
Abstract
Abstract
Human embryonic stem cells (hESC) are self-renewing and pluripotent cells that hold great promise. Our objective was to compare the effect of three different embryo culture methods for derivation of human embryonic stem cells from discarded embryos. A prospective and randomized trial was conducted using 381 discarded human embryos at days 2–3 postfertilization in Beijing Obstetrics and Gynecology Hospital IVF center. After removal of the zona pellucida, discarded human embryos were cultured by three different methods as multiple embryo aggregates, single embryo, and blastomeres. Outgrowth of embryos and hESC derivation were observed. The outgrowth rate of embryos cultured as multiple embryo aggregates was higher than that of those cultured as single embryos or blastomeres (p < 0.05). Three propagating hESC lines were derived from poor quality day 2–3 postfertilization nonblastocyst embryos cultured as multiple embryo aggregates. Derived hESC lines expressed hESC-specific markers of pluripotency and had normal diploid karyotype. The cells were able to form derivatives of all three germ layers in vivo as teratomas. Our results demonstrate that culturing these discarded embryos as multiple embryo aggregates was more profitable for outgrowth and derivation of ESC line than culturing these as single embryo or blastomeres.
Introduction
During IVF procedures, early cleavage-stage embryos are evaluated for uterine embryo transfer or cryopreservation according to morphological and growth characteristics (Scott et al., 2007; Sjoblom et al., 2006; Wharf et al., 2004). Embryos deemed poor quality due to developmental arrest or morphological abnormalities are routinely discarded as medical waste (Alikani et al., 2000; Munné et al., 2007). However, there is evidence that these poor-quality embryos still contain a substantial number of viable cells, suggesting that this class of nonviable embryos could be a rich source of cells for derivation of hESC lines (Alikani and Willadsen, 2002; Gavrilov et al., 2009).
Recently, several hESC lines have been derived from blastocysts that were developed from poor-quality embryos (Chen et al., 2005; Liu et al., 2009; Mitalipova et al., 2003). These embryos were cultured in vitro prior to the derivation of hESC lines, but only a small number developed successfully to blastocysts (Chen et al., 2005; Liu et al., 2009; Mitalipova et al., 2003). There have been the only two reports of the successful derivation of hESC lines directly from poor-quality embryos that were not blastocysts, and the derivation efficiency was low (Lerou et al., 2008; Zhang et al., 2006). It will be important to explore whether derivation efficiency from poor-quality embryos can be improved by embryo aggregation, hES cell coculture, or other methods (Bendall et al., 2007; Boiani et al., 2003; Lerou et al., 2008).
In the present study, poor-quality embryos that would otherwise be discarded were taken at day 2–3 postfertilization and cultured in vitro as blastomeres, single embryo, and multiple embryo aggregates after removal of the zona pellucida. Three hESC lines were derived from the poor-quality embryos cultured as multiple embryo aggregates. The derived hES cells expressed hESC-specific pluripotency markers such as Oct-4, SSEA-4, TRA-1-60, TRA-1-81, and demonstrated pluripotency as evidenced by teratoma formation following transplantation into immunodeficient mice. This demonstrates that the outgrowth rate of poor-quality embryos and hESC derivation efficiency can be improved by culturing the embryos as multiple embryo aggregates.
Materials and Methods
Collection of embryos
This study was approved by the Ethics Committee of Beijing Obstetrics and Gynecology Hospital, affiliated with the Capital University of Medical Science. Embryos produced by IVF for clinical purposes evaluated to be poor quality at day 2–3, and therefore unsuitable for uterine transfer or cryopreservation, were donated for research after patient informed consent was obtained. Embryologists blinded to study enrollment status graded the embryos according to the morphological grading criteria (Brinsden, 1999). Only grade 4 embryos that were not used for transfer or freezing on day 2–3 were used.
