
Abstract
Abstract
Given the difficulties in establishing bona fide porcine embryonic stem cells, we considered it would be interesting to investigate histone modifications, X chromosome inactivation (XCI), deacetylation, DNA methylation, and gene expression around the time of inner cell mass (ICM) and epiblast formation in sexed embryos. We found that the porcine epiblast expressed lower levels of NANOG and C-MYC, of which, we speculate may be one indication for the difficulties in obtaining embryonic stem cells (ESCs) from the porcine embryonic epiblast. Our research revealed distinct expression of lineage-specific-, early gastrulation-, and pluripotency-associated genes between the E10 epiblast and trophectoderm and between sexes. We determined that H3K27me3 was hypermethylated in the E6 embryo and hypomethylated in the E10 epiblast. Interestingly, we also observed exclusive localization of H3K4me3 in the E6 ICM, which may be a key marker for early lineage segregation in the pig. We also observed that the methyltransferases of H3K4me3, H3K27me3 and H3K9me3 and the DNA methyltransferases differed between male and female E10 embryos, and between tissues. We consider that epigenetic mechanisms, which are modified by specific enzymes, may be important for both early lineage segregation events and XCI, and these may further effect the levels of downstream-targeted gene expression in the different sexes.
Introduction
During mammalian embryonic development, the first lineage specification occurs at the morula stage, and results in the establishment of the ICM (which gives rise to the embryo proper) and the trophectoderm (TE) (which gives rise to the extra-embryonic tissues). Shortly thereafter, another fate specification is made, whereby the ICM differentiates into the hypoblast and the epiblast. Embryonic stem cells (ESCs) are pluripotent cells, derived from either the ICM or epiblast (Evans et al., 1981; Tesar et al., 2007; Thomson et al., 1998). It is generally accepted that murine ESCs are derived from the ICM (Martin, 1981), whereas human ESCs and mouse epiblast stem cells (EpiSCs) are derived from the epiblast (Brons et al., 2007; Tesar et al., 2007; Thomson et al., 1998). Key features differ between these embryonic-derived stem cell types. Cell renewal and proliferation of mouse ESCs and human ESCs is regulated through different cell signaling pathways, and mouse EpiSCs share very similar signaling pathways to human ESCs (Tesar et al., 2007; Valdimarsdottir et al., 2005). Therefore, cell signaling differences are related to the embryonic origin of the stem cells, rather than differences between the species. Although both mouse and human ESCs can differentiate into all cell types of the body in vitro, some evidence suggests that chimerism and germ-line transmission is lower in EpiSCs (Guo et al., 2009; Tesar et al., 2007). For ethical reasons, human ESCs have not been used to form chimeras. To date, there is lacking evidence on whether epigenetic differences may exist between the ICM and epiblast. Furthermore, gender may play a role in epigenetic changes, which we consider is worth investigating further.
In mammals, one of the female X chromosomes is silenced during early development, in order to balance the unequal X-chromosome dosage between the male and female sex chromosomes. This process is termed XCI, and occurs by both imprinting and inactivation. In female (XX) mouse blastocysts, the imprinted XCI occurs when the ICM and TE are formed and is retained as a gender-specific inactivation of the paternal X chromosome (Xp) in TE (Takagi and Sasaki, 1975; West et al., 1977). Later, in the epiblast, either Xp or the maternal X chromosome (Xm) is randomly selected for inactivation (Rastan, 1982; Takagi et al., 1982). In both imprinted XCI and random XCI, the silencing of future inactive X chromosomes is initiated by the X-inactive specific transcript (Xist) (a noncoding gene), through propagation of its RNA along the X chromosome (Kay et al., 1993; Marahrens et al., 1997; Penny et al., 1996). Immediately following the destined inactivated X (Xi) is coated by Xist RNA, several types of epigenetic events occur in the mouse on the Xi, including, deacetylation of histone 3 (H3) and histone 4 (H4), methylation of lysine 9 and lysine 27 of H3, lysine 20 of H4, ubiquitination of H2A and DNA methylation of CpG islands (de Napoles et al., 2004; Heard et al., 2001; Keohane et al., 1996; Kohlmaier et al., 2004; Plath et al., 2003; Silva et al., 2003; Smith et al., 2004). Given this important epigenetic event, XIST expression should be evaluated together with the investigation of epigenetic marks in developing porcine embryos.
Research has shown that the first lineage segregation event, where the ICM and TE form (following compaction of the blastomeres and blastulation of the embryo), results in a complex change in gene expression between the resultant cell types. OCT4, also named POU5F1 (POU domain, class 5, transcription factor 1), is a gene that is expressed in the mouse and human ICM (Nichols et al., 1998). Evidence suggests that OCT4 is essential for ICM maintenance, but is not required for initial lineage segregation (Nichols et al., 1998). In contrast, Cdx2 (caudal-related homeobox 2) expression is, at least in the mouse, restricted to the TE, and embryos that lack Cdx2 are unable to form TE, which suggests that Cdx2 may play a role in segregation. Mutual antagonism of Oct4 and Cdx2 is also evident (Niwa et al., 2005). Another key gene involved in pluripotency and lineage segregation is Nanog. Expression of this transcription factor in the mouse is restricted to the ICM of blastocysts and is required for the specification of the epiblast (Chambers et al., 2003; Mitsui et al., 2003). The absence of Nanog expression in mouse ESCs results in loss of pluripotency and differentiation into extraembryonic endoderm (Hatano et al., 2005; Mitsui et al., 2003). In mouse embryos, specification of the primitive endoderm is thought to result from mutual antagonism between Nanog and Gata6. Interestingly in the pig, OCT4 and NANOG expression differs compared to mouse and human (Kuijk et al., 2008). Specifically, OCT4 is not exclusive to the ICM, but is also expressed in the TE (Hall et al., 2009; Keefer et al., 2007). NANOG is not detected at all in the porcine ICM (Blomberg et al., 2008; Hall et al., 2009). Later in development, OCT4 and NANOG are found localized in the porcine epiblast. Given these differences, we considered it would be interesting to investigate the expression profile of these genes and relate these to epigenetic changes in the embryo.
Establishment and maintenance of specific genomic DNA methylation patterns is crucial for nomal development. Following fertilization of the mammalian embryo, demethylation of both parental genome occurs. In the mouse and several other species, the paternal genome is actively demethylated, whereas the DNA of the maternal genome undergoes passive demethylation (Mayer et al., 2000). DNA methyltransferases (Dnmts) are responsible for the establishment and maintenance of DNA methylation. Dnmt3a and Dnmt3b catalyze de novo methylation of unmethylated DNA, and in contrast, Dnmt1 or its isoforms, only methylate hemimethylated DNA, and is important for DNA maintenance methylation (Bestor, 2000; Li, 2002). We consider that it would be worth to investigate the expression of the Dnmts in porcine development and determine how these may be influenced by methylation changes at the time of lineage specification.
Key regulators have been identified, which regulate the methylation state. Polycomb group (PcG) proteins are known to maintain the repressed state of genes. In contrast, trithorax group (trxG) proteins maintain the permissive state. In mammals, two distinct types of PcG proteins have been reported to be tightly associated with the repressive mark H3K27me3, including polycomb repressive complex 1 (PRC1) and 2 (PRC2). PRC1 is considered the effector of PcG-dependent gene repression and is recruited by H3K27me3. PRC2, which is primarily responsible for trimethylating lysine 27 of histone H3, mainly consists of three core components: enhancer of zeste homolog 2 (Ezh2), embryonic ectoderm development (Eed), and suppressor of zeste 12 homolog (Suz12). Ezh2, a SET-domain containing subunit, is the methyltransferase that catalyzes the formation of H3K27me3 (Cao et al., 2002; Czermin et al., 2002; Muller et al., 2002), whereas both Eed and Suz12 are required for Ezh2 to carry out its methyltransferase activity (Cao and Zhang, 2004; Montgomery et al., 2005; Pasini et al., 2004). Interestingly, the Eed–Ezh2 complex plays an important role during initiation of XCI and has been shown to be recruited by Xist to mediate the methylation of lysine 27 of histone H3 (H3K27me3) on the Xi. Furthermore, in differentiated cells of the TE lineage, Eed is required for maintenance of the imprinted XCI (Plath et al., 2003; Wang et al., 2001). Recent evidence has revealed that methyl groups can also be actively removed from H3K27, and two demethylases of H3K27me3, including, Utx and Jmjd3, were identified (Agger et al., 2007; Hong et al., 2007; Lan et al., 2007). Furthermore, specific trxG proteins have been linked to trimethylation of H3K4 (associated with transcriptional permission). In mammals, two trxG proteins, Mll1 and Ash1L, have been identified as key methylases of H3K4me3 (Dou et al., 2006; Gregory et al., 2007). A demethylase of H3K4me3, Rbp2, has also recently been identified (Christensen et al., 2007). From our previous research we have shown that bivalent histone methylation may be involved in porcine lineage differentiation (Gao et al., 2010), but this phenomenon has not been addressed in detail. In this study, we investigate the bivalent histone methylation events in the porcine epiblast and TE in greater detail.
