating and reprogramming mechanisms of pig embryos.
Introduction
Oct-4, a member of Class V of the POU (Pit-Oct-Unc) transcription factor family, plays a crucial role in the establishment and maintenance of transcriptional regulation during early embryo development and cell differentiation (Kellner and Kikyo, 2010). The expression of Oct-4 is specifically found in pluripotent cells, including inner cell mass (ICM) cells, embryonic stem (ES) cells, germ cells, and embryonic
To achieve above goals, in the present study, we constructed a vector, pOGN2, in which enhanced green fluorescent protein (EGFP) was driven by the porcine Oct-4 promoter. Then we established fibroblast cell lines containing pOGN2 vectors. To verify if the undifferentiated cells reprogrammed from the fibroblast cell lines containing pOGN2 vectors could express EGFP, the transgenic cells were either used as donor nuclei to make somatic cell nuclear transfer (SCNT) embryos or induced with the four transcription factors (Oct-4, Sox2, Klf4, and c-Myc). We found that EGFP was expressed in both cloned embryos and iPS cell colonies.
Materials and Methods
Isolation of porcine Oct-4 promoter and construction of pOGN2 vector
For cloning 3.2-kb porcine Oct-4 promoter, two primers were designed, namely, poct-Hind-F: CCCaagcttGGATCCCTAGCCTGGAAACCT and poct-Kpn-R: CGGggtaccGGGGAAGGAAGGCGCCCCAAG. The genomic DNA was extracted as a template from the fetal fibroblast cells of Tibetan miniature pigs with a cell genomic DNA extraction kit from Takara. Long and accurate (LA) PCR was performed with the following cycling conditions: 4 min at 95°C, 30 cycles of 95°C for 30 sec, 68°C for 30 sec, 72°C for 3 min, and finally 72°C for 5 min. The amplified porcine Oct-4 promoter DNA was subcloned to the pMD18T vector (Takara, Tokyo, Japan) to create pT-Oct-pro for sequencing.
The porcine Oct-4 promoter was isolated by digestion of pT-Oct-pro with Hind III and Kpn I and inserted upstream of EGFP cDNA in pEGFP-N2 (BD Biosciences Clontech, Palo Alto, CA) to produce pCOGN2. Then the CMV promoter in pCOGN2 was removed by digestion with Ase I and Bgl II. The DNA fragment with 3′- and 5′-protruding ends produced by Ase I and Bgl II digestion was blunt and self-ligated with a DNA blunting kit from Takara to create the vector pOGN2 (Supplementary Fig. S1; see online supplementary data at www.liebertonline.cell/com).
pOGN2 was linearized by Hind III before being used for cell transfection.
Expression assay of porcine Oct-4 promoter in pOGN2
Pig iPS colonies (Esteban et al., 2009b) were seeded onto 24-well dishes coated with a layer of mitotically inactivated mouse feeder cells at a density of 2 × 103 cells/well 1 day before transfection and grown in KSR medium supplemented with 15% KnockOut serum replacer (Gibco, Grand Island, NY) at 39°C in an atmosphere of 5% CO2 in air. For transfection, 1.4 μg of linearized pOGN2 or pEGFP-N2 was mixed with 4 μL Lipofectamine 2000 (Invitrogen, Carlsbad, CA) in Opti-MEM I Medium (Invitrogen). The complexes were added to the cell culture medium and incubated for 6 h at 39°C. One day after transfection, cells were observed for EGFP expression by fluorescence microscopy.
Establishment of Oct4–EGFP transgenic PEF cell lines
Porcine fetal fibroblasts (PEFs) were isolated from a day 35 Tibetan miniature pig fetus. After removal of the head, viscera, and extremities, the fetuses were thoroughly rinsed three times with phosphate-buffered saline (PBS). Then the tissues were minced into 1 mm3 pieces using a scalpel blade and digested for 4 h with collagenase (200 unites/mL, Gibco) at 39°C. The cell pellets were aspirated with a pipette, centrifuged at 700 rpm for 10 min, and resuspended in PEF culture medium (DMEM high glucose supplemented with 2 mM L-glutamine, 0.1 mM Na-pyruvate, penicillin/streptomycin (Gibco), and 15% fetal bovine serum (FBS) (Hyclone, Salt Lake City, UT). After cells reached confluence, they were cryopreserved in liquid nitrogen.