Derivation of hESC lines
All discarded embryos were transferred to the laboratory for derivation of hESC lines. After a short period of incubation in acid Tyrode's solution to remove the zona pellucida (ZP), the denuded embryos were plated on inactivated mouse embryonic fibroblast feeder (MEF) and cultured as single embryos (Fig. 1A). Moreover, three to four denuded embryos were plated on inactivated mouse MEF as multiple embryos aggregates (Fig. 1B). In order to ensure that the statement of the patient source, the embryos that have been pooled came from the same patients. In addition, some denuded embryos were separated into single blastomeres by brief exposure to 0.05% Typsin (Gibco-BRL, Gaithersburg, MD), after which single blastomeres were plated on inactivated MEFs and cultured (Fig. 1C). The culture medium consisted of Knock-out Dulbecco's modified Eagle's medium (Gibco) supplemented with 20% Knock-out serum replacement (Gibco), 0.1 mM β-mercaptoethanol (Hyclone, Logan, UT), 1 mM nonessential amino acids (Hyclone), 2 mM glutamine (Hyclone) and 4 ng/mL basic fibroblast growth factor (bFGF2; Sigma, St. Louis, MO). After the embryos attached and showed formation of primary hESC outgrowth, they were mechanically dispersed into small clumps. The clumps were then transferred to freshly prepared feeders in the presence of ESC medium.
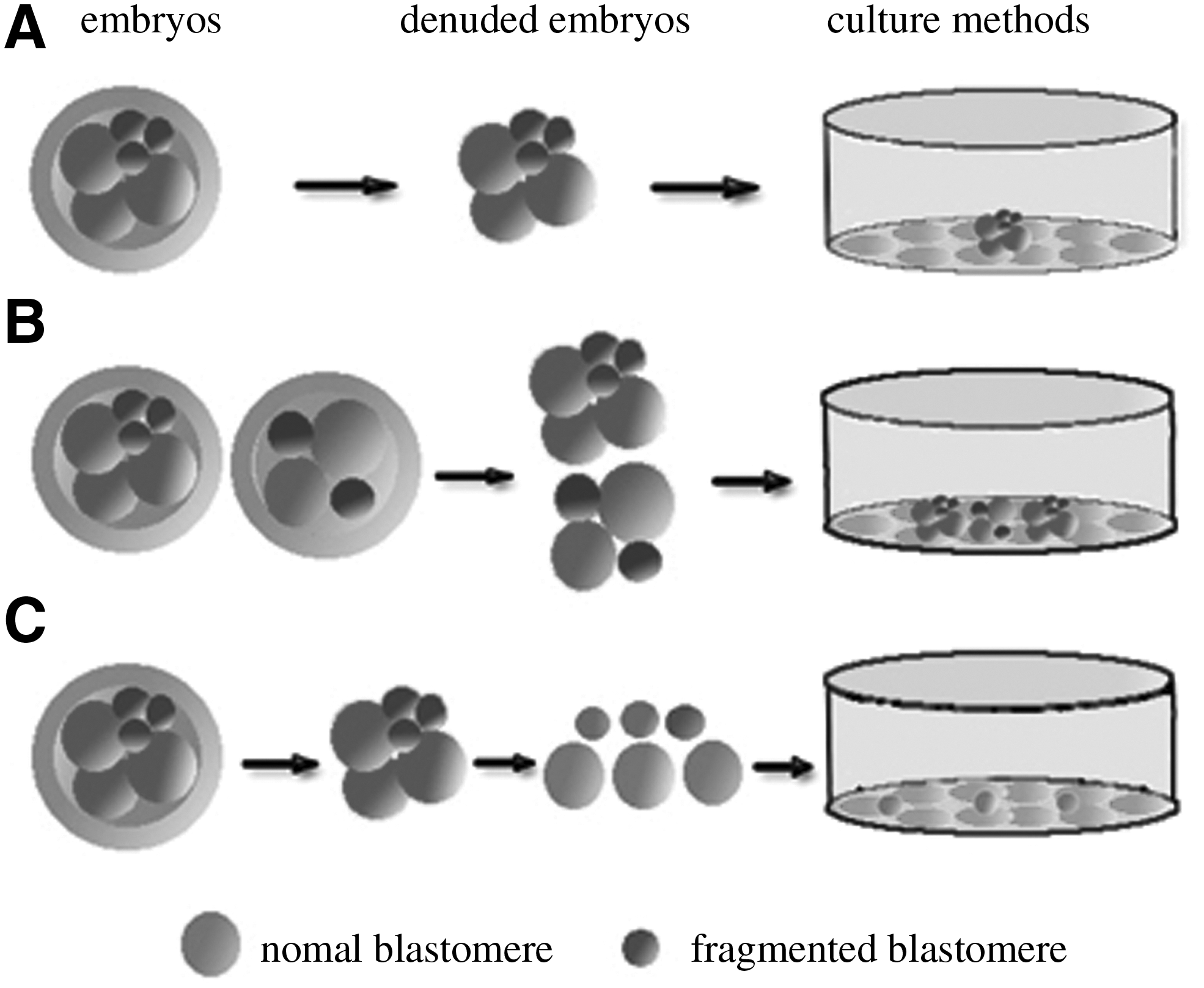
Poor-quality, day 2–3 embryos were cultured under three different methods after a short period of incubation in acid Tyrode's solution to remove the zona pellucida: (
Immunocytochemistry
Cells were analyzed by immunohistochemical staining for hESC surface markers including SSEA-1, SSEA-4, TRA-1-60, TRA-1-81, SOX2 (all from Chemicon, Temecula, CA), and OCT4 (Santa Cruz Biotechnology, Santa Cruz, CA). Colonies of hESCs were incubated with primary monoclonal antibodies (1:10-1:50 dilution) at 4°C overnight. Localization of antigens was visualized with fluorescent-lableled anti-rabbit or antimouse IgG secondary antibodies (Santa Cruz) for 1 h at room temperature, and nuclei were counterstained with DAPI. Mouse or rabbit normal serum was used as a negative control.