Other methylation events of H3 are also of particular interest. Trimethylation of lysine 9 on H3 has been associated with heterochromatin packing. In both mouse and human, Suv39h1 has been reported to trimethylate H3K9 and create a binding site for the heterochromatic protein, HP1 (Fischle et al., 2003; Jacobs et al., 2001), whereas G9a is believed to be an H3K9me2 methyltransferase, which targets euchromatic loci (Esteve et al., 2005; Lachner et al., 2003; Tachibana et al., 2002, 2005). A crucial role of G9a has been implicated for early mammalian development from knockout studies in mice (Li et al., 1992; Tachibana et al., 2002, 2005). From this, we decided to address the role of H3K9 further in lineage specification.
Hox (homeobox) genes play an important role in the development of the anterior–posterior axis and in spatial patterning of the developing embryo in Drosophila and vertebrates (Morgan, 1997). These genes are also regulated by epigenetic mechanisms (Barber and Rastegar, 2010; Soshnikova and Duboule, 2009). In Drosophila, the regulation of expression is performed by a dynamic alteration of PcG and trxG proteins (Schwartz and Pirrotta, 2007; Simon et al., 1992). Given the important role of Hox genes during development and our own interest in epigenetic regulation in the preimplantation embryo, we considered to study the expression of a number of Hox genes at the time of lineage segregation into the ICM and TE and relate the outcomes to the epigenetic state in the porcine embryo.
Finally, the epigenetic event of acetylation of core histones is associated with exposure of chromatin and subsequent gene transcription. Conversely, deacetylated histone tails, which interact more closely with the DNA, leads to gene repression (Grunstein, 1997; Imhof et al., 1997; Roth and Allis, 1996). Histone deacetylases (HDACs) are responsible for removal of acetyl groups from histone tails and are therefore considered to be transcriptional corepressors. Hdac1 and Hdac2, which belong to class I HDACs, are well studied in mammals (Montgomery et al., 2007; Zimmermann et al., 2007). Although evidence in the mouse indicates that Hdac1 and Hdac2 may play essential roles during embryogenesis (Cowley et al., 2005; Hendrich et al., 2001; Lagger et al., 2002; Montgomery et al., 2007), the roles of Hdac1 and Hdac2 during embryonic lineage specification have not been studied. Given this, we decided to analyze the expression of HDAC1 and HDAC2 at the time of lineage specification and relate this to other epigenetic events.
To conclude, in the mouse, at least three types of epigenetic asymmetry distributions have been reported during ICM and TE segregation. First, the pluripotent ICM exhibits extensive global methylation of H3K27, whereas in the TE, methylation of H3K27 is only detected on the Xi (Erhardt et al., 2003). This was later confirmed by results from a genome-wide survey of promoter occupancy by H3K27me3 (Dahl et al., 2010). Second, during cell specification of the mouse embryo, both the ICM and TE gain DNA methylation gradually, but the level of methylation is higher in the ICM than in the TE (Dean et al., 2001; Santos et al., 2002). Third, H3R26me, a histone modification that has been associated with pluripotency in mouse embryos, displayed different levels in the blastomeres of four-cell stage embryo (Torres-Padilla et al., 2007). It is interesting that those blastomeres that have a higher level of H3R26me are likely to localize to the ICM, and upregulation of H3R26 methyltransferase Carm1 can induce the upregulation of the pluripotency markers Nanog and Sox2 (Torres-Padilla et al., 2007). However, no study has examined the distribution of epigenetic marks in later-stage embryos containing the epiblast, prior to gastrulation, and how gender and related epigenetic events might be involved. Given this, we have compared DNA methylation and histone methylation between the ICM and TE, as well as between the epiblast and TE, in sexed porcine in vivo produced blastocysts. We perform relative real-time PCR to investigate the expression of the related enzymes of several types of epigenetic marks. We also compare the spatial distributions of H3K27me3, H3K4me3 and DNA methylation between the porcine epiblast and TE, and between the porcine ICM and TE by immunocytochemistry, to further investigate the link of epigenetic marks and XCI.
Materials and Methods
Antibodies and reagents
Polyclonal rabbit antibodies against H3K27me3 (Millipore, Billerica, MA) and H3K4me3 (Abcam, Cambridge, UK) were diluted 1:500 and the monoclonal mouse antibody for 5-Methylcytidine (Abcam) was diluted 1:200 (final concentrations of antibodies against H3K27me3, H3K4me3 and 5-Methylcytidine were 2, 2, and 5 μg/mL, respectively). Secondary antibodies used were goat antirabbit IgG (H+L) conjugated with Alexa Fluor 594 (Molecular Probes, Leiden, The Netherlands) and goat antimouse IgG (H+L) conjugated with Alexa Fluor 594 (Molecular Probes) and were both diluted at 1:500 (final concentration of both secondary antibodies was 4 μg/mL). The H3K4me3 peptide (Abcam) and H3K27me3 peptide (Abcam) were diluted 1:250 and 1:50, respectively (final concentrations of peptides H3K4me3 and H3K27me3 were 4 μg/mL and 20 μg/mL, respectively). Mouse IgG1 (Dako, Glostrup, Denmark) was diluted 1:20 (final concentration was 5 μg/mL), and it was used as negative primary antibody control for the monoclonal mouse antibody for 5-Methylcytidine. Other reagents were purchased from Sigma-Aldrich (St. Louis, MO) unless otherwise noted.
Embryo collection
In vivo produced porcine embryos were collected from a total of six pregnant sows with Duroc boar sperm inseminated over 2 consecutive days, by nonsurgical flushing (Hall et al., 2009). Embryo stages obtained included E6 and E10 (days following fertilization). Embryos were morphologically staged according to Vejlsted et al. (2006), and only embryos of distinct morphology were used. This morphology consisted of two stages: the expanded blastocyst containing the pluripotent ICM and TE, and the prestreak I embryo containing the formed pluripotent embryonic disc (epiblast), the underlying hypoblast, and expanded TE. Immediately after flushing, embryos were washed in phosphate-buffered saline (PBS) and the outer zona pellucida was removed by use of 0.5% pronase. A total of 12 E6 embryos were fixed in 4% paraformaldehyde (PFA) and stored short term at 4°C in PBS for immunocytochemical analyses. Twenty-five E10 embryos were randomly assigned for immunocytochemical analyses and 14 were assigned for comparative RT-PCR. In the case of comparative RT-PCR, embryos were mechanically dissected into epiblast and TE (Fig. 1). The isolated epiblast and an equal size of dissected TE from the same embryo was then separately transferred into centrifuge tubes containing RNA lysis buffer (Gao et al., 2010), and plunged into liquid nitrogen. The frozen tissues were stored at −80°C. The remaining TE was collected separately for genomic DNA extraction and later sexing. In the case of immunocytochemical analyses, a small piece of the TE was cut from each E10 embryo, prior to fixation (as described above) and used for DNA extraction and sex determination by PCR.
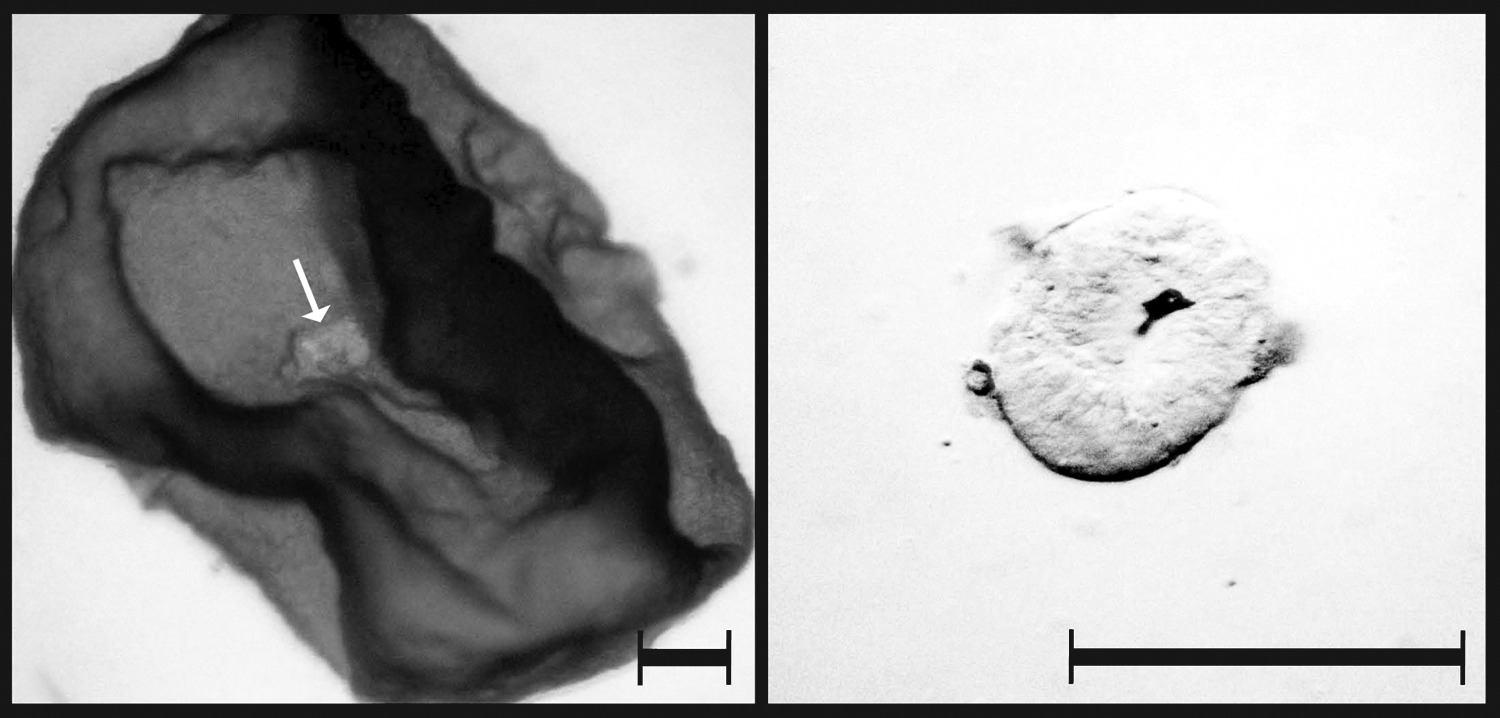
Isolation of epiblasts (white arrow) from E10 blastocyst by mechanical dissection. Scale bar = 300 μm.
