
Abstract
Abstract
Somatic cell nuclear transfer (SCNT) has had an enormous impact on our understanding of biology and remains a unique tool for multiplying valuable laboratory and domestic animals. However, the complexity of the procedure and its poor efficiency are factors that limit a wider application of SCNT. In this context, oocyte meiotic arrest is an important option to make SCNT more flexible and increase the number of cloned embryos produced. Herein, we show that the use of butyrolactone I in association with brain-derived neurotrophic factor (BDNF) to arrest the meiotic division for 24 h prior to in vitro maturation provides bovine (Bos indicus) oocytes capable of supporting development of blastocysts and full-term cloned calves at least as efficiently as nonarrested oocytes. Furthermore, the procedure resulted in cloned blastocysts with an 1.5- and twofold increase of POU5F1 and IFNT2 expression, respectively, which are well-known markers of embryonic viability. Mitochondrial DNA (mtDNA) copy number was diminished by prematuration in immature oocytes (718,585±34,775 vs. 595,579±31,922, respectively, control and treated groups) but was unchanged in mature oocytes (522,179±45,617 vs. 498,771±33,231) and blastocysts (816,627±40,235 vs. 765,332±51,104). To our knowledge, this is the first report of cloned offspring born to prematured oocytes, indicating that meiotic arrest could have significant implications for laboratories working with SCNT and in vitro embryo production.
Introduction
When the oocyte is removed from the follicular environment it spontaneously resumes the meiotic division progressing into a mature oocyte (Kotsuji et al., 1994). This occurs because the oocyte is released from the influence of meiosis inhibiting factors as reviewed by Tripathi et al. (2010). However, spontaneous meiosis resumption can be avoided if the oocyte is prematured in the presence of specific cell cycle inhibitors (i.e., butyrolactone I) after removal from the follicle (Adona and Leal, 2004; Kubelka et al., 2000). Butyrolactone I is a selective inhibitor of maturation promoting factor (MPF) activity through cyclin-dependent kinase (CDK) inhibition (Knockaert et al., 2002) that has been successfully employed to arrest meiotic progression in several species without harming preimplantation development (Adona and Leal, 2004; Adona et al., 2008a, 2008b; Hashimoto et al., 2002; Kubelka et al., 2000). Prematuration in the presence of low concentrations of butyrolactone I (i.e., 10 μM), but in the absence of bovine serum albumin (BSA), arrests over 97% of bovine oocytes at the germinal vesicle (GV) stage of meiosis and has no effect on metaphase II (MII) formation rate after maturation when compared to controls (Adona et al., 2008b).
Oocyte developmental competence is acquired before maturation while the gamete undergoes several changes at both structural and molecular levels (Blondin et al., 1997; Mermillod et al., 1999). These have been termed oocyte capacitation and are essential to support embryonic development, whereas the embryo relies on maternal factors (Blondin et al., 1997). Prematured oocytes can be kept arrested at the GV stage of the meiotic division for up to 48 h before being subjected to in vitro maturation (IVM) (Adona and Leal, 2004; Coy et al., 2005a; Galli et al., 2003a; Lagutina et al., 2002; Ponderato et al., 2002). Thus, in vitro prematuration (IVPM) has been suggested as a mean to provide the oocyte with additional time to acquire developmental competence prior to IVM (Hashimoto et al., 2002; Hendriksen et al., 2000). This can be accomplished by the use of specific compounds that exert a positive effect on oocyte competence. For instance, addition of brain-derived neurotrophic factor (BDNF) (Kawamura et al., 2005; Seifer et al., 2003) during IVPM has been suggested to improve oocyte competence (Adona et al., 2011). In addition, IVPM has the advantage of enabling more flexibility to the procedure of in vitro embryo production by allowing adjustments to the time of IVPM–IVM according to the needs of the laboratory (Galli et al., 2003a; Lagutina et al., 2002). This is particularly relevant to SCNT because it requires a long period of oocyte manipulation (i.e., microsurgery).
The feasibility of IVPM to produce viable offspring has been shown by in vitro fertilization (IVF) using pig oocytes (Coy et al., 2005b). Regarding the poor efficiency of SCNT to produce viable offspring, it remains to be shown that oocyte produced by IVPM can be used to produce cloned animals by SCNT. Thus, this work was aimed at testing the hypothesis that prematured oocytes can support development to term after SCNT.
Material and Methods
Unless otherwise stated, all chemical and reagents were purchased from Sigma-Aldrich Chemical Co. (St. Louis, MO), and in vitro experimental procedures were conducted in humidified incubators maintained at 38.5°C with 5% CO2 in air.
Oocyte collection
Oocytes were obtained postmortem from the ovaries of cows (Bos indicus) slaughtered at a local abattoir. Ovaries were transported in 0.9% saline solution with 100 U/mL penicillin and 100 U/mL streptomycin at 25–30°C to the laboratory. The ovaries were washed in saline solution, and follicles with diameter between 3 and 6 mm were aspirated with an 18-G needle attached to a 10-mL syringe. The follicular fluid aspirated was placed in a 50-mL centrifuge tube. The decanted material was placed in a 60-mm plastic dish with HEPES-buffered tissue culture medium-199 (TCM-199; GIBCO BRL, Grand Island, NY) supplemented with 10 μg/mL gentamycin and 5% fetal bovine serum (FBS; GIBCO BRL). Oocytes with homogeneous cytoplasm and at least three layers of cumulus cells were selected and distributed randomly among the treatments.
IVPM
IVPM was performed as previously reported (Adona et al., 2011). Briefly, butyrolactone I (Biomol Research Laboratories, Plymouth Meeting, PA) was diluted at 50 mM in dimethyl sulfoxide (DMSO) and stored in aliquots at −20°C. Oocytes were incubated with butyrolactone I at 10 μM in bicarbonate-buffered TCM-199 (GIBCO BRL) supplemented with 10 μg/mL gentamycin and 0.2 mM sodium pyruvate in 100 μL droplets (15–20 oocytes per droplet) under mineral oil for 24 h. Oocytes were then subjected to IVM as described below.