Alkaline phosphatase activity
To detect alkaline phosphatase (AP) activity, the cells were fixed with 4% paraformaldehyde, rinsed in a buffer of pH 9.5 containing 100 mM Tris-HCl, 100 mM NaCl, and 50 mM MgCl2, and incubated with nitro blue tetrazolium chloride/5-bromo-4-chloro-3-indolyl phosphate (Roche Applied Science, Mannheim, Germany) in the dark for approximately 20 min. When the desired color intensity was obtained, the reaction was stopped and the stained colony was observed under light microscopy.
Karyotype analysis
To analyze the karyotype of the hESCs, cells were treated with 0.1 μg/mL colcemid for 3–4 h, trypsinized with 0.25% trysin for 3–5 min, resuspended in hypotonic KCl solution (Sigma), incubated for 30 min at 37°C, and then fixed in 3:1 methanol:acetic acid at room temperature for 5 min, pipetted and harvested. After Giemsa staining, at least 20 cells were examined for the chromosome analysis. Chromosomes were visualized using a standard G-band staining technique. Karyotype was analyzed using chromosome pair software (LEICA Q550CW Roche Applied Science).
DNA fingerprinting
To confirm the hESC origins, short tandem repeat (STR) analysis was performed in the center of forensic sciences at Beijing Genomics institute. Genomic DNA was extracted using Qiagen Dneasy Tissue Kit (Qiagen, Chatswirth, CA) according to the manufacturer's instructions. STR analysis was conducted using the Promega PowerPlex 16 System Kit (Promega, Madison, WI). The 18 STR loci were D3S1358, TH01, D21S11, D18S51, Penta E, D5S818, D13S317, D7S820, D16S539, CSF1PO, amelogenin, vWA, D8S1179, TPOX, D2S1338, D19S433, D6S1043, and FGA. Capillary electrophoresis were performed on an automated ABI 3100 Genetic Analyzer and the data were analyzed using GeneMapper V3.2 software (Applied Biosystems, Bedford, MA).