After passaging 1–2 generations, 4 × 104 PEFs were seeded into one well of a six-well plate 1 day before transfection. For transfection, 4 μg of Hind III-linearized pOGN2 and 12 μL Lipofectamine 2000 were mixed together. The transfection was performed according to the manufacturer's protocol. Media were changed after 6 h of incubation. One day after transfection, cells were placed in three 100-mm dishes. At 2 days of culture, selective media with 1000 μg/mL G418 (Geneticin; Invitrogen, Carlsbad, CA) were added, and cells were cultured for an additional 8–10 days. G418-resistant cell colonies were picked with the cloning ring (Sigma, St. Louis, MO) and propagated in a 48-well culture plate. Four to 6 days later, wells in which cells had grown confluently were trypsinized (0.05% trypsin) and seeded into a 24-well plate to be frozen soon after. Part of each cell colony was used for PCR screening. Cells were resuspended in lysis buffer (0.121 g Tris-base, 0.4 mg proteinase K, 0.09 mL Triton X-100, to 10-mL final volume with ddH2O) for 1 h at 37°C, 1 h at 55°C, and 95°C for 10 min (to inactivate proteinase K), and 1 μL lysate was used as template in a 20 μL PCR reaction. The PCR conditions were as follows: 4 min at 95°C, 30 cycles of 95°C for 30 sec, 65°C for 30 sec, and 72°C for 30 sec, and then 72°C for 5 min.
DNA fragments (300 bp) were expected with the primers egfp-p1 (5′-CTCGTGACCACCCTGACCTA-3′) and egfp-p2 (5′-CTTGATGCCGTTCTTCTGCT-3′) from cells integrated with pOGN2. The PCR products were sequenced to confirm their authenticity.
Production of SCNT embryos with Oct4–EGFP transgenic PEFs
Stably transfected PEF cell lines were used as donor cells for SCNT. Porcine SCNT was performed as described by Lai and Prather (2004). Briefly, ovaries were obtained from a local slaughterhouse and transported to the laboratory in saline with penicillin/streptomycin. After cumulus–oocyte complexes (COCs) were aspirated from the antral follicles, COCs were cultured in maturation medium(Lai and Prather, 2004) for 42–44 h at 39°C. After oocyte maturation, cumulus cells were removed and metaphase II oocytes were enucleated by removal of the polar body and the associated metaphase plate. The cycling cells were used as donor nuclei. The donor cells were inserted into the perivitelline space of each enucleated oocyte using the micropippette. Then the reconstructed embryos were transferred into an electrical fusion chamber containing fusion medium(Lai and Prather, 2004) for fusion and activation with two direct current pulses (1.2 kV/cm, 60 μsec). The fused reconstructed embryos were transferred to 500 μL PZM-3 medium (Lai and Prather, 2004) in a four-well dish covered with light mineral oil and cultured in incubator (100% humidity, 39°C, 5% CO2 in air). Expression of EGFP in the embryo was assessed 5–7 days after activation by fluorescence microscopy (Olympus, Tokyo, Japan). EGFP-positive embryos were then subjected to an immunofluorescence test to detect endogenous Oct-4 expression. Day 5–7 parthenogenetic embryos were used as controls.