IVM
Oocytes were subjected to IVM as previously reported (Adona et al., 2011). IVM was performed in bicarbonate-buffered TCM-199 with 10% (v/v) FBS, 5 μg/mL luteinizing hormone (LH; Lutropin-V, Vetrepharm, London, Canada), 0.5 μg/mL follicle stimulating hormone (FSH; Folltropin-V, Vetrepharm), 0.2 mM sodium pyruvate, and 10 μg/mL gentamycin in 100 μL droplets (15–20 oocytes per droplets) under mineral oil for 24 h. Afterward, oocytes were either denuded, fixed, and stained for analysis of nuclear maturation and first polar body (PB) extrusion, or assessed for developmental competence through parthenogenetic activation or SCNT. Nuclear maturation was analyzed as reported by Adona and Leal (2004) using lacmoid staining and classified as GV, MI, anaphase I, telophase I, or MII stage.
Developmental assessment after parthenogenetic activation
After IVM, oocytes were denuded of cumulus cells and selected for the presence of the first PB. Selected oocytes were chemically activated (at 26 h post-IVM) by incubation in 5 μM ionomycin in HEPES-buffered TCM-199 supplemented with 0.1% BSA (fatty acid-free), 0.2 mM sodium pyruvate and 10 μg/mL gentamycin for 5 min, followed by incubation in HEPES-buffered TCM-199 with 3% BSA for 1 min and in 2 mM 6-dimethylaminopurine (6-DMAP) diluted in modified synthetic oviduct fluid (SOFaa) (Holm et al., 1999) for 3 h. Oocytes were then extensively washed in SOFaa and in vitro cultured in groups of 15–20 in 100 μL droplets of SOFaa on a monolayer of cumulus cells for 192 h. Part of the activated oocytes were, instead of cultured, used for pro-nuclear analysis by staining for 15 min in 10 μg/mL Hoechst diluted in phosphate-buffered saline (PBS) with 0.1% (w/v) polyvinylalcohol (PVA) in the dark. After washing twice in PBS with 0.1% PVA, oocytes were mounted on slides with coverslips using VectaShield Mounting Medium (Vector Laboratories Inc., Burlingame, CA) and visualized on a fluorescence microscope (Axioplan; Carl Zeiss, Zeppelingstrasse, Germany). On the other hand, cultured embryos were assessed for developmental rates at day 2 [cleavage at 48 h postparthenogenetic activation (48 hpa)], day 7 (blastocysts at 168 hpa), and day 8 (blastocysts and hatched blastocysts at 192 hpa). Cleavage and blastocyst rates were reported in relation to the number of presumptive zygotes placed in culture, whereas hatched blastocyst rate was reported in relation to the number of blastocysts at day 7. Moreover, blastocysts at day 8 were evaluated for the total number of cells and number of DNA-fragmented cells.
SCNT
Somatic cell nuclear transfer was performed as Miranda et al. (2009) with few modifications. Briefly, skin fibroblasts from four cultures from Nellore (B. indicus) animals (three females and one male) were used as a source of nuclear donors. Fibroblasts were in vitro cultured in Dulbelcco's modified Eagle's medium (DMEM; GIBCO BRL) supplemented with 10% FBS and 10 μg/mL gentamycin, and maintained in a humidified incubator at 38.5°C with 5% CO2, 5% O2, and 90% N2. Fibroblasts between third and fifth passages were grown to confluence and arrested at the G1/G0 stages of the cell cycle via contact inhibition to be used as nuclear donors. Fibroblasts from a single culture in the same cell passage and at the same confluence level were used as nuclear donors for all treatment groups in a single experimental routine.
Matured oocytes used as recipient ooplasts for SCNT were obtained after selection for the presence of the first PB as described above, after 18 h post-IVM. These oocytes were incubated in groups of 20–30 in SOFaa containing 10 μg/mL Hoechst and 7.5 μg/mL cytochalasin B for 15 min. For microsurgery, the group was transferred to a 400-μL droplet of HEPES-supplemented SOFaa with 7.5 μg/mL cytochalasin B. Microsurgery was performed using an inverted microscope (Leica DMI RB; Leica, Wetzlar, Germany) equipped with micromanipulators and microinjectors (Narishige, Tokyo, Japan). The first PB and MII plates were removed by aspiration with a 15-μm inner diameter glass pipette (ES transferTip; Eppendorf, Hamburg, Germany). First PB and MII removal was confirmed by exposure of the biopsy to ultraviolet light.
Enucleated oocytes were reconstructed by injection of a single fibroblast into the perivitelline space using the same micromanipulation system described above. The resulting couplet was placed in a fusion chamber (Eppendorf) filled with electrofusion solution (0.28 M mannitol, 0.1 mM MgSO4, 0.5 mM HEPES, and 0.05% BSA in water) and subjected to one pulse of alternating current (0.05 kV/cm for 5 sec) and two pulses of continuous current (1.75 kV/cm for 45 μsec) to promote fusion between the somatic cell and the recipient ooplast. Successfully fused couplets were activated and in vitro cultured as described above. Parthenogenetic embryos were used as controls. These experiments intended to produce embryos for analysis of developmental rates (see above), number of nuclei in blastocyst (day 8), TUNEL (day 8), mtDNA copy number (day 8), gene expression (day 8), or embryo transfer (day 7).
Embryo transfer into recipient cows
Blastocysts were individually transferred nonsurgically into the uterus of previously synchronized recipient cows (Nasser et al., 2004). These recipient cows were evaluated for pregnancy by ultrasound visualization of fetal heart beat on days 35 and 60; the pregnancies were allowed to develop to term.
Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL)
The number of blastocyst DNA-fragmented cells and total nuclei were determined as reported by Miranda et al. (2009). Briefly, cells with DNA fragmentation were detected based on the TUNEL assay using the In situ Cell Apoptosis Detection Kit (Roche Molecular Biochemicals, Mannheim, Germany). Blastocysts were washed in 0.1% PVA in PBS and fixed for 1 h at room temperature in 3.7% (w/v) paraformaldehyde diluted in PBS with 0.1% PVA. Blastocysts were then washed in PBS with 0.1% PVA and permeabilized in PBS containing 0.5% (v/v) Triton X-100 and 0.1% (w/v) sodium citrate for 1 h at 4°C. Positive controls were made by incubation for 1 h at 37°C in 50 U/mL DNAse I solution (Invitrogen, Carlsbad, CA). Afterward, blastocysts were washed thrice in PBS with 0.1% PVA and incubated in the dark for 1 h at 37°C in a humidified chamber in fluorescein isothiocyanate-conjugated dUTP plus the enzyme terminal deoxynucleotidyl transferase. Negative controls were made by incubation in the absence of the enzyme. Blastocysts were washed thrice in PBS with 0.1% PVA and stained in the dark for 15 min in 10 μg/mL Hoechst diluted in PBS with 0.1% PVA. After washing twice in PBS with 0.1% PVA, blastocysts were mounted on slides with coverslips using VectaShield Mounting Medium and visualized on a fluorescence microscope (Axioplan). Total blastocyst cell nuclei and TUNEL-positive nuclei were counted using the AxioVs40 software (V4.6.1.0; Carl Zeiss).
Absolute quantification of mtDNA
Oocytes used for mtDNA analysis were denuded of cumulus cells by vortexing, extensively washed in PBS with 0.1% of PVA, and stored at −80°C individually in 0.2 mL polystyrene PCR tubes with 10 μL of PBS with 0.1% of PVA. Blastocysts used for mtDNA analysis were simply washed thrice in PBS with 0.1% PVA and stored as described for oocytes. For quantification of mtDNA, samples were lysed as carried out by Wai et al. (2008). Briefly, samples were subjected to three rounds of heating (at 95°C for 5 min) and freezing (in liquid nitrogen for 3 min), and spun down to measure the final volume of the lysate.
A standard curve used for mtDNA quantification was prepared using a 703-bp fragment encompassing part of MT-RNR2, MT-TL1, and MT-ND1 genes. This fragment was obtained by PCR amplification in a 50-μL reaction containing 0.5 μM of standard primers (Table 1), 0.2 mM dinucleotide triphosphates (Invitrogen), 1.5 mM MgCl2 (Invitrogen), 1×PCR buffer (Invitrogen), 0.01 U/μL Taq DNA polymerase (Invitrogen), and 1 ng/μL of genomic DNA extracted from white blood cells (Chiaratti and Meirelles, 2006). The following cycling conditions were applied for amplification: initial denaturation at 95°C for 10 min, 40 cycles consisting of 95°C for 20 sec, 54°C for 30 sec, and 72°C for 1 min, and a final extension step at 72°C for 10 min. The PCR product was purified using the Wizard SV Gel and PCR Clean-up System (Promega, Madison, WI) and cloned using the TOPO TA Cloning kit (Invitrogen). The plasmid containing the mtDNA fragment was purified using the Wizard Plus SV Miniprep (Promega). The concentration of copies of plasmid DNA was determined using a Nanodrop 2000 spectrophotometer (Thermo Scientific, Wilmington, DE) and a stock solution prepared at 0.2×109 copies/μL. This stock solution was stored at −20°C in single-use aliquots until use.
Quantitative real-time PCR for mtDNA was performed as described by Chiaratti et al. (2010), with few modifications. Briefly, a TaqMan assay was designed to amplify an 87-bp fragment of the mitochondrial encoded 16S RNA (MT-RNR2), which is fully conserved between B. taurus and B. indicus (Table 1). Quantification was performed in an ABI PRISM SDS 7500 HT Real-Time PCR System (Applied Biosystems, Foster, CA). Reactions consisted of a total of 20 μL containing 0.9 μM of primer plus 0.25 μM of the TaqMan probe plus 1×TaqMan Gene Expression Master Mix (Applied Biosystems) plus 1 μL of template (sample or standard). Samples were always run in duplicates and each duplicate was averaged for quantification. Standard curves were always amplified in parallel to samples using five 10-fold serial dilutions (103–107 copies) of the standard. This TaqMan assay showed 100% of amplification efficiency. Based on the standard curve values, it was possible to quantify the starting copy number of mtDNA in each sample using the SDS software (V2.3; Applied Biosystems). The final value of mtDNA amount was corrected by the volume of the lysate.
Relative quantification of mRNA
The target genes of interest were the POU5F1 (previously known as OCT4) and the IFNT2 (interferon tau), and GAPDH (glyceraldehyde-3-phosphate dehydrogenase) was used as an endogenous control as suggested by Vigneault et al. (2007). Primers used for PCR amplification are presented in Table 1.
Blastocysts used for gene expression analysis were washed thrice in PBS with 0.1% PVA plus 1 U/μL ribonuclease inhibitor (RNAse OUT; Invitrogen), placed individually into 0.2-mL polystyrene microtubes containing 1 μL of PBS with 0.1% PVA and 1 U/μL RNAse OUT, snap frozen in liquid nitrogen, and stored at −80°C until use. The RNA was extracted from pools of six embryos using the RNeasy Protect Mini Kit (Quiagen, Hilden, Germany) according to the manufacturer's recommendations. Immediately after extraction the RNA was reversed transcribed into cDNA using SuperScript II (Invitrogen) according to the manufacturer's protocol and stored at −20°C until use.