Differentiation potential in vivo
The differentiation potential of the hESC lines in vivo was assessed by their ability to form teratomas. Clumps of 200∼300 cells from hESC colonies digested with collagenase IV (Gibco) were injected into the hind leg muscles of 6-week-old severe combined immunodeficiency (SCID) mice. Tumor formation was observed weekly until it reached a size of ∼2 cm. Tumor was fixed in 4% paraformaldehyde, embedded in paraffin, and examined histologically with hematoxylin and eosin staining and immunohistochemistry assays. The primary antibodies were applied, including polyclonal rabbit anti-β-III-tubulin (Sigma, 1:1000) for neuronal, monoclonal mouse antimuscle actin (Santa Cruz, 1:100) for mesodermal and anti-alpha fetoprotein (Sigma, 1:1000) for endoderm, as well as monoclonal antibodies specific for proliferating cell human nuclear antigen (Chemicon, 1:100). Then incubated in a moist chamber at 4°C overnight. Immunodetection was performed using the Anti-Mouse and the Anti-Rabbit Non-Biotin HRP Detection System (Zymed Laboratories, San Francisco, CA).
Results
Culture of the poor-quality embryos
In this study, hESC line derivation was attempted with a total of 381 poor-quality human embryos (Table 1). Embryos were obtained at 2–3 days postfertilization with severe fragmentation or low cell number. Following removal of the zona pellucida, 126 embryos were cultured as single blastomeres, 115 were cultured as single embryos, and 140 were cultured as multiple embryo aggregates on MEF feeder layers. Cell attachment, survival, and outgrowth were observed daily.
Results of three methods of culture
Of the 126 embryos disaggregated into single blastomeres, 4 (3.2%) formed clumps that produced outgrowths, but no stable hESC lines were derived. Of the 115 embryos cultured as single embryos, 5 (4.3%) developed outgrowths, but similarly no stable hESC lines derived. Of the 140 embryos cultured as multiple embryo aggregates, 12 (8.5%) developed outgrowths, and three stable hESC lines were derived. The derivation efficiency for the embryos cultured as multiple embryo aggregates was 2.1% (Table 1).