Immunofluorescence test of SCNT embryos
The immunofluorescence test was carried out as described by Miyoshi et al. (2009), with some modifications. First, SCNT embryos were fixed with PBS containing 4% (v:v) paraformaldehyde for 1 h at room temperature, washed three times with PBS containing 1% FBS (washing buffer), then incubated for 40–60 min in blocking buffer (PBS, 10% FBS, and 0.1% Triton X-100) and transferred into a drop of washing buffer containing 5 μg/mL goat anti-Oct3/4 antibodies (Santa Cruz Biotechnology, Santa Cruz, CA). The drop was covered with paraffin oil to avoid evaporation of the solution. The reaction was performed overnight at 4°C. The embryos were washed three times with washing buffer and subsequently immersed in a drop of washing buffer containing 4 μg/mL Alexa Fluor 594-labeled rabbit antigoat IgG antibodies (AbChemical, Shanghai, China) overnight at 4°C. After being washed three times, embryos were incubated with 10 μg/mL Hoechst33342 (Sigma) for 10 min at room temperature. Finally, embryos were washed three times, loaded on a slide, and covered with a cover slip for fluorescence observation.
Isolation of porcine iPS cells
Retroviral vectors encoding human Oct-4, Sox2, Klf4, and c-Myc were produced using HEK293T cells by plasmid transfection of pMXs–Oct4, pMXs–Sox2, pMXs–Klf4, and pMXs–c-Myc (purchased from Addgene, Cambridge, MA). PEF cells (4 × 104) with pOGN2 vectors per well in a six-well dish were infected with a 2.0-mL unconcentrated retroviral vector in the presence of 8 mg/mL Polybrene (Sigma). Two days after infection, the cells were trypsinized, and 1 × 104 cells were transferred onto feeders in a 10-cm dish containing dFBS medium (DMEM high glucose with antibiotics, glutamine, pyruvate, nonessential amino acids, β-mercaptoethanol, bFGF, and 10% defined FBS) (Hyclone, Salt Lake City, UT). Fluorescence of ES-like colonies were monitored by fluorescence microscopy and picked up for subculture at days 17–22.
Results
Isolation of the Oct-4 promoter from Tibetan miniature pig cells
The genome sequence of the porcine Oct-4 gene and its flanking sequence have been reported; however, the data was derived from domestic pigs. We isolated a 3.2-kb DNA fragment spanning the Oct-4 gene promoter of a Tibetan miniature pig. The sequence of the promoter is 97% homologous to NCBI data (NW_001886435), suggesting subtle differences among different porcine strains.