Relative quantification of mRNA amounts was performed as reported by Chiaratti et al. (2010). Briefly, cDNA was preamplified using the TaqMan PreAmp Master Mix Kit (Applied Biosystems) and a mix of GAPDH, POU5F1, and IFNT2 primers at 45 nM each. Samples were subjected to 10 thermal cycles (95°C for 15 sec and 60°C for 4 min) and stored at −20°C until use. The linearity of preamplification of all transcripts was checked as suggested by the manufacturer. Afterward, relative quantification of gene-specific mRNA transcripts was done in 20-μL reactions containing 0.2 μM of primer (GAPDH, POU5F1, or IFNT2), 1×SYBR Green PCR Master Mix (Applied Biosystems) and 2 μL of template. For each sample, preamplified cDNAs were diluted seven-fold (POU5F1 and IFNT2) or three- to ninefold (GAPDH) to be used as template. All gene-specific cDNAs amplified for a particular sample were always run in the same PCR plate. The following cycling conditions were applied for amplification: initial denaturation at 95°C for 15 min followed by 40 cycles consisting of 95°C for 20 sec and 60°C for 1 min. The SYBR Green fluorescence was read at the end of each extension step (60°C). Standard curves were generated for each sample and for each gene-specific cDNA analyzed using seven-fold serial dilutions of samples. The specificity of each PCR product was checked by dissociation curves using the ABI PRISM SDS 7500 HT Real-Time PCR System and by loading the PCR products onto 2% agarose gels. The averaged amplification efficiency of each primer set was 97, 90, and 96%, respectively, for GAPDH, POU5F1, and IFNT2. Relative target transcript amounts in each sample were determined using the standard curve method (Livak and Schmittgen, 2001).
Experiment 1: Effect of BDNF Supplementation During IVPM and IVM
In this first experiment we sought to determine whether IVPM or IVM should be carried out in the presence of BDNF. BDNF was diluted at 1 μg/mL in bicarbonate-buffered TCM-199 and stored in aliquots at −20°C. It was added to IVPM or IVM medium at 10 ng/mL. As controls, two additional groups were assessed: IVPM or IVM without BDNF supplementation. After IVPM, oocytes were subjected to IVM. To evaluate the effect of BDNF during IVM, oocytes were not subjected to IVPM. Therefore, four groups were assessed for analysis of nuclear maturation (MII formation rate) and embryo quality after parthenogenetic activation: (1) IVM, (2) IVM with BDNF, (3) IVPM + IVM, and (4) IVPM with BDNF + IVM. Experiments were run in parallel between IVM and IVM + BDNF or IVPM and IVPM + BDNF. At least 80 oocytes per group had nuclear maturation assessed whereas a minimum of 440 oocytes per group were used for parthenogenetic activation. Embryo quality was assessed by developmental rates (cleavage, blastocyst, and blastocyst hatched rates) and, by the number of nuclei and TUNEL-positive nuclei (9–11 embryos per group) in parthenogenetic blastocysts. At least three experimental routines were carried out for each data collection.
Experiment 2: Developmental Competence of IVPM-Derived Oocytes in SCNT
Our aim in this experiment was to evaluate the developmental competence of prematured oocytes as recipient ooplasts for SCNT. Based on the results from the first experiment, oocytes were subjected to IVPM in the presence of 10 ng/mL BDNF followed by IVM in the absence of BDNF (treated group). As a control, an additional group was exclusively matured in the absence of BDNF. These two groups (control and treated, respectively) were used as recipient ooplasts for SCNT (1000 and 1006 oocytes per group, respectively) to assess their embryonic development (cleavage and blastocyst rate). Moreover, cloned blastocysts were assessed for the number of nuclei (15 blastocysts per group), TUNEL-positive nuclei (15 blastocysts per group), and expression of POU5F1 and IFNT2 genes (18 embryos per group). Parthenogenetic embryos cultured in parallel with cloned embryos (18 blastocysts per group) were used for gene expression analyzes as well. Finally, to further evaluate the effect of prematuration on embryos, cloned blastocysts were transferred to recipient cows (26 and 28 embryos, respectively for treated and control groups) to assess their ability to support development to term. At least three experimental routines were carried out for each data collection.
Experiment 3: Mitochondrial DNA Replication During IVPM
Our aim in this third experiment was to evaluate whether oocyte mtDNA copy number is increased by prematuration. Based on the results from Experiments 1 and 2, oocytes were subjected to IVPM in the presence of 10 ng/mL BDNF followed by IVM in the absence of BDNF. As a control group, oocytes were exclusively subjected to IVM (in the absence of BDNF). Oocytes were collected for mtDNA analysis (1) before being subjected to IVPM and IVM (immature oocytes—control); (2) after IVPM but before IVM (immature oocytes—treated); (3) after IVM but without being subjected to IVPM (mature oocytes—control), and (4) after IVPM and IVM (mature oocytes—treated). Moreover, mtDNA was quantified in cloned blastocysts produced using oocytes prematured in the presence of BDNF (blastocysts—treated) and oocytes subjected to IVM exclusively (blastocysts—control). The mtDNA content was compared between control and treated groups within immature oocytes, mature oocytes, and blastocysts, or among stages (immature oocytes, mature oocytes, and blastocysts) within group (control and treated). Thirty oocytes per group were assayed for mtDNA analysis. At least three experimental routines were carried out for each data collection.
Statistical analysis
Statistical analyses were performed using the SAS System (V8; Cary, NC). Data presented as percentage were analyzed using chi-square or Fisher's Exact tests. The number of nuclei, TUNEL-positive nuclei, and gene expression were analyzed using Student t-test. Mitochondrial DNA copy number was analyzed by one-way analysis of variance (ANOVA) followed by Tukey post hoc test. All experiments were repeated at least three times. Differences with probabilities (p)<0.05 were considered significant. The standard error of the mean (SEM) is presented for continuous values.
Results
Supplementation with BDNF during IVPM improves maturation and cleavage rates
First, we designed an experiment to determine whether the use of BDNF during IVPM or IVM improves oocyte maturation and embryonic development. The following data are representative of at least three experimental replicates. In this first experiment, when the effect of the drug was studied during IVM, oocytes were subjected exclusively to IVM (not to IVPM) in the presence or absence of BDNF. On the other hand, when the effect of drug was tested during IVPM, oocytes were subjected to IVPM in the presence of BDNF but to IVM in the absence of BDNF to compare with oocytes subjected to IVPM and IVM in the absence of BDNF. Thus, although the use of BDNF during IVM (77/98, 78.6%) had no effect (p=0.86) on MII formation rate compared to when IVM was performed in the absence of BDNF (76/98, 77.6%), its use during IVPM did increase (p=0.01) MII formation rate (73/80, 91.3%) compared to oocytes prematured in the absence of BDNF (66/86, 76.7%). Lacmoid staining of oocytes after follicle aspiration showed that 41 out of 44 oocytes were at the GV stage (three oocytes were classified as abnormal). These findings suggest that the increase in MII formation regarding IVPM was caused by supplementation with BDNF.