RNA extraction and amplification
Total RNA from pooled epiblasts and TE was isolated using the RNeasy micro kit (Qiagen, Valencia, CA). The numbers of pooled male epiblasts, male TEs, female epiblasts, and female TEs used for RNA extraction were 6, 6, 8, and 8, respectively. An additional DNA digestion step was included by using DNase I and performed according to the manufacturer's protocol. An extended (8-h) RNA amplification was performed with approximately 30 ng total RNA from each sample by using the QuantiTect Whole Transcriptome Kit (Qiagen). Amplified cDNA was quantified, aliquoted and stored at −80°C, ready for use.
Gene expression by PCR
Nine Hox genes (HOX A2, HOX A3, HOX A7, HOX A10, HOX B4, HOX B7, HOX C8, HOX D8, and HOX D10) were investigated by use of semiquantitative PCR in porcine E10 embryos (six male epiblasts, six male TEs, eight female epiblasts, and eight female TEs). The amplification conditions and components for each 25-μL reaction are the same as used previously (Gao et al., 2010). The PCR product were run on 2% agarose gels containing ethidium bromide and visualized using a UV gel documentation system (Bio-rad, Hercules, CA).
Immunocytochemistry
Embryos were assigned as following. Nineteen E10 embryos (11 male and 8 female) and five E6 embryos were analyzed for H3K27me3. For 5-Methylcytidine immunostaining, three E10 and five E6 embryos were used, and for H3K4me3 immunostaining, three E10 and two E6 were used. One E10 embryo for H3K4me3 was used for preabsorption immunostaining. The fixed embryos were permeabilized with 1% Triton X-100 in PBS overnight at room temperature and washed briefly with 0.05% Tween-20 in PBS. The E6 embryos were further treated with 2 N HCl for 1 h prior to blocking. Embryos were subsequently blocked for either 1 or 5 h, for E6 and E10 embryos, respectively, at room temperature in either 0.25% bovine serum albumin (BSA) for E10 embryos, or 2% BSA in PBS for E6 embryos. The embryos were then incubated in primary antibodies, diluted in blocking buffer at room temperature for either 1 or 5 h, for E6 and E10 embryos, respectively, washed twice in 0.05% Tween-20 in PBS for 30 min, and incubated with secondary antibodies for 1 h in the dark at room temperature. Finally, embryos were washed twice for 30 min in 0.05% Tween-20 in PBS and counterstained with a DNA labeling dye, 1 μg/mL Hoechst 33258 for 10 min. The embryos were mounted on glass slides (Hounisens Laboratorieudstyr A/S, Risskov, Denmark) using fluorescent mounting medium (Dako, Glostrup, Denmark), covered using a glass coverslip (Hounisens Laboratorieudstyr A/S) and assessed by fluorescence microscopy. Negative controls for the secondary antibody were performed by omitting the primary antibody. Furthermore, negative controls for the specificity of the primary antibodies were also performed: in the case of anti-H3K4me3 and anti-H3K27me3, preabsorptions of the primary antibodies with H3K4me3 and H3K27me3 peptides were performed for 1 h prior to incubation with the embryos. For anti-5-methylcytidine, replacement of the primary antibody with an isotype IgG1 at the same protein concentration was performed.
Sex determination
Genomic DNA from each embryo was extracted from trophectoderm tissue using the DNeasy Blood & Tissue Kit (Qiagen). Sequences of ZFX (X chromosome) and SRY (Y chromosome) genes were amplified by normal PCR using primers from Petkov (Petkov et al., 2009).
Primer design
Primers were designed using Primer Express (Applied Biosystems, Bedford, MA) or manually designed based on available sequences from Database of GenBank (NCBI) or TIGR. The sequences of the PCR primers used for amplifying the targeted products are shown in Supplementary Table S1 (online supplementary data at www.liebertonline.com). Each of the amplicons were sequenced and blasted against their original sequences for verification.
Comparative real-time PCR
Comparative real-time PCR was performed on the LightCycler 480™ (Roche, Switzerland) using LightCycler 480 SYBR Green I Master Mix (Roche Diagnostics, Mannheim, Germany). A total of 2.5 μL cDNA (diluted according to instructions from the QuantiTect Whole Transcriptome Kit) was added to a total volume of 10 μL containing 1× SYBR Green PCR master mix (final concentration) and 0.3 μM (final concentration) of each primer. Reactions were performed using single biological, pooled embryo triplicates, and were repeated in technical triplicates. The housekeeping gene GAPDH was used as the reference gene and E9 (10 E9 pooled embryos, as a control for genes XIST, CDX2, GATA6, EOMES, T, EZH2, EED, SUZ12, JMJD3, UTX, ASH1L, MLL1, and RBP2) or E21 (10 pooled limb bud stage embryos, as a control for genes DNMT3A, DNMT3B, HDAC1, and HDAC2) embryo cDNA was used as the endogenous control. Tissues examined include pooled six male porcine epiblasts, six male porcine TEs, eight female porcine epiblasts, and eight female porcine TEs. The PCR program was as follows: 95°C for 5 min; 45 cycles, 95°C for 10 sec, 60°C for 10 sec, and 72°C for 20 sec; a melting cycle with a range of 65 and 95°C was then performed to confirm specificity of the amplicon and absence of primer dimers and contaminating DNA.
Comparative real-time PCR analysis and statistical analyses
To analyze comparative RT-PCR data, fold changes of gene expression among different samples were calculated by using the delta CT method (2−ΔCT) or the delta delta CT method (2−ΔΔCT). To make sure whether we could compare the transcript levels of different pluripotency genes (OCT4, SOX2, NANOG, C-MYC, and GAPDH) in the same tissue, we calculated the PCR efficiencies of these genes by making standard curves with six different dilutions. Differences in the levels of gene expression among groups were performed using one-way analysis of variance (ANOVA) in Excel. Differences were considered significant at p ≤ 0.05.
Quantification of DNA global methylation
To analyze differences between global methylation in the ICM and TE in E6 embryos and epiblast and TE in E10 embryos, quantification of the nuclear fluorescence (5-methylcytidine) intensity was performed using Image J free software (http://rsbweb.nih.gov/ij/). A total of 16 epiblast cells and 16 TE cells were analyzed in E10 embryos (n = 2) and 12 ICM cells and 12 TE cells in E6 embryos (n = 5). The mean ratio between the fluorescent intensity of the cell types was calculated for both embryo stages. Statistical analysis was performed using one-way ANOVA and statistical differences were observed at p ≤ 0.05.
Results
Comparative expression of sexed embryos shows differences in lineage-specific and gastrulation related genes, but not pluripotency markers
We first decided to evaluate the expression of pluripotency-, lineage-, and gastrulation-associated genes in E10 porcine male and female epiblast and TE, in order to identify whether differences could be observed. Comparative real-time PCR was performed for XIST, the pluripotency markers (NANOG, OCT4, SOX2, and C-MYC) and genes linked to lineage specification (CDX2 and GATA6) and early gastrulation (EOMES and BRACHYURY (T)). XIST, a noncoding RNA associated with the initiation of X chromosome inactivation, was observed to be highly expressed in the female epiblast and TE, and low in the male epiblast and TE (Fig. 2A).