The Oct-4 promoter region contains three main elements: the distal enhancer (DE), the proximal enhancer (PE), and the core promoter. After alignment of Oct-4 promoter among pigs, mice, and humans, highly conserved DNA sequence in the DE and core promoter regions but low homology in the PE region were found (Fig. 1).
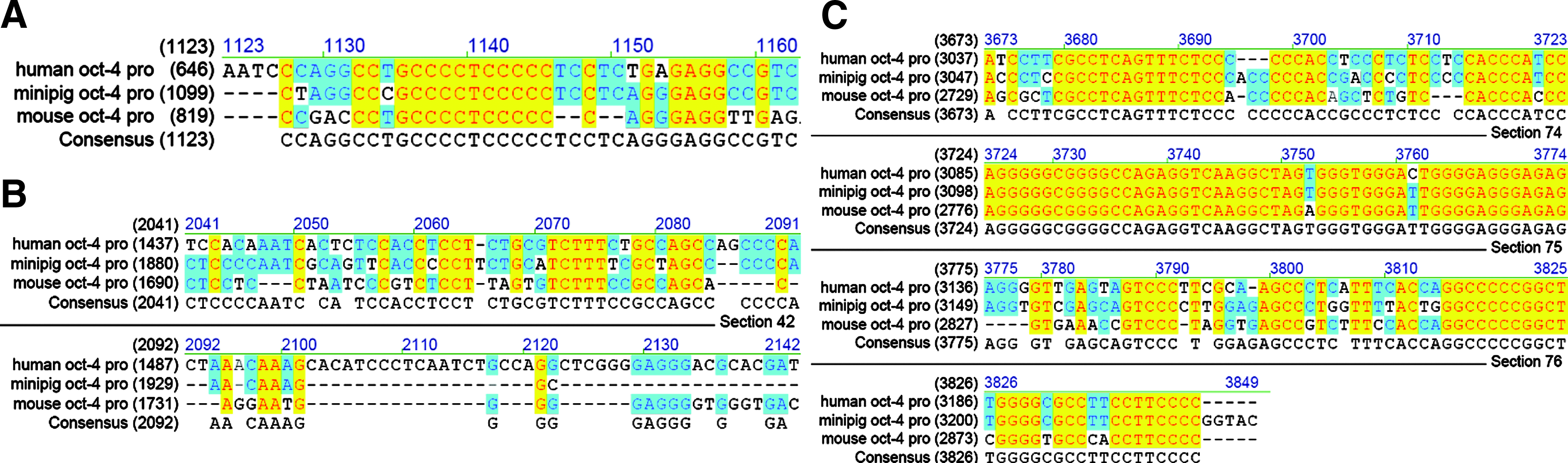
Alignment of human, pig, and mice Oct-4 promoter. (
Transient expression assay
Preliminary experiments were performed to evaluate the specificity of the porcine Oct-4 promoter in pluripotent cells. Porcine iPS-like cells established in our lab, in which the endogenous Oct-4 was activated, were used as test cells. The vector pEGFP-N2, in which EGFP expresseion was driven by ubiquitous CMV promoter, was used as a control. Twenty-four hours after transfection, EGFP expression was observed in porcine iPS cell colonies but not in differentiated feeder cells. In contrast, EGFP was expressed in both feeder cells and iPS cells in the cells transfected with pEGFP-N2. These data show that pOGN2 is a useful marker specific for undifferentiated Oct-4-expressing cells (Supplementary Fig. S2).
Establishment of Oct4–EGFP transgenic PEFs
The reporter construct pOGN2 was linearized and transfected into PEFs. Transfection was performed using Lipofectamine 2000 in one well of a six-well tissue culture plate.
After transfection of the fibroblasts in a well of a six-well dish with Lipofectamine 2000 followed by selection with G418 for 8–10 days, 20 colonies were picked up for subculture to expand the cell number. Eight of the 20 cell lines were integrated with pOGN2 and confirmed by PCR analysis (Fig. 2).
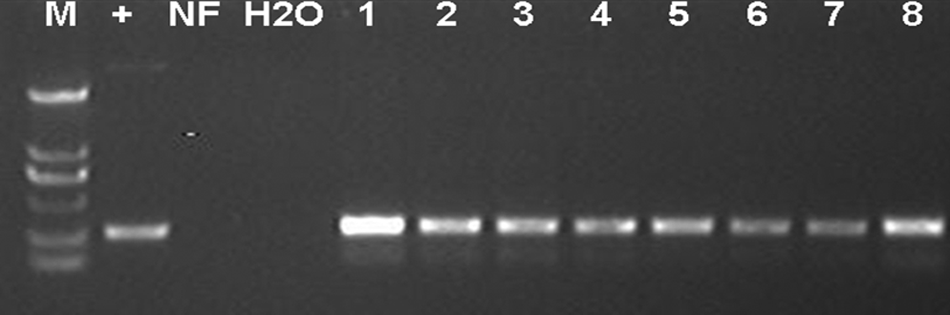
PCR analysis for PEF cell lines stably transfected with pOGN2. Genomic DNA was prepared by cell lysate and subjected to PCR using primers (egfp-p1, egfp-p2), amplifying a 300-bp DNA fragment. M, DL2000 maker; +, positive control with pOGN2 as PCR template; NF, negative control with nontransfected PEFs as PCR template; H2O, blank control with H2O as PCR template; 1–8, cell lines.
EGFP expression in SCNT embryos
The eight transgenic cell lines were used as donor cells for SCNT. Between 30 and 45 NT embryos were made for each cell line. The embryos were checked 5–7 days after culture, and those derived from three cell lines—#3, #4, and #8—strongly expressed EGFP (Fig. 3 and Table 1). Weak expression was observed in embryos from #1 and #2 PEF cell colonies. No expression of EGFP was found in parthenogenetic embryos and in those from #5, #6, and #7. Interestingly, on days 7 after culture, the developmentally arrested embryos in all stages before reaching blastocyst stage were found to express EGFP.