To investigate whether BDNF also affects the embryonic development, oocytes produced as described above were parthenogenetically activated and in vitro cultured to evaluate developmental rates. It is worth to note that comparisons were only made between groups that were run in parallel: IVM versus IVM + BDNF or IVPM versus IVPM + BDNF. A negative effect of BDNF was found (p=0.02) on pro-nuclear rates when the drug was added during IVM (30/42, 71.4%) compared to IVM not supplemented with BDNF (41/45, 91.1%). However, supplementation with BDNF during IVPM did not affect (p=0.19) pro-nuclear rates (25/39, 64.1%) compared to IVPM made in its absence (31/40, 77.5%). Although cleavage rates were superior (p=0.04) when adding BDNF exclusively during IVPM (65.2% vs. 57.7%), developmental rates to blastocyst were unaffected (p>0.05) by the addition of BDNF during both IVM and IVPM (Table 2). Blastocyst quality, evaluated by the hatched rate (Table 2), TUNEL-positive cells (Table 3) and total nucleus number (Table 3), did not show (p>0.05) an effect of BDNF regardless of its use during IVPM or IVM. In summary, these data suggest the use of BDNF during IVPM, but not during IVM, is beneficial to oocytes by increasing MII formation and their cleavage rate after parthenogenetic activation. Therefore, in the following experiments prematured oocytes were always treated with BDNF during IVPM but not during IVM and compared with oocytes exclusively subjected to IVM (not to IVPM) in the absence of BDNF.
Due to the experimental design, statistical analysis compared IVM versus IVM+BDNF or IVPM versus IVPM+BDNF; other comparisons than these cannot be done. Values are representative of at least five experimental replicates.
Values with different superscript letters within column denote a significant difference between groups (p<0.05).
Rates of first polar body (PB), cleavage and blastocyst were estimated in relation to the number of activated oocytes cultured. The rate of hatched blastocyst was estimated in relation to the number of blastocysts at day 7.
Due to the experimental design, statistical analysis compared IVM versus IVM+BDNF or IVPM versus IVPM+BDNF; other comparisons than these cannot be done. Values are representative of at least five experimental replicates.
No significant difference was found between groups within column (p>0.05). The percentage of blastocyst TUNEL-positive cells was estimated in relation to the total number of blastocyst cells.
Prematuration increases expression of POU5F1 and IFNT2 in cloned blastocysts
To investigate the effect of prematuration on oocyte quality regarding production of cloned embryos, oocytes were subjected to IVPM followed by IVM. A significant (p=0.006) increase in the first PB extrusion rate was observed (Table 4) in prematured oocytes (693/1000, 69.3%) compared to control matured oocytes (639/1006, 63.5%). In addition, the fusion rate (Table 4) was also increased (p=0.02) by prematuration (397/501, 79.2%) compared to oocytes not subjected to IVPM (345/473, 72.9%). However, embryonic development as assessed by cleavage (p=0.68) and blastocyst (p=0.84 and 0.8, respectively for days 7 and 8) rates was not affected by prematuration (Table 4). Also, the number of nuclei (p=0.59) and TUNEL-positive nuclei (p=0.63) in blastocyst did not differ between the treated and the control groups (Table 5), indicating oocyte prematuration does neither alter the development nor the quality of cloned embryos.
Values with different superscript letters within column denote a significant difference between groups (p<0.05).
The percentage of cleavage and blastocyst were estimated in relation to the number of fused oocytes. Values are representative of six experimental replicates.
No significant difference was found between groups within column (p>0.05). The percentage of blastocyst TUNEL-positive cells was estimated in relation to the total number of blastocyst nuclei. Values are representative of three experimental replicates.
To further investigate the effect of prematuration on cloned and parthenogenetic development we evaluated the expression levels of genes related to the quality of the inner cell mass and trophoblast of blastocyst, that is, POU5F1 and IFNT2, respectively (Fig. 1). For POU5F1, a 1.5-fold increase in expression levels was found in the treated blastocysts compared to the control both in cloned (p=0.02) and parthenogenetic (p=0.0002) embryos. Similarly, cloned (p=0.02) and parthenogenetic (p<0.0001) blastocysts produced with prematured oocytes (treated group) showed a two- and a 2.5-fold increase in IFNT2 expression compared to the control group. In summary, these results reveal a positive effect of prematuration on blastocyst quality.