Expression of pluripotency-, lineage- and early gastrulation-associated genes differ in the porcine epiblast and trophectoderm (TE). Comparative real-time PCR expression of (
The PCR efficiencies of NANOG, OCT4, SOX2, C-MYC, and GAPDH were 1.98, 2.11, 1.99, 1.98, and 2.04, respectively, and the Ct values of GAPDH in the four investigated tissues were similar (Ct values of GAPDH in ME, MT, FE, and FT were 14.9, 15.6, 15.6, and 15.9, respectively). Given the similiarity in PCR efficiency, we are able to compare the transcript levels of of NANOG, OCT4, SOX2, and C-MYC in the same tissue. Few differences were determined between gender and the expression of the pluripotency genes. The expression pattern of OCT4 was high in both the male and female epiblast (Fig. 2B). Expression of SOX2 was also high, but at a lower level in male and female epiblast compared with OCT4 (Fig. 2B). Expression of NANOG was also significantly higher in male and female epiblast compared to TE, but lower than both OCT4 and SOX2 (Fig. 2B). Interestingly, the level of NANOG expression was significantly lower in the pooled female epiblast compared with the male epiblast (Fig. 2B). Expression of these pluripotency genes was either extremely low or could not be detected in the TE (Fig. 2B). Interestingly, C-MYC was expressed at low levels both in the epiblast and TE (Fig. 2B).
To examine genes linked to lineage specification and early gastrulation, we analyzed the expression of CDX2, GATA6, EOMES, and BRACHYURY (T). We observed higher expression of CDX2 and GATA6 in the TE, compared to the epiblast (Fig. 2C and D). However, the expression of CDX2 and GATA6 in the female epiblast was significantly higher than in the male counterpart (Fig. 2C and D). Both EOMES and BRACHYURY (T) (two of the T-box family members) were exclusively detected in the epiblast, but not in the TE (Fig. 2E and F). Interestingly, the level of expression of both EOMES and T between male and female epiblast differed significantly, with both being significantly lower in female epiblasts (Fig. 2E and F).
We also evaluated the expression of HOX A2, HOX A3, HOX A7, HOX A10, HOX B4, HOX B7, HOX C8, HOX D8, and HOX D10 by semiquantitative PCR in E10 male and female epiblast and TE. From this, we could determine that HOXA3 and HOXD8 were specifically expressed in the TE of embryos. HOXB4 could also be detected in TE and to a lesser extent, in the epiblast (Supplementary Fig. S1).
The repressive mark, H3K27me3, is globally hypermethylated in the E6 porcine embryo but specifically hypomethylated and showed sexual dimorphism in the E10 epiblast
In order to evaluate the epigenetic mechanism of gene repression at the time of lineage segregation, and in relation to XCI, we immunolabeled E6 embryos (containing the ICM and TE) and E10 embryos (containing the epiblast and TE) with a commercial antibody against H3K27me3 (validated with H3K27me3 peptide in porcine E10 TE) (Supplementary Fig. S2). H3K27me3 was hypermethylated in both the E6 porcine ICM and the TE (Fig. 3). Due to limited cell numbers, these E6 embryos could not be sexed, nor the ICM removed from the TE to perform further expression analyses. However, the E10 embryos were sexed prior to immunolabeling, and high levels of H3K27me3 were observed in the nuclei of both male and female TE cells, with no observable expression in the male epiblast (Fig. 3). A single, small, punctate spot could be detected in most of the female epiblast and TE nuclei, which was not observed in their male counterparts (Fig. 3). We detected hypermethylation of H3K27me3 and higher expression of the methylase (EZH2) and the methylase cofactors (EED, SUZ12) in both the male and female TE (Fig. 4A–C), whereas the methylase and its cofactors were repressed in the epiblast (Fig. 4A–C). Interestingly, EED expression was higher in the female epiblast and TE compared to their male counterparts (Fig. 4B). Surprisingly, the expression of H3K27me3 demethylases JMJD3 and UTX, were also higher in the TE compared to the epiblast (Fig. 4D and E) and did not correspond well to the methylase expression levels.
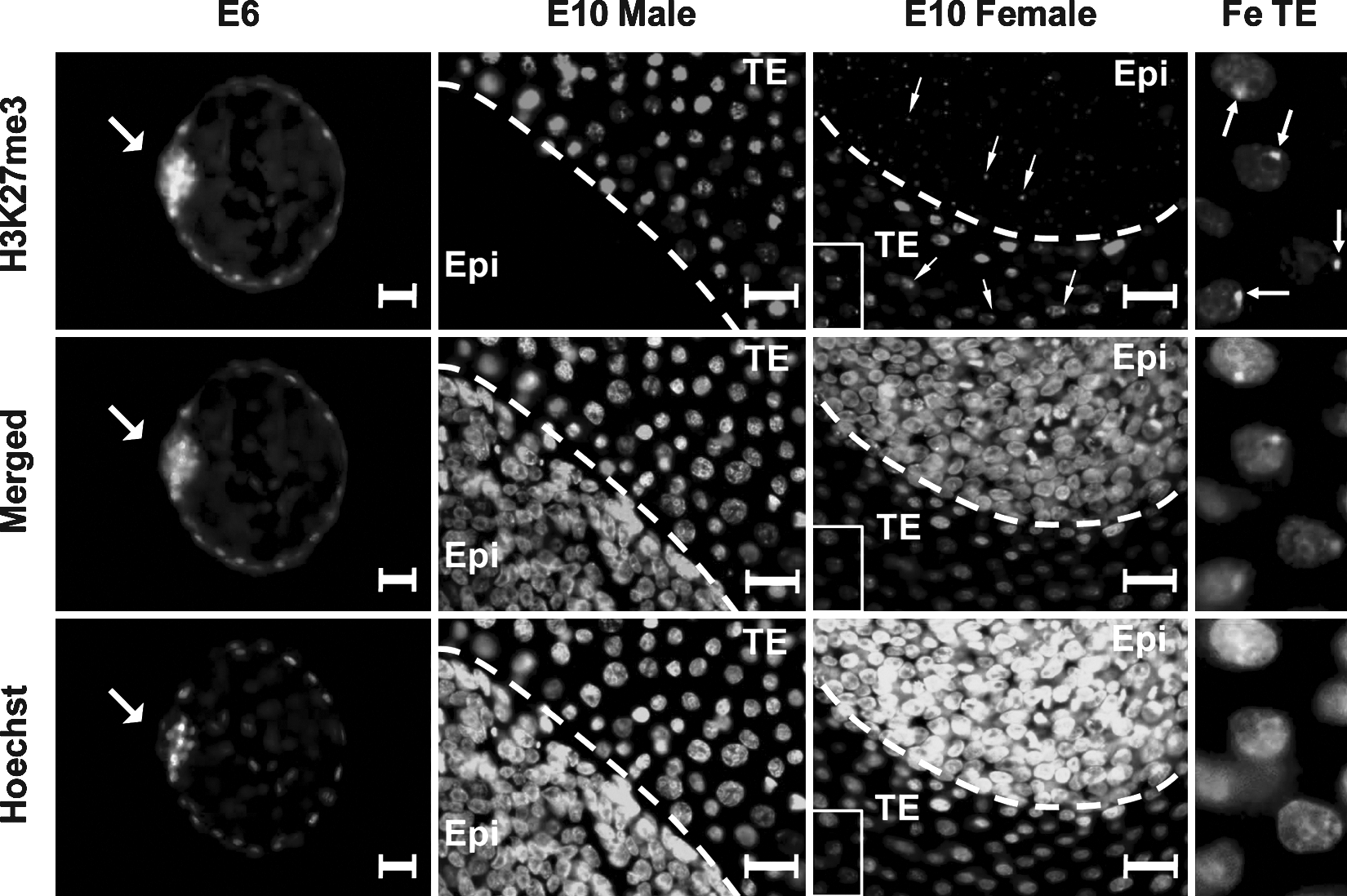
H3K27me3 immunostaining in the unsexed porcine E6 embryo, and in the male and female (Fe) porcine E10 epiblast (Epi), and trophectoderm (TE). Arrowhead points to the inner cell mass (ICM) in the E6 embryo and to single small spots visible in each cell of the epiblast and TE of female E10 embryos. Scale bar = 30 (m. Embryos are counterstained with the DNA labeling dye, Hoechst. Merged images show the overlay of the Hoechst and H3K27me3 images.