All outgrowths were dissected mechanically and replated on fresh MEF feeder layers. From the 12 outgrowths formed by the multiple embryo aggregates, three gradually developed increased stem cell-like morphology (Fig. 2C and D). These large outgrowths were dissociated into several cell aggregates mechanically. Cell clumps were plated on fresh feeder layers and the colonies that developed were passaged using standard enzymatic methods. All hESC lines derived were characterized by morphology similar to hESC lines described in other studies (Reubinoff et al., 2000; Thomson et al., 1998). Colonies were flat with a compact colony structure (Fig. 2I) and contained round and small cells with a high ratio of nucleus-to-cytoplasm and prominent nucleoli (Fig. 2I).
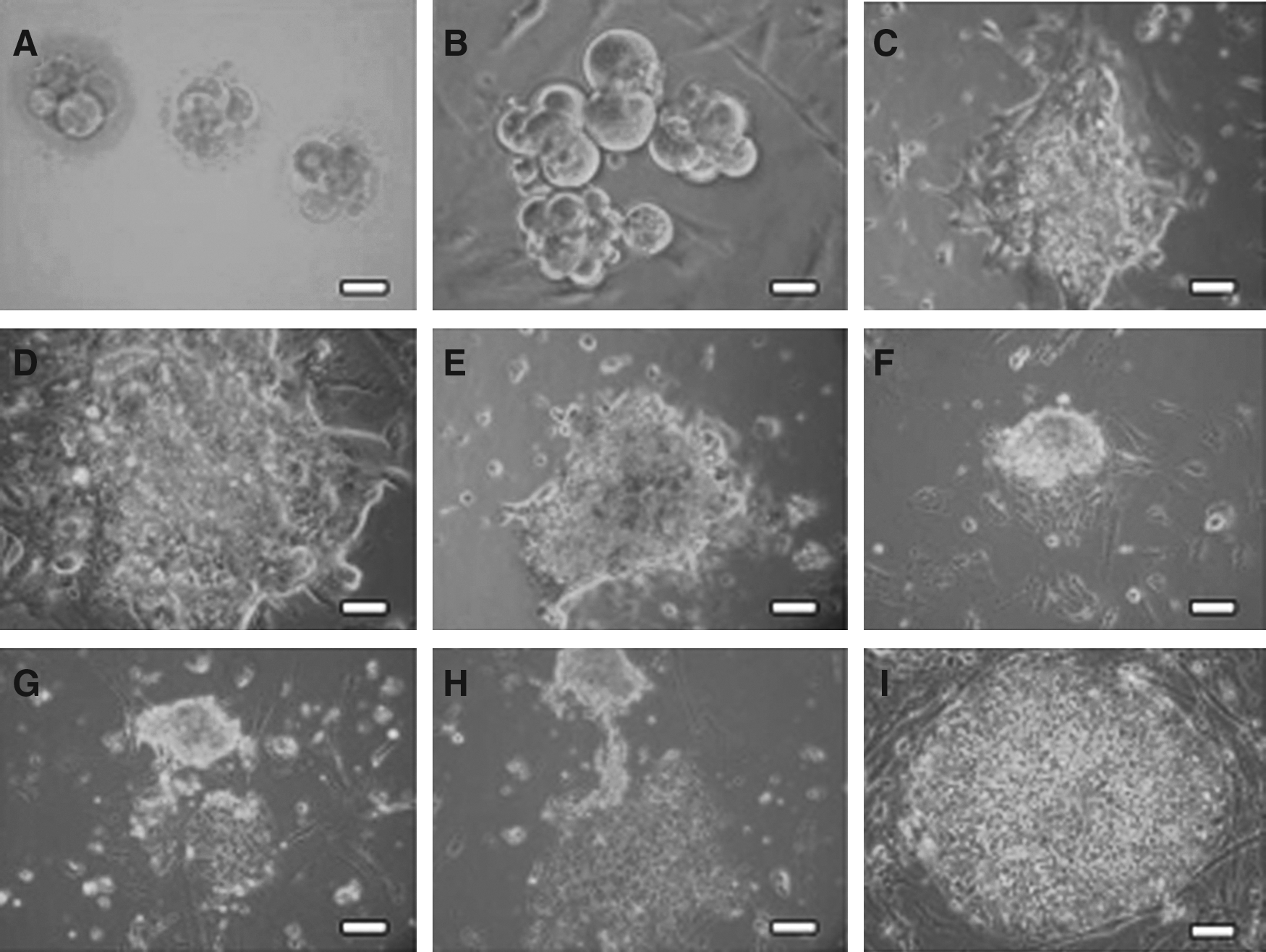
Morphology of poor-quality day 2–3 embryos and culture of multiple embryo aggregates for derivation of human embryonic stem cell (hESC) lines. (
Characterization of the hESC line
All hESC lines derived were positive for hESC-specific markers including OCT4, SOX2, NANOG, SSEA-4, TRA-1-60, and TRA-1-81 (Fig. 3). Cells also stained positive for alkaline phosphatase activity (Fig. 3). Karyotype analysis revealed that these three hESC lines had normal 46 XX karyotypes (Fig. 4A). Short tandem repeat (STR) loci analysis demonstrated a unique genetic identity of each line (Fig. 4B). They also showed in vivo pluripotency, able to form teratomas when cells were injected into SCID mice. Histological examination of derived tumors revealed tissues of all three embryonic germ layers (Fig. 5A and B), and the staining of human specific nuclear antigen showed the teratoma was human origin (Fig. 5C).