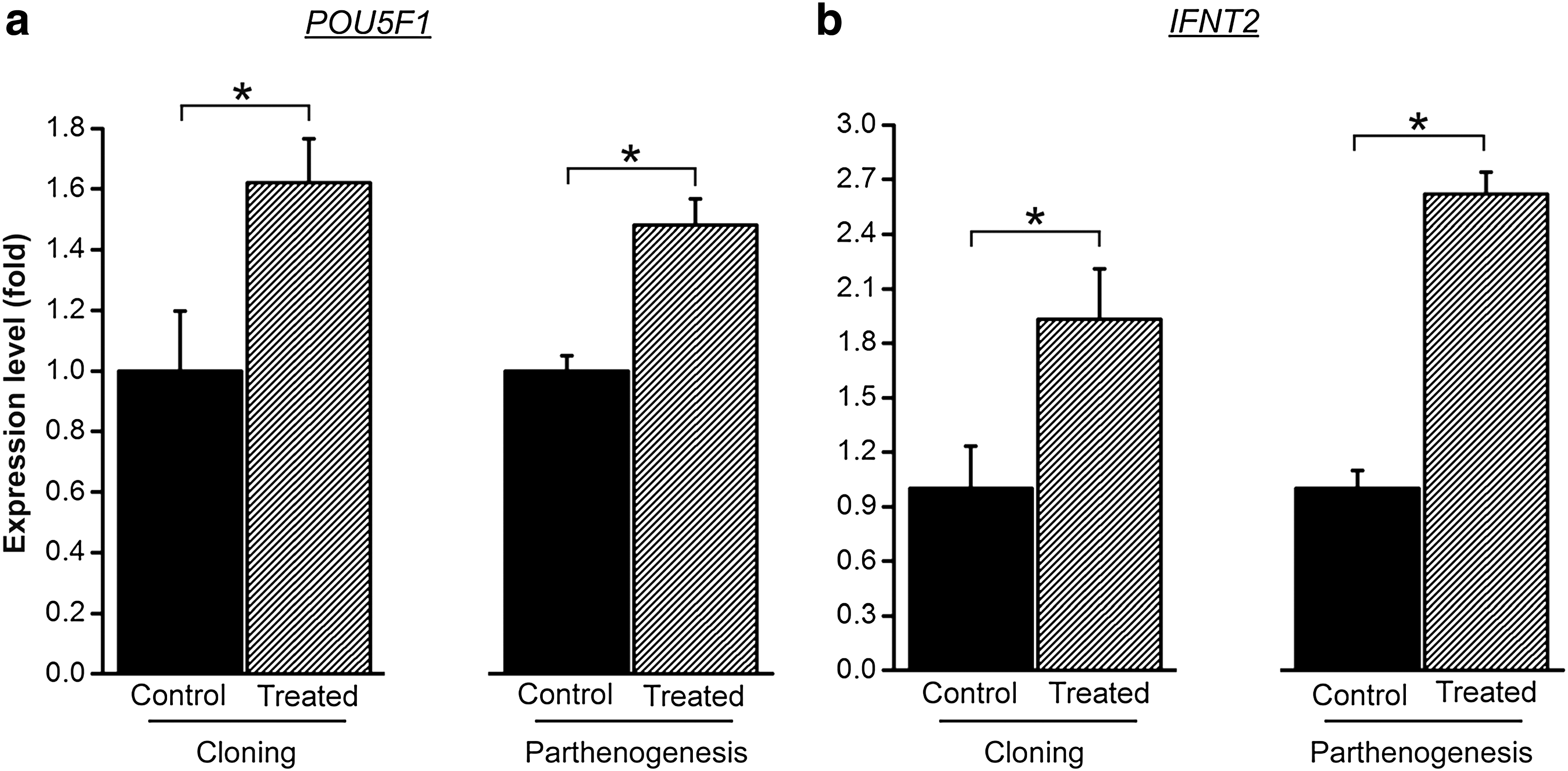
Blastocyst expression of POU5F1 and IFNT2 is increased by prematuration. The amount of POU5F1 (
Oocyte mtDNA pool is diminished by prematuration in immature oocytes but not in mature oocytes and cloned blastocysts
By increasing the time oocytes are cultured in vitro before resuming meiotic maturation, IVPM oocytes would have a window of opportunity to continue replicating and therefore, increase their mtDNA amounts. Thus, we aimed at investigating whether prematuration affects oocyte and blastocyst mtDNA copy number. First, IVPM oocytes (not subjected to IVM), presumably at the GV stage, were compared with oocytes at the GV stage (immature oocytes collected immediately after follicle aspiration). A significant difference (p=0.01) was observed between these two groups (Fig. 2). Immature oocytes that were prematured (treated) contained 595,579±31,922 (mean±SEM; ranging from 205,389 to 968,269) mtDNA copies, whereas GV oocytes that were not prematured (control) contained 718,585±34,775 (ranging from 425,142 to 1,203,532). After IVM, the amount of mtDNA in the oocytes from the control group dropped (p<0.05) to 522,179±45,617 (mean±SEM; ranging from 42,065 to 1,184,177) copies, whereas the 498,771±33,231 (ranging from 228,184 to 926,332) copies found in mature oocytes from the treated group remained the same (p>0.05) compared to immature oocytes from the treated group (Fig. 2). Therefore, mtDNA copy number did not differ (p=0.68) between both treated and control groups after IVM (Fig. 2). Similarly, the 765,332±51,104 (mean±SEM; ranging from 466,760 to 1,549,368) copies found in cloned blastocysts from the treated group was not different (p=0.42) than the 816,627±40,235 (ranging from 378,163 to 1,212,006) copies present in cloned blastocysts from the control group (Fig. 2). In summary, although mtDNA was diminished by prematuration in immature oocytes, these results suggest that the treatment does not affect mtDNA copy number in mature oocytes and cloned blastocysts.