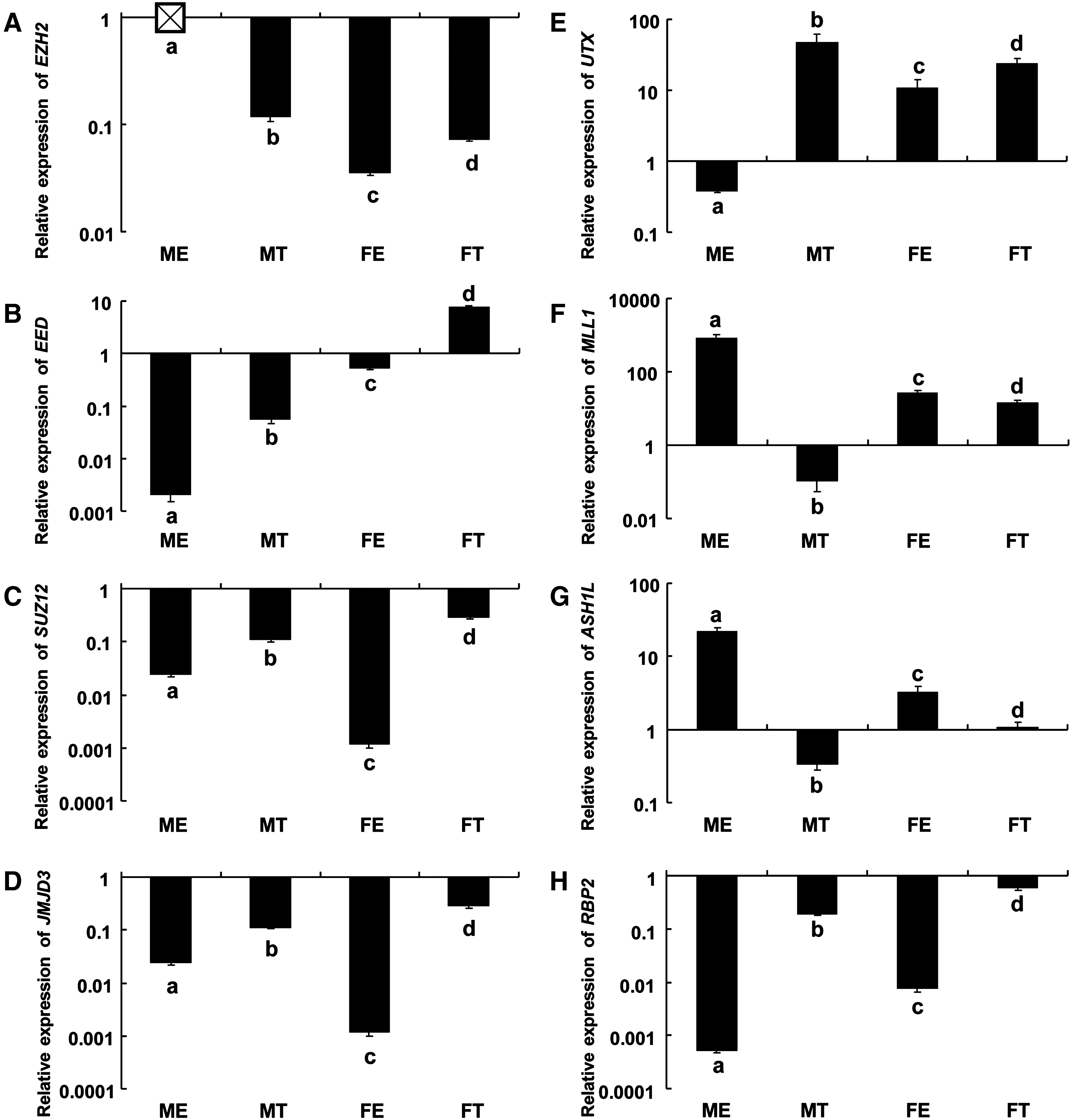
Expression of the H3K27me3 methylase (EZH2), cofactors (EED, SUZ12), demethylases (JMJD3, UTX), and H3K4me3 methylases (ASH1L, MLL1) and demethylase (RBP2) in E10 sexed porcine epiblast and trophectoderm (TE). Relative expression of (
The activation mark, H3K4me3, is exclusively hypermethylated in the E6 ICM
In order to assess the activation effects of H3K4me3 during lineage segregation, we performed immunolabeling against H3K4me3 in E6 and E10 porcine embryos. In the E6 embryos, H3K4me3 was exclusively hypermethylated in the nuclei of the ICM and not in the TE (Fig. 5). Use of a preabsorptive peptide revealed unspecific labeling in the cytoplasmic area in the later stage (E10) of development, however, due to lack of unspecific labeling in the nuclei, we believe this antibody to be nuclear-specific (Supplementary Fig. S3). By use of relative realtime PCR, we also evaluated a number of known methylases (ASH1L, MLL1) and demethylase (RBP2) in both sexed tissues at this stage of development. We detected high expression of H3K4me3 methylases MLL1 and ASH1L, specifically in the epiblast compared to the TE (Fig. 4F and G), and these levels were considerably higher in the male epiblast compared to the female epiblast. The H3K4me3 demethylase RBP2 was expressed at low levels in the epiblast and at high levels in the TE (Fig. 4H).
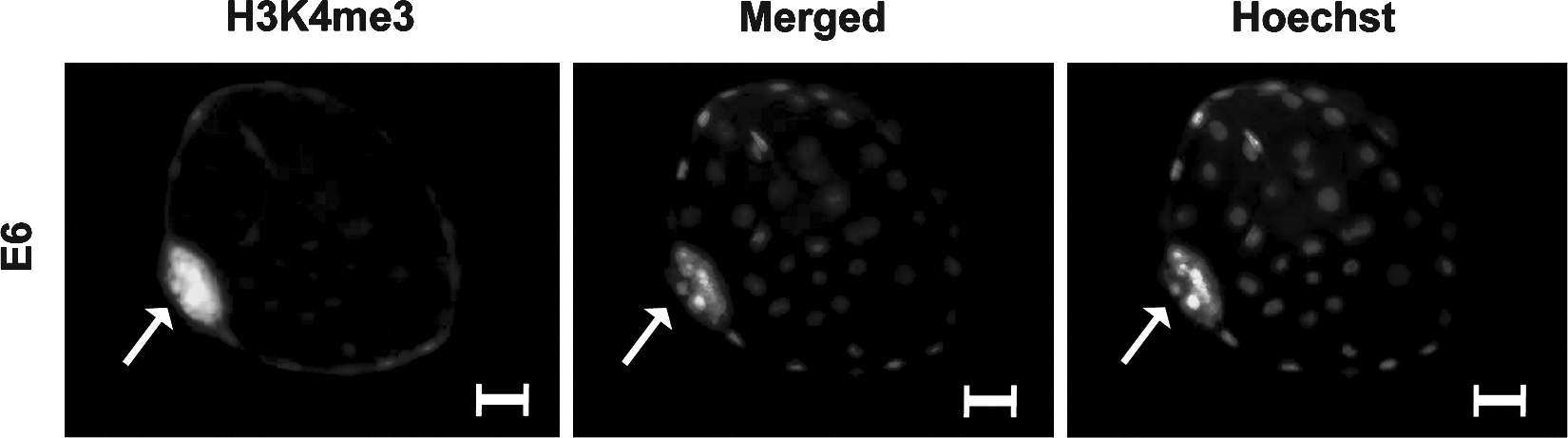
H3K4me3 immunostaining in the porcine nonsexed E6 blastocyst including the inner cell mass (ICM) and the trophectoderm (TE). Arrowhead points to the ICM and scale bar = 30 (m. Embryos are counterstained with the DNA labeling dye, Hoechst. Merged images show the overlay of the Hoechst and H3K4me3 images.
Modulation of heterochromatic packing is higher in the female epiblast and deacetylation events are not obvious in the porcine E10 embryo
In order to investigate heterochromatin packing, we evaluated methylases of H3K9 (G9A and SUV39H1) at the E10 stage of development. The expression of SUV39H1 was low in both the epiblast and TE of male and female embryos, although found to be significantly higher in the female epiblast. In contrast, the expression of G9A was very high (15∼277 times higher than SUV39H1) (Fig. 6A and B). The expression of G9A was particularly high in the female epiblast compared to the other tissues and significantly higher than the male epiblast (Fig. 6B).