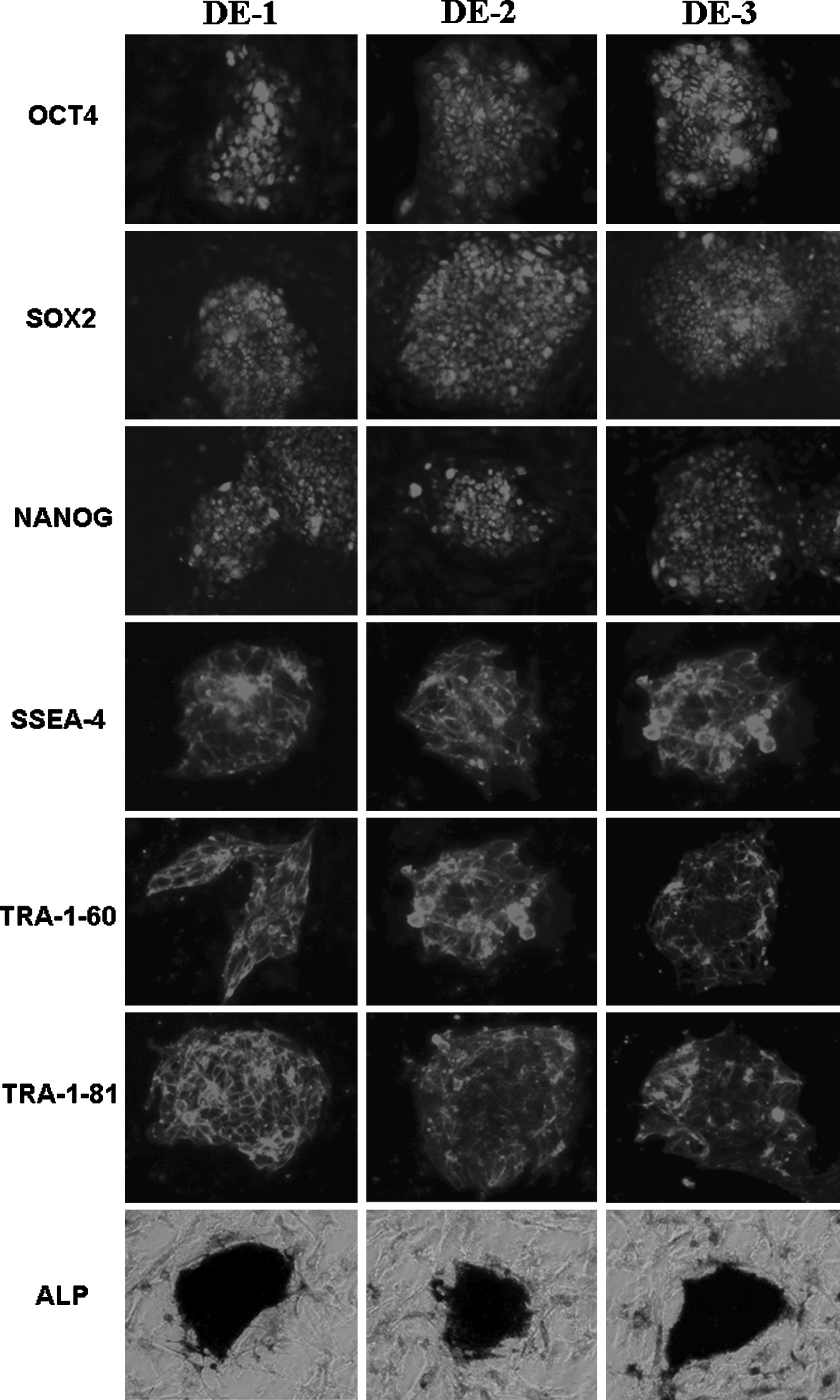
Human embryonic stem cell-specific surface marker and alkaline phosphatase expression of the hESC lines. Immunochemical staining indicated that the hESC lines expressed pluripotent markers: Oct-4, SOX2, NANOG, SSEA-4, TRA-1-60, TRA-1-81. Scale bar: 50 μm. Photomicrographs showed alkaline phosphatase (AP) activity-positive. Scale bar: 100 μm.