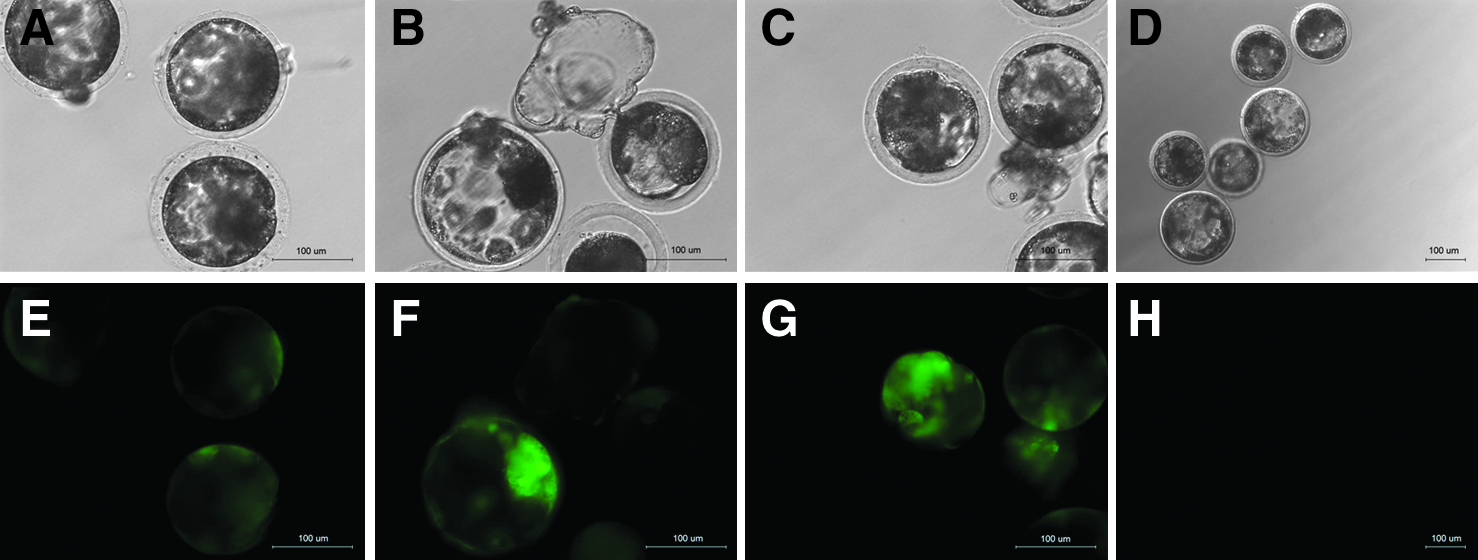
EGFP expression in SCNT embryos reconstructed with transgenic cell lines #3 (
Weak EGFP fluorescence observed.
PA, parthenogenetic embryos.
Expression of endogenous Oct-4in EGFP-positive SCNT embryos
To determine if EGFP expression driven by the porcine Oct-4 promoter in SCNT embryos correlated with endogenous Oct-4 gene expression, EGFP expressing embryos in different stages were subjected to an immunofluorescence test with anti-Oct-4 antibodies after fixation. All the EGFP-positive embryos also expressed endogenous Oct-4 (Supplementary Fig. S3).
EGFP expression in reprogrammed porcine iPS cells
Fifteen days after the transgenic cell lines (a mixture of #3, #4, and #8 cell lines) were infected with retroviral vectors encoding the 4 factors (Oct-4, Sox2, Klf4, and c-Myc), hundreds of iPS-like cell colonies appeared in each dish. Out of them, about 10 to 20 colonies displayed fluorescence in different intensity when cultured to days 20 (Fig. 4). When EGFP-positive colonies were picked for passaging, no iPS cell lines were established. The subcultured colonies gradually stopped either growing or differentiating, with fluorescence simultaneously becoming faint.