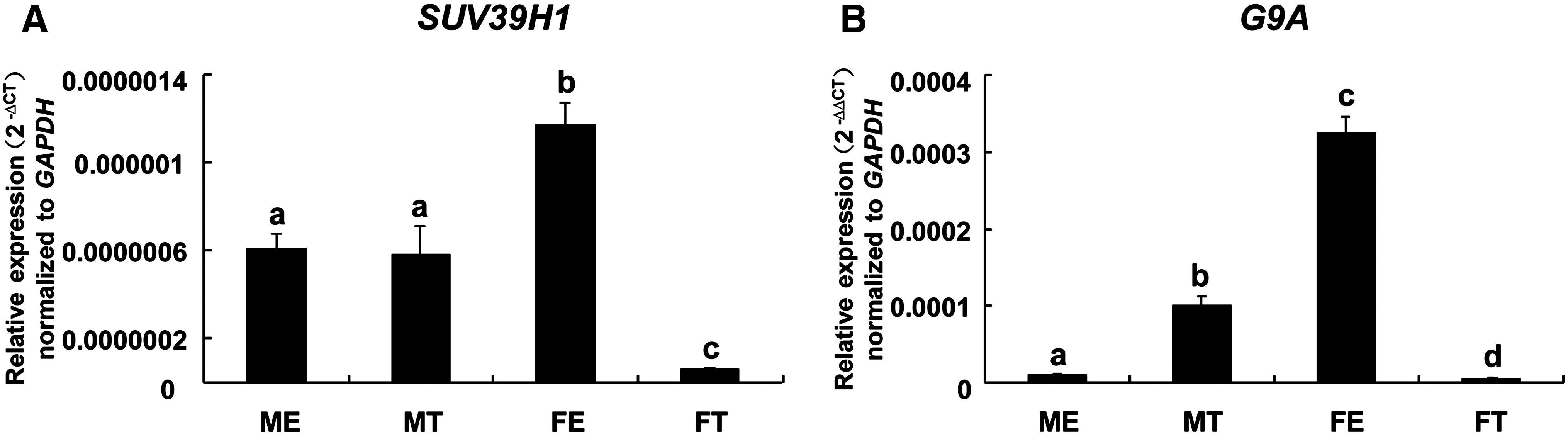
Expression of the H3K9 methylases, (
In order to evaluate deacetylation events in the developing porcine embryo, we investigated the expression levels of the histone deacetylases, HDAC1 and HDAC2. In general, a low expression of both genes was detected in the epiblast and TE, compared to levels observed in limb bud (E21) stage embryos. The expression of histone deacetylase HDAC1 differed between the TE and epiblast and also differed between male and female embryos (Fig. 7A), but in both male and female embryos, higher expression of HDAC2 was observed in the epiblast compared to the TE (Fig. 7B).
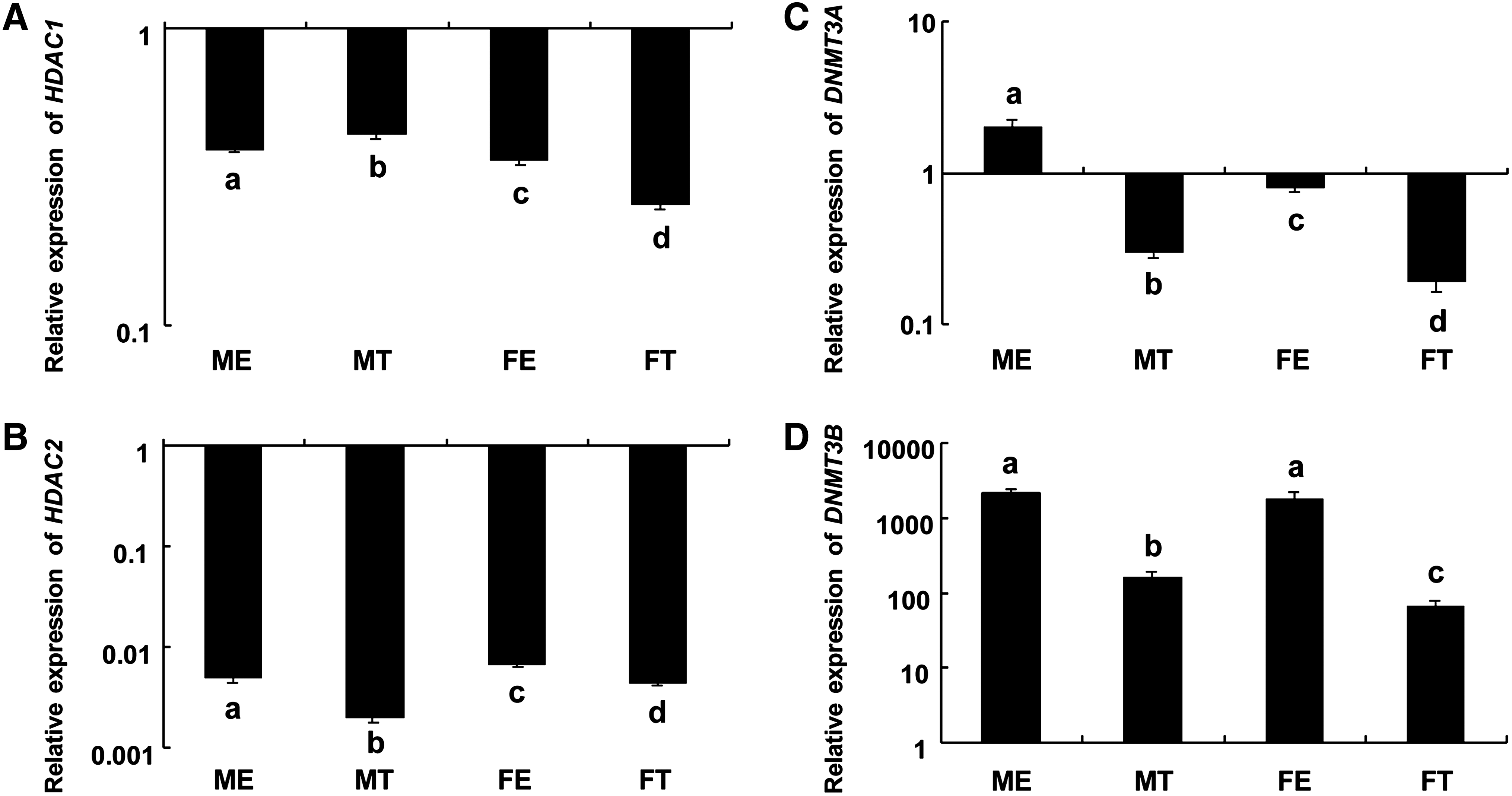
Expression of histone deacetylases (HDAC1 and HDAC2) and DNA methyltransferases (DNMT3A and DNMT3B) in sexed E10 porcine epiblast and trophectoderm (TE). Relative expression of (
An undistinct pattern of DNA methylation was found in the inner cell mass and trophectoderm of porcine E6 embryos, but a distinct pattern of DNA methylation was observed and correlated with the expression of DNA methyltransferases, DNMT3A, and DNMT3B in the epiblast and TE of E10 embryos
To investigate the role of DNA methylation in porcine lineage specification, we performed immunolabelling with an antibody against 5-methylcytidine in both E6 and E10 porcine embryos (validated with nonimmune IgG1 in porcine E10 TE) (Supplementary Fig. S4), and compared mRNA expression of DNA de novo methyltransferases, DNMT3A and DNMT3B, in the E10 porcine epiblast and TE by use of comparative realtime PCR. We observed expression of 5-meC in both the ICM and TE of E6 embryos, as well as in the epiblast and TE of E10 embryos (Fig. 8). The fluorescence intensity of 5-MeC in E6 ICM and TE did not differ, but was significantly different in E10 embryos, when compared with E6 embryos (p = 1.01 × 10−5) with a significantly higher methylation observed in the epiblast (Supplementary Fig. S5). The expression of DNA de novo methyltransferases DNMT3A and DNMT3B were significantly higher in the epiblast compared to the TE in E10 embryos (Fig. 7C and D). The level of DNMT3B expression was higher compared to DNMT3A, in both the epiblast and TE (Fig. 7C and D). In addition, the expression of DNMT3A was significantly higher in the male epiblast and TE compared to the female counterparts, and the expression of DNMT3B was significantly higher in the male TE compared to the female TE (Fig. 7C and D).