Karyotype analysis and DNA fingerprinting of the hESC lines. (

The hESC lines demonstrated pluripotency in vivo by teratoma formation following transplantation of cells into SCID mice. (
Discussion
hESC lines are typically derived from the inner cell mass (ICM) of blastocysts (Stojkovic et al., 2004; Thomson et al., 1998), with a minority of lines derived from the eight-cell stage morulae (Streclchenko et al., 2004). The embryos used in the current study were of poor quality at day 2–3 of development, and were designated for disposal because they were unsuitable for uterine transfer or cryopreservation. In IVF laboratories, many human embryos are discarded because of their poor quality and low implantation rates. We derived three functional hESC lines directly from nonblastocyst poor-quality embryos at day 2–3 postfertilization. These show that, under proper culture conditions, poor-quality embryos at the preblastocyst developmental stage can be used for the derivation of hES cells. Until now, there have been only two reports of the successful derivation of hESC lines directly from nonblastocyst poor quality embryos (Lerou et al., 2008; Zhang et al., 2006). Embryos used in these studies were 6–7 days old (Zhang et al., 2006) or 3–5 days old (Lerou et al., 2008). To the best of our knowledge, the day 2–3 discarded embryos used in our study represent the earliest embryos to date that have successfully yielded stable hES cells.
Our results indicate that the outgrowth rate of embryos and the derivation efficiency of hES cells can be affected by method of embryo culture. We show for the first time that different embryo culture methods, including culturing cells as blastomeres, single embryo, and embryo aggregates, affect the outgrowth rate and the success of using preblastocyst poor-quality embryos to establish hESC lines. In our study, the outgrowth rate of the multiple embryo aggregates was significantly higher than that of blastomeres or single embryos (p < 0.05); the derivation efficiency of hESC lines under multiple embryo aggregates culture was 2.1%, whereas no hESC lines were derived by a single embryo or blastomere culture. In contrast, the derivation efficiency was only 0.6% (1/171) from day 3 poor-quality embryos (Lerou et al., 2008), and 0.8% (1/132) from early (days 3–5) and late (days 6–7) arrested embryos (Zhang et al., 2006). Our findings demonstrate that multiple embryo aggregates culture has a higher outgrowth rate of embryos and a higher efficacy for hESC line derivation than a single embryo or blastomeres culture. It proves that the derivation efficiency of hESC lines from poor-quality embryos can be improved by embryo aggregation. Moreover, the DNA fingerprinting demonstrated a unique genetic identity of each line. Our result maybe provides a benefit for research on finding more effective methods to establishing an hES cell line, which is still crucial for scientific research. It gives the clue to increase the efficacy for hESC line derivation by using aggregation of embryos to create cell lines. The higher outgrowth rate of multiple embryo aggregates may be due to enhanced interaction of blastomeres, which may the result in an increased ability of cells to attach, grow, and proliferate. Cytokines or growth factors secreted by blastomeres may also promote and direct the transition of these cells to form stably growing hESCs. Further research is necessary to identify the specific factors and molecular mechanisms that account in higher efficiency of culture.
These hESCs we derived in this study exhibited morphology similar to that previously described (Reubinoff et al., 2000; Thomson et al., 1998). Specifically, these cells appeared as compact colonies containing small and round cells with high nuclear-to-cytoplasm ratios and prominent nucleoli. Characterization of these hESC lines revealed presence of hESC specific markers, pluripotency, and normal karytype. DNA fingerprinting demonstrated a unique genetic identity of each line. Typically, day 2–3 postfertilization embryos that appear to be poor quality are discarded during IVF. Our results confirm further that such embryos contain viable blastomeres with the potential to form stable and pluripotent embryonic stem cell lines with normal karyotype.
Taken collectively, these findings demonstrate that discarded poor-quality embryos have viable blastomeres and can be induced to become hES cells under proper conditions. Multiple embryo aggregate culture is more profit for the outgrowth and derivation of human embryonic stem cells from discarded embryos than single-embryo culture and blastomeres culture.
Footnotes
Acknowledgments
The authors are thankful to Dr. Ling-song Li for technique support. This study was supported by grants from Beijing New Star Technology Project (H020821200190), Beijing Collective Talents Project (20042B0301037), and Beijing Tens-Hundreds-Thousands of Health Personnel Project.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.