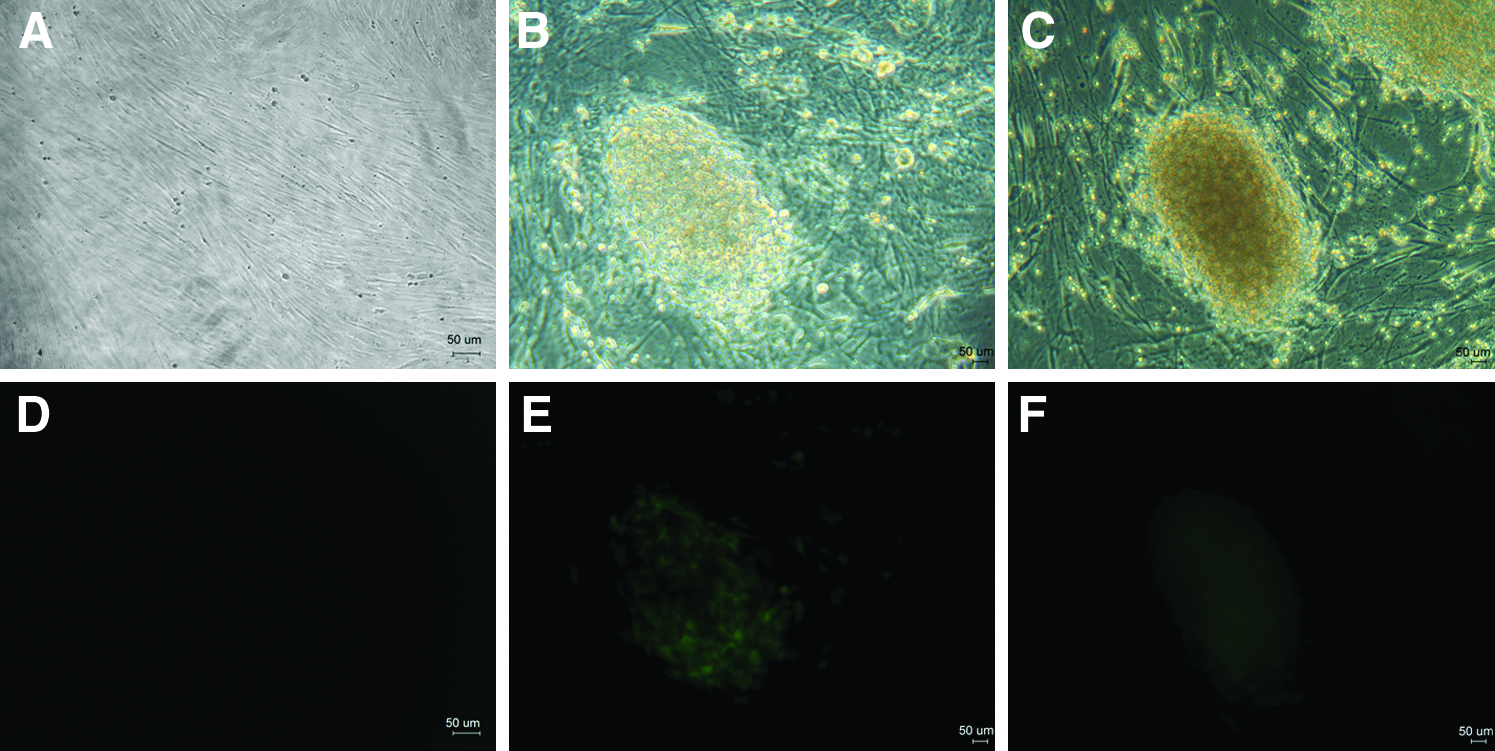
EGFP expression in reprogrammed porcine iPS cells. (
Discussion
Since the first murine iPS was generated in 2006 (Takahashi and Yamanaka, 2006), there have been many successful reports of establishment of iPS from both human and mouse cells (Esteban et al., 2009a; Jia et al., 2010; Kim et al., 2009; Woltjen et al., 2009), and the reprogramming system has been greatly improved. As with the establishment of ES cells, establishment of iPS from pigs lags far behind that from humans and mice. One of the main reasons might be lack of knowledge about the regulatory mechanism of differentiation and reprogramming of swine embryos and cells. To study those basic issues, an animal model with a reporter system for monitoring differentiating and reprogramming status is essential. We considered that an Oct4–EGFP reporting system could be an ideal model for such a purpose, similar to the Oct4–EGFP transgenic mouse used extensively in murine iPS and ES cell research (Kunisato et al., 2009).