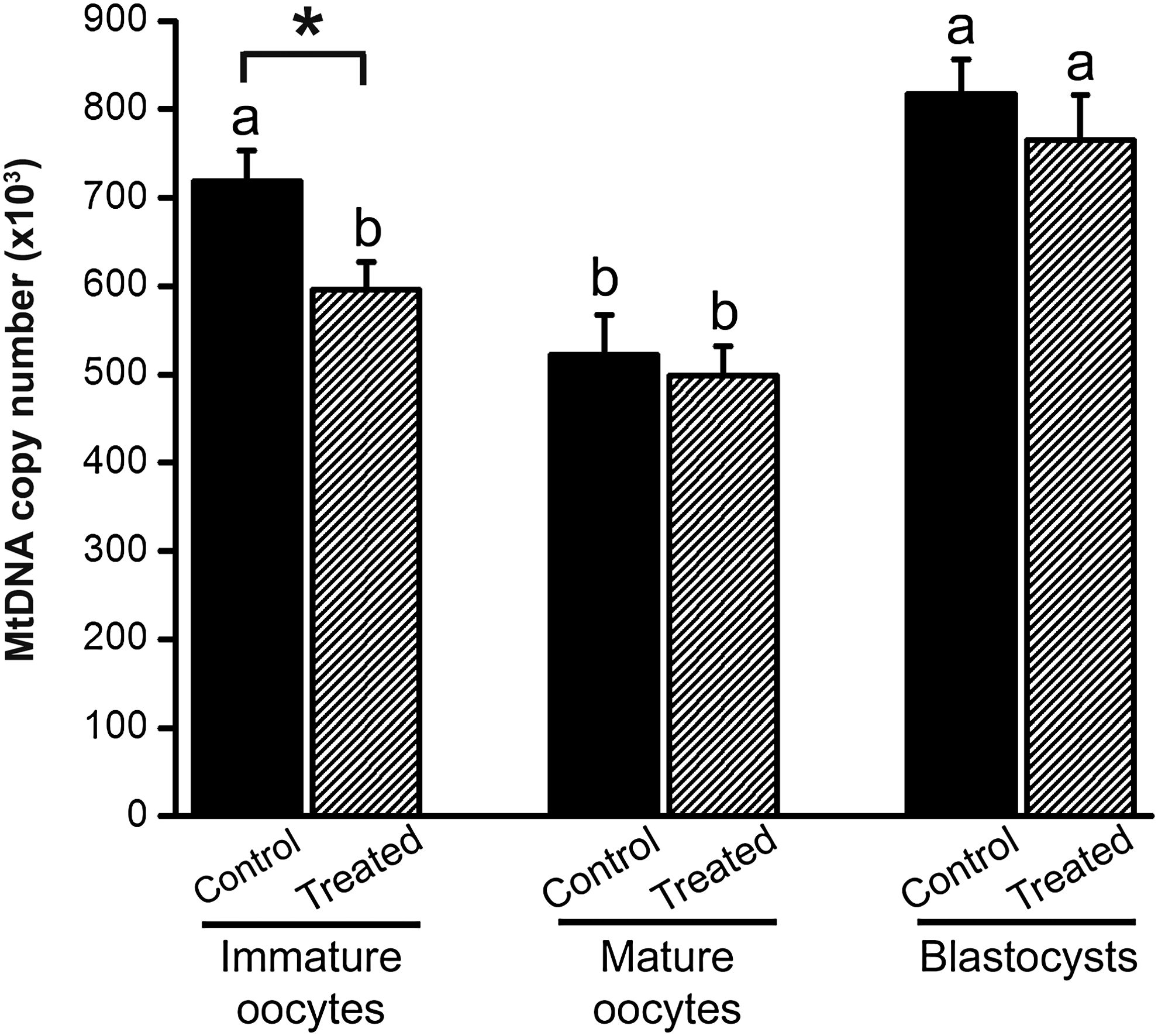
Oocyte mtDNA pool is diminished by prematuration in immature oocytes but not in mature oocytes and cloned blastocysts. The amount of mtDNA is expressed as copy number per oocyte or blastocyst regarding IVM (control) and IVPM (treated) treatments and three stages (immature oocyte, mature oocyte, and blastocyst). The (*) denotes difference between IVM and IVPM (p<0.05). Different letters over bars denote a significant difference among days within treatment (p<0.05). Values are representative of three experimental replicates and are reported as mean±SEM.
Prematuration enables development to term of viable clones
To definitely test the effect of prematuration on cloned embryos, 26 and 28 blastocysts from the treated and the control groups, respectively, were transferred to recipient cows to assess pregnancy rates. No significant difference in pregnancy rates were found between the treated and the control groups at day 35 (p=0.24) and day 60 (p=0.92) of development (Table 6). Moreover, development to term of these pregnancies resulted in the live birth of two treated calves and one control calf (p=0.51), all three weighing within the newborn breed average for the Nellore breed (Table 6). These findings indicate that the prematuration of bovine oocytes does not affect oocyte competence and is a viable alternative for SCNT procedures using standard IVM protocols.
No significant difference was found between groups within column (p>0.05). Pregnancy rates at day 35 and day 60, and birth rate were estimated in relation to embryos transferred to recipient cows. Values are representative of three experimental replicates.
Discussion
One of the limiting components of SCNT is that approximately 400 immature oocytes are required for each healthy cloned calf born (Meirelles et al., 2010). However, even when a larger number of oocytes can be obtained from slaughterhouse ovaries, an experienced operator can only micromanipulate a limited number of oocytes within a day due to the short life span of oocytes and adverse consequence of aging on developmental competence. In this sense, protocols for inducing a temporary arrest of meiotic maturation, that is, IVPM, allows more flexibility for the timing of manipulations, and thus, enables the use of the oocytes from a single batch of ovaries in 2 consecutive days (Galli et al., 2003a; Lagutina et al., 2002). Moreover, it is believed that IVPM provides the GV-arrested oocyte with additional time to further acquire developmental competence prior to IVM (Ferreira et al., 2009b; Hashimoto et al., 2002; Hendriksen et al., 2000). In the present study we show that delaying meiotic resumption for 24 h prior to IVM results in oocytes capable of supporting development of cloned calves as efficiently as nonarrested oocytes. Thus, when the number of oocytes collected from a batch of ovaries is superior to the amount that can be handled in a day, IVPM can double the number of clones produced by a single operator with a single batch of ovaries.
Apart from its role in the nervous system, BDNF is secreted by cumulus cells and is present in the follicular fluid (Seifer et al., 2003), which may explain its ability to improve oocyte competence when added to prematuration and maturation media (Adona et al., 2011; Kawamura et al., 2005). In the present study, BDNF was added to prematuration medium at 10 ng/mL as previously suggested by Adona et al. (2011). Although we did not observe a positive effect of BDNF during IVM, oocytes prematured in its presence showed higher rates of MII formation and cleavage. No other significant effect was seen on embryonic development when IVPM was performed with BDNF. On the other hand, oocytes exposed to BDNF during IVM showed lower pronuclear development after parthenogenetic activation. Using an IVF model, Adona et al. (2011) found that BDNF supplementation during IVPM had no effect on oocyte gene expression or embryonic development. Although the use of BDNF during prematuration did not lead to better rates of embryo production in our study, MII formation and cleavage rates were superior to control, indicating an improvement of oocyte developmental competence. However, it is worthwhile to mention that rather than improving the ability of the oocyte to complete subsequent nuclear maturation, the higher rates of MII formation might reflect an acceleration of nuclear maturation. Moreover, because BDNF during IVM led to lower pro-nuclear rates, we decided to use BDNF exclusively during prematuration in the subsequent experiments.