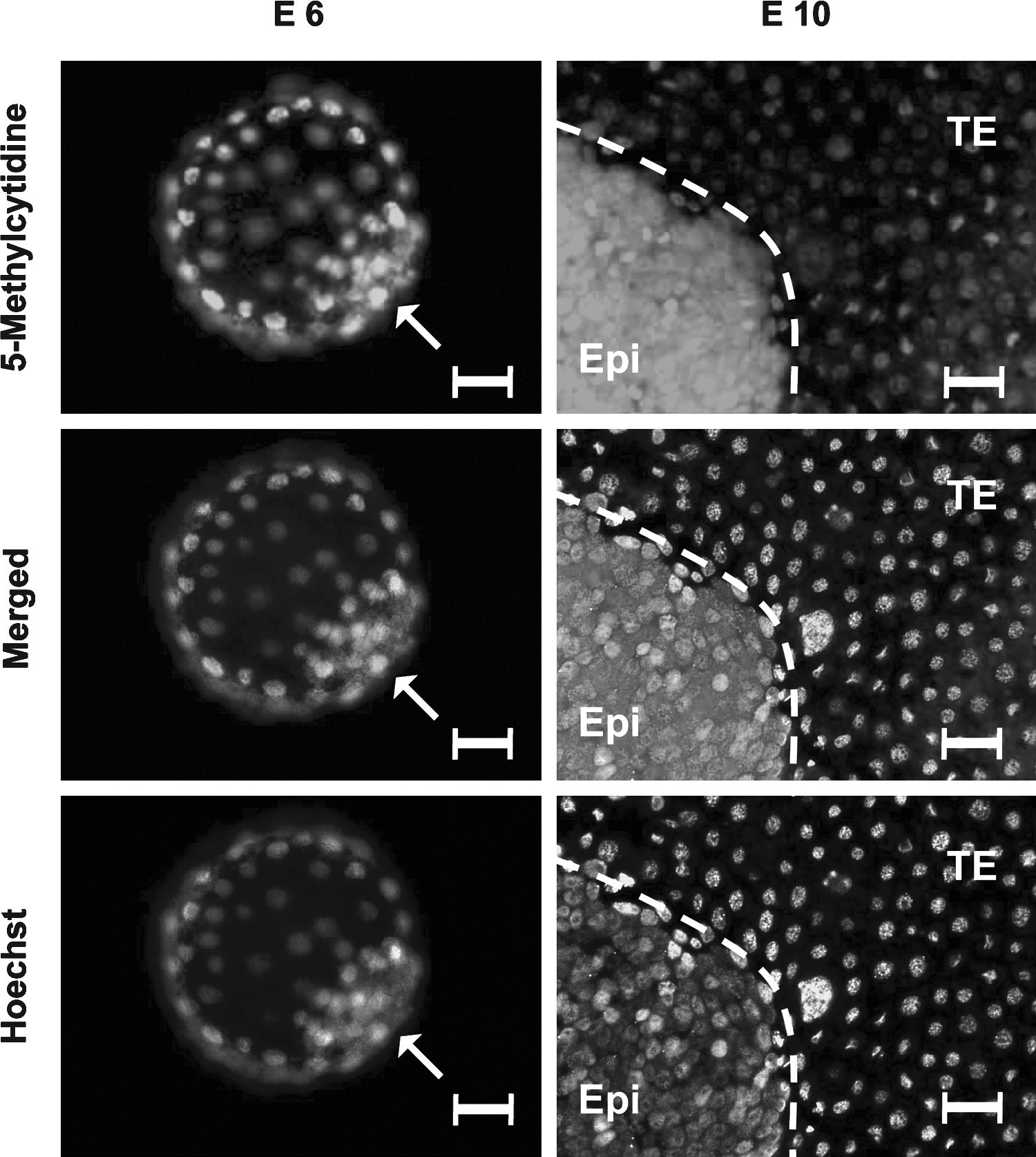
5-Methylcytidine immunostaining in porcine E6 blastocyst, containing the inner cell mass (ICM), and E10 blastocyst including the epiblast (Epi) and the trophectoderm (TE) with antibody against 5-Methylcytidine. Arrowhead points to the ICM and scale bar = 30 (m. Embryos are counterstained with the DNA labeling dye, Hoechst. Merged images show the overlay of the Hoechst and H3K4me3 images.
Discussion
In order to further understand the dynamics of gene expression and epigenetic marks involved in mammalian lineage specification and XCI during early development, we performed immunolabeling and comparative real-time PCR on sexed porcine preimplantation embryos, to evaluate patterns of histone modification and DNA methylation.
Distinct gene expression differences were observed in male versus female epiblast, which relate to lineage specification and early gastrulation, but not pluripotency, and could be accounted by a modulation of sex chromosomes on the genome machinery
In our data, we observed a high expression of XIST in the female porcine E10 embryos, which indicates that initiation of XCI has already occurred by this point of development. Xist expression has been observed in four-cell stage in mouse parthenogenetic embryos (Matsui et al., 2001), and as early as eight-cell stage in vitro derived bovine embryos (De La Fuente et al., 1999). In previous studies, the promoter regions of the pluripotent markers Oct4 and Nanog have been shown to be both permissive and repressive for a number of epigenetic mechanisms in both the mouse ICM and embryonic stem cells (Hattori et al., 2007; O'Neill et al., 2006). In analysis of pluripotent genes in the E10 porcine embryo, we found that the expression levels of OCT4, NANOG, and SOX2 differed in the epiblast. Interestingly, OCT4 expression was the highest, followed by SOX2 and a much lower expression of NANOG. Research suggests that levels of NANOG can vary considerably in embryonic stem cells (Chambers et al., 2007), but generally, low, or no NANOG in ESCs, leads to increased susceptibility of differentiation (Mitsui et al., 2003). Also, the expression of C-MYC was very low in the epiblast. Therefore, we speculate that low NANOG and possibly C-MYC, may be one indication for the difficulties in obtaining ESCs from the porcine embryonic epiblast. Our previous research also indicates that NANOG expression differs in the developing porcine embryo compared to human and mouse (Hall, 2008). It is likely that gene regulation differs in the porcine. Transcriptome profiling of the ICM and epiblast may be one way of discovering the genetic regulation important for stem-ness in this species. Culture of epiblasts in large throughput in vitro chemical screenings could be another approach for determining which factors/molecules may be required for maintaining pluripotency in vitro. This approach has been recently used to identify compounds for identification of small molecules and regulatory pathways in embryonic stem cells (Abujarour et al., 2010; Xu et al., 2010; Zhou et al., 2010). Previous research has also shown that overexpression of Oct4 inhibits Cdx2 expression, while increasing expression of Cdx2 reduces the expression of Oct4 (Niwa et al., 2005). Consistent with this observation, we observed high expression of OCT4 and low expression of CDX2 in the porcine epiblast. In mouse embryos, the primitive endoderm lineage has also been hypothesized to result from mutual antagonism between Nanog and Gata6 (Ralston et al., 2005). In line with this, we observed higher expression of NANOG and low expression of GATA6 in the porcine epiblast, and no expression of NANOG, but higher expression of GATA6 in the TE.
A striking observation was the higher levels of both CDX2 and GATA6 in female epiblast compared to male epiblast. In this tissue, the genes were not as well repressed as in the male. Similarly, the expression of EOMES and T were also repressed in the female epiblast compared to the male counterpart. We could speculate that these gene expression differences in the porcine blastocyst may be influenced either directly or indirectly by the sex chromosomes. Gene expression differences have previously been shown to differ in bovine sexed blastocysts (Bermejo-Alvarez et al., 2010). In this study, almost one-third of the detected transcripts (a total of 9322 transcripts were detected) showed sexual dimorphism in day 7 in vitro-produced blastocysts (Bermejo-Alvarez et al., 2010). We also observed a significantly higher expression of EED in the female epiblast (methylase cofactor of H3K27me3), which corroborates previous research showing EED to be an essential PcG protein required for protecting the reactivation of XCI (Kalantry et al., 2006). Interestingly, other significant alterations were also observed between male and female epiblast for the other methylases and demethylases of both H3K27me3 (i.e., EZH2, SUZ12, JMJD3, UTX) and H3K4me3 (MLL1, ASH1L, RBP2), which suggest that subtle differences in repression and activation of the histone marks may lead to differences in the level of expression of impacted genes. Interestingly, given that OCT4 and SOX2 levels did not differ between the sexes, we wonder if there is some protective role by other molecules that may preserve the temporally short, but critical, undifferentiated state.
Erasure of the H3K27me3 globally in the porcine ICM might be required for epiblast derivation, and the sexual dimorphism pattern of H3K27me3 in the E10 epiblast is likely linked with X chromosome inactivation
Our results clearly indicated that a segregation in the expression of H3K27me3 between the different cell types was not observed in E6 embryos, and only in E10 embryos. This is in accordance to our previous research (Gao et al., 2010). Erasure of H3K27me3 may be required for epiblast derivation, although an alternate mechanism may also occur, which results in subsequent loss of this methylation mark. However, what we could observe from sexed embryos, was the presence of small punctate spots of H3K27me3 in the female epiblast nuclei. In a previous study, female bovine TE cells from in vitro fertilization (IVF) day 7 bovine blastocysts, contained such spots, which were absent in each of the male TE cells (Breton et al., 2010). H3K27me3 has also been shown to colocalize to the inactive X chromosome expressing Xist in the mouse blastocyst (Okamoto et al., 2004). Therefore, we believe these spots may putatively indicate Xi, and represent the inactive X chromosome, which is repressed by H3K27me3, which has been shown previously to be the case in embryonic stem cells (Rougeulle et al., 2004).
In the case of the expression of the PcG genes, we were able to analyze their expression in both the epiblast and the TE and also in different sexed embryos. The expression of the methylases correlated very well to the observed H3K27me3 pattern, with higher expression in the TE compared to the epiblast. However, this was not the case, with respect to the demethylases (UTX and JMJD3), where expression was also higher in the TE. We speculate that demethylases might contribute to balance H3K27me3 levels in the embryos, because histone modification is a very dynamic event. Higher expression of both H3K27me3 methylases and demethylases in the TE than in the epiblast indicates that the histone mark H3K27me3 in the TE is more dynamic than in the epiblast to regulate gene expression during epiblast derivation. However, further studies are required for revealing the exact role of the demethylases of H3K27me3 in the embryos.