Differences in the Oct-4 promoter might exist between different species. This hypothesis was supported by the alignment of Oct-4 promoter between pig, mouse and human cells, where differences in some regions were found, especially in the PE region. These differences might lead to different expression efficiency when the Oct-4 promoter from one species is used in another. As reported in rabbits, embryos failed to express EGFP with a mouse Oct4–EGFP reporter system (Hao et al., 2007). When combined with those above, our results suggested that there was discrepancy in the regulatory mechanism of Oct-4 among different animals. In order to monitor a specific animal species accurately, EGFP expression in the Oct4–EGFP reporting system had better be under the control of the Oct-4 promoter from the species itself. Therefore, to establish an applicable Oct4–EGFP reporting system in pigs, Oct-4 promoter from pigs instead of mice was chosen to monitor the endogenous Oct-4 expression.
SCNT is a good approach for reprogramming a differentiated cell to a pluripotent cell, which can lead to the birth of cloned animals when the reconstructed embryo is transferred to a surrogate (Lai et al., 2002). We utilized this nuclear transfer approach to verify the feasibility of our porcine Oct4–EGFP reporting system. Somatic cells integrated with pOGN2 vector could be reprogrammed into pluripotent cells in SCNT embryos, and fluorescence could be observed in such cells. In our experiment, we used eight cell lines to make SCNT embryos, and the fluorescence was observed in embryos derived from three of them. Not all cell lines gave rise to glowing embryos, because the epigenetic modification and random integration of the transgene in some cell lines might cause difficult to be reprogrammed and gene silencing, respectively, leading to different expression levels of the foreign gene in different cell lines. The fluorescence of the embryos from those three lines could be observed with the naked eye, indicating that our system works well in monitoring pluripotency of porcine cells. A similar attempt has been made in swine by Miyoshi et al. (2009, 2010) and Ozawa et al. (2010), in which EGFP was driven by the murine Oct-4 promoter. From the pictures presented by the authors, fluorescence in SCNT embryos appeared not strong enough to be observed clearly with the naked eye and a special software system was used to test the intensity of fluorescence, which would not be convenient for practical use in monitoring the differentiating status of the cells.
One of the important uses of Oct-4 promoter-driven EGFP reporter system is to monitor the progress of establishment of induced pluripotent cells from somatic cells. The same group in Japan fused the transgenic PEFs bearing Oct-4–EGFP transgene with F9 mouse embryonal carcinoma cells and exhibited generation of cell colonies with distinct EGFP expression. To further verify our reporting system more directly, we tentatively used the transgenic cells to induce pluripotent cells. We observed some colonies expressing EGFP after 20 days' induction by transfection with four factors (Oct-4, c-Myc, Sox2, and Klf4), but no iPS cell lines were established, mainly because the starting transgenic cells for induction were senescent due to long-term drug selection in vitro. There was a clear trend of decreasing induction efficiency over the course of cell passage in vitro. The induction efficiency for the less-passaged cells was higher than that for the more passaged ones (Park et al., 2008). Previous experiments in our group had also found that the induction efficiency of porcine iPS was extremely low when the PEFs were cultured over five passages in vitro (unpublished data).
In conclusion, we have successfully established a porcine Oct4–GFP reporting system. This work lays a foundation for the generation of a pig strain with the Oct4–EGFP reporter system, which would be greatly helpful in studying the differentiating and reprogramming mechanism of pig embryos and cells and could eventually lead to the establishment of totipotent porcine stem cell lines.
Footnotes
Acknowledgments
This work was supported by grants from the National High Technology Research and Development Program of China (2006AA02A103), The Hundred Talents Project of Chinese Academy of Science, (A0858), the National 973 Program of China (2009CB941001), and the Natural Science Foundation of Guangdong Province, China (9251066302000001, 8451066302001811) to L.X.L.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.