Distinct pattern of H3K4me3 in the E6 blastocyst
There has been very limited research investigating the expression and dynamics of H3K4me3 in the developing mammalian embryo. One previous study has reported that methylation of H3K4 can be observed in mouse early development, and specifically, in both the ICM and TE (Sarmento et al., 2004). In our study, we analyzed in detail the trimethylation of this histone mark in the porcine blastocyst containing the ICM and TE. Herein, we can confirm that, as in the mouse embryo, H3K4me3 expression is exclusive in the porcine E6 ICM. We believe this might indicate an important role of H3K4me3 in lineage segregation into the ICM and TE. In the early hatched blastocyst containing the early epiblast, the global level of H3K4me3 is lower in the epiblast than in the correspondent TE (Gao et al., 2010). A more unspecific antibody labeling is detected in the later E10 embryo, specifically in the cytoplasm (confirmed by peptide conjugate control). However, we can observe specific nuclear expression in the E10 TE. Interestingly, our examination of H3K4me3 methylases and demethylases suggested in contrast to the antibody immunolabeling, that H3K4me3 may be higher in the epiblast than the TE (i.e., higher MLL1 and ASH1L and lower RBP2 in the E10 epiblast). Upon further evaluation, ASH1L is found also to be a methylase of H3K9 and H4K20 (Beisel et al., 2002; Gregory et al., 2007); besides, ASH1L, MLL1, and RBP2 are only three of those well-identified H3K4me3 methylases and demethylase in mammal and there are over 10 different mammalian H3K4me3 methylases, such as Mll1-4, Set1a/b, Ash2L, Set7/9, Meisetz, Smyd3, etc., as well as many demethylases, such as Jhdm1a/b, Jarid1b/c/d, and Rbp2 etc (Hublitz et al., 2009). In addition, ASH1L, MLL1, and RBP2 might be associated with lineage segregation, but it is difficult for us to make definitive conclusions regarding the regulation of H3K4me3 in the E10 embryos. Further research is required to investigate H3K9 and H4K20 methylation and other H3K4me3 methylases and demethylases at E10 to gain more insight into the epigenetic regulation events in the porcine epiblast and TE.
Roles of H3K9me2, H3K9me3, and acetylation in embryonic development of E10 porcine embryos
Following the investigation of two methylases of H3K9me3 in E10 embryos, we observed a generally higher level of expression of G9A in E10 embryos compared to SUV39H1. Of particular note, both methylases were more highly expressed in the female epiblast compared to the other tissues. From previous research, an association between H3K9me2 and its role in potentially protecting against DNA methylation has been reported in the female pronucleus (Santos et al., 2005). It could be postulated that these genes may be involved in an important mechanism relating to XCI, occurring specifically in the epiblast. G9A has been suggested to also play a role in nonmethyltransferase activity, or may perform H3K9 methylation in a development-specific manner (Chin et al., 2007). Further investigation is required in order to determine the exact function of G9A and SUV39H1 expression in the female epiblast. Based on our own observations, we suggest that global levels of both H3K9me3 and H3K9me2 might not be involved in lineage segregation.
Ubiquitous expression of HDAC1/HDAC2 is observed in a variety of species during embryogenesis, but the levels of mRNA and protein vary between different organisms and embryonic stages (Brunmeir et al., 2009; Fischle et al., 1999). In this study, we found that both HDAC1 and HDAC2 were expressed at a low level in both the epiblast and TE of the E10 embryos. This finding may indicate that the deacetylases might contribute very little to the gene repression at this stage of development, or may be reflective of the less complex differentiation events occurring. It also suggests that acetylation events are likely high, which has been shown previously in the mouse, where hypoacetylation of H3K9 has been reported and coupled to the mechanism of Xi (Okamoto et al., 2004).
Dynamic change and distinct patterns of DNA methylation in the porcine blastocyst might associate with the lineage differentiation
We were interested in further analyzing DNA methylation events around the time of lineage segregation. It has been reported that de novo DNA methylation occurs in the ICM rather than the TE in both mouse and bovine blastocysts, which is consistent with a higher DNA methylation of the ICM genome compared to the TE genome (Dean et al., 2001; Santos et al., 2002). According to our present results, no difference in the level of DNA methylation expression could be determined between the ICM and TE in the E6 embryos, but could be determined between the EPI and TE in E10 embryos. This is in accordance with previous research, which has shown no methylation differences exist between the ICM and TE in pig E5 and E6-E7 embryos (Deshmukh et al., 2011; Fulka et al., 2006), but differences have been observed between the ICM and TE in E7-E8 stage embryos (Fulka et al., 2006). We did, however, observe a high level in the expression of the DNA de novo methyltransferase DNMT3B in E10 embryos, which was significantly higher in the epiblast compared to the TE. This also substantiates our own previously published work showing high expression of DNMT3B in the E11 porcine epiblast (Hall et al., 2010), which could not be detected in the earlier E6 embryo. The higher level of expression in the epiblast for both DNMT3A and DNMT3B confirm previous studies that indicate that distinct DNA methylation patterns may exist between the epiblast and TE and that may further affect subsequent cell fate. Previous studies have also reported that high levels of DNA methylation are important in ESCs in order to repress differentiation associated genes (Fouse et al., 2008). Similarly, in mouse embryos, a lack of DNMT expression results in differentiation of the ICM into extraembryonic tissues (Sakaue et al., 2010). Thus, high methylation is necessary for maintaining the stem cell state. Interestingly, the expression of both DNMT3A and DNMT3B showed sexual dimorphism, which is higher in the male E10 blastocyst than the female. This is in line with a previous observation in bovine day 7 blastocysts (Bermejo-Alvarez et al., 2008).
In the present study, we evaluated expression differences that differed slightly to our previous publication in pig embryos (Gao et al., 2010); that is, that HOXA7 and HOXA10 could not be detected in the epiblast, which was observed prior in the porcine hatched blastocyst and HOXA3 could now be detected in the TE of embryos in the present study. One reason for these differences may be due to differences in the developmental stages of the embryos used in both these studies. The stage of the embryos we used for the previous published study were designated to range from E8 to E10 (Gao et al., 2010), of which the developmental stage is up to 2 days earlier than the embryos used for the present study. Given the pools were more variable in the previous study, it could be possible that transcripts present in E10 embryos may not be detectable under a 35 cycle PCR, if more E9 or E8 embryos were present in the pool. Similarly, in the present study, we may not be able to identify the expression of genes that may be downregulating at either E8 or E9. One other possible reason to explain these differences is that we used the whole blastocyst (containing the epiblast, TE and hypoblast) for investigating the Hox gene expression in the previous study (Gao et al., 2010). In the present study, we manually dissected the TE and epiblast for profiling Hox gene expression and removed the hypoblast, this giving a more clear indication of the spatial expression of these genes in the different cell types of the embryo. Our research also indicates that no HOX genes were exclusively detected in the E10 epiblast, but HOXA3 and HOXD8 were exclusively expressed in the TE. The expression of these homeobox genes in the TE, have not been previously described and their role remains elusive. Considering trxG proteins are required to keep Hox genes active (Klymenko and Muller, 2004), it could be possible that the demethylases, JMJD3 and/or UTX, which we observed to be highly expressed in the TE, may play a potential role in the regulation of HOXA3 and HOXD8 and other possible HOX genes. Further studies are required to examine the epigenetic regulation of Hox genes during preimplantation development.
In conclusion, our study attempts to highlight the epigenetic events coupled to the preimplantation porcine blastocyst containing the ICM and the epiblast. It is important to understand potential epigenetic and gene expression differences between the pig compared to the mouse and human, which may help to indicate why bona fide embryonic stem cells are difficult to derive in this species. We have studied epigenetic mechanisms in E10 embryos, including histone trimethylation, DNA methylation, and epigenetic events potentially related to XCI. Differences observed from this study include a low expression of NANOG and C-MYC in the porcine epiblast. These genes are important in the regulation of mouse and human ESCs and induced pluripotent stem cells. The lower expression of these genes may be a clue to differences in gene regulation in the porcine epiblast. Interestingly, we also discovered a large number of genes differed in their expression level between the male and female epiblast, which included genes such as CDX2, GATA6, EOMES, and T, as well as genes that regulate methylation, such as EED, SUV39H1, MLL1, ASH1L, HDAC1, and DNMT3A. Thus, gene expression differences can be observed in the porcine epiblast compared to other species. In addition, genes regulating epigenetic events such as H3K27me3 and global methylation differ between porcine male and female embryos and are likely attributable to sexual dimorphic events such as XCI.
Footnotes
Acknowledgments
We acknowledge the financial support of EU projects, from the 7th Framework, PluriSyS (223485), and PartnErS (218205), as well as from the Danish National Advanced Technology Foundation, Pigs, and Health project, in Denmark, and from the National Natural Science Foundation of China (30700574).
Author Disclosure Statment
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.