Our second aim was to verify the benefits of prematured oocytes as recipient ooplasts for producing cloned blastocyst by SCNT. Previous studies have shown that prematuration can successfully be used to arrest meiotic division without negative consequences to oocyte viability (Adona and Leal, 2004; Adona et al., 2008a, 2008b; Coy et al., 2005a; Galli et al., 2003a; Hashimoto et al., 2002; Kubelka et al., 2000; Lagutina et al., 2002; Ponderato et al., 2002). In this context, development to term of pigs produced by IVF has been reported using prematured oocytes (Coy et al., 2005b). However, due to the poor efficiency of SCNT to produce viable offspring (Meirelles et al., 2010), it seems logical to examine the advantages of prematured oocytes to produce cloned offspring. Our data provide evidence that the use of prematured oocytes for producing cloned blastocyst by SCNT has at least the same efficiency as control oocytes not subjected to prematuration. Although we have found an increase in first PB extrusion and fusion rates using prematured oocytes compared to controls, no effect was observed on developmental rates (cleavage and blastocyst). Moreover, embryo quality as judged by the number of nuclei and fragmented nuclei (TUNEL positive) in blastocysts was not affected by the treatment. Nonetheless, in disagreement with these findings, analysis of gene expression suggested a positive effect of prematuration on quality of cloned blastocysts. Both the POU5F1 (previously known as OCT4) and the IFNT2 (interferon tau) are related to embryo competence to develop into a viable offspring. The POU5F1 is a transcription factor known for its role in embryonic development, especially during early embryogenesis, and it is necessary for embryonic stem cell pluripotency. On the other hand, IFNT2 plays a major role in maternal recognition of pregnancy in cattle. Transcript levels of these two genes were upregulated by prematuration in cloned and parthenogenetic blastocysts, suggesting that blastocysts produced using prematured oocytes are more competent in establishing their germ cell lineage and in interacting with the maternal environment. Yet, although an increase in the transcription levels of a specific gene usually results in increased levels of the protein encoded by the same gene, it remains to be proven that POU5F1 and IFNT2 protein levels were augmented in cloned blastocysts produced with prematurated oocytes. Regarding the low efficiency of SCNT, any improvement of the technique that allows an increase in viable offspring is welcome (Meirelles et al., 2010).
To further assess the effect of oocyte competence, our next goal was to determine whether prematuration affects the content of mtDNA (May-Panloup et al., 2007). During folliculogenesis, mtDNA copy number increases sharply in oocytes due to continuous replication, leading to fully grown oocytes with hundreds of thousands of copies (May-Panloup et al., 2007; Poulton et al., 2010). Because oocytes are often recovered from follicles before they have reached their final size, we hypothesized that prematuration would provide an extra time for the oocyte to continue its growth, and therefore, increase mtDNA copy number. In contrast to our hypothesis, our data indicate that the mtDNA content is diminished during prematuration, resulting in immature oocytes with lower levels of mtDNA than the levels found in immature oocytes collected immediately after removal from follicles. Because in our experiment mtDNA copy number decreased during IVM in control oocytes (not subjected to prematuration), we think the mtDNA content decreased during prematuration not due to the treatment itself but due to oocyte removal from follicle. Corroborating with this hypothesis, mtDNA copy number was unaltered in mature oocytes between IVPM and IVM, suggesting the mtDNA content reaches after prematuration the same levels as mature oocytes not prematured. Moreover, the mtDNA content was not different between IVM and IVPM in cloned blastocysts. Although there is no report showing a similar decrease of mtDNA content during oocyte maturation, mtDNA copy number was shown to diminish during the earliest stages of development (i.e., until early morulae) in cattle (May-Panloup et al., 2005; Smith et al., 2005). An active destruction of mtDNA molecules occurring during IVM might explain the lower levels of mtDNA found in immature oocytes after prematuration if butyrolactone I could not prevent anticipation of mtDNA destruction. On the other hand, although we have found the mtDNA content is unchanged by prematuration in mature oocytes, mtDNA depletion would not necessary imply in poorer oocyte competence. Previous studies indicate that mtDNA copy number in oocytes does not correlate to its competence to develop into a viable blastocyst in cattle (Chiaratti and Meirelles, 2010). Indeed, a recent work of our group has shown that mtDNA replication is modulated during embryogenesis in order to compensate for mtDNA depletions (Chiaratti et al., 2010). Thus, in the case of oocyte mtDNA depletion, it is expected that the mtDNA content be replenished until the blastocyst stage in cattle, preventing thus an effect on developmental competence.
Finally, we decided to investigate whether the beneficial effect of prematuration on blastocyst quality (i.e., POU5F1 and IFNT2 expression) would induce higher pregnancy and birth rates of clones. With this aim, blastocysts produced by SCNT using oocytes subjected or not to prematuration were transferred to synchronized recipient cows and their pregnancies were monitored until term. No significant difference was found between the IVPM and the control group regarding pregnancy evaluation on days 35 and 60, or birth rate. However, two viable calves were born using prematured oocytes, which indicates that oocytes can be arrested at the GV for at least 24 h previous to SCNT without negative consequences to calves produced. Although we cannot confirm this statement, it is possible that a positive effect of prematuration on blastocyst quality would only become apparent with a larger number of embryo transfers. However, due to restriction in the number of recipient available, we were only able to perform about 30 embryo transfers per group, which was insufficient to show significantly that prematuration increases birth rate.
These findings indicate that oocytes arrested at the GV stage of meiosis for 24 h can be used as recipient ooplasts for SCNT without negative consequences to term development, providing an interesting option to SCNT. Prematuration of oocytes that exceed the number necessary for a SCNT routine (about 250 immature oocytes) becomes an interesting option for using surplus oocytes, eliminating the need for another oocyte collection the following day. In our experience, over 400 viable oocytes are routinely retrieved at each aspiration routine from bovine ovaries collected at the abattoir. Prematuration of half of the oocyte pool (which in other circumstances would have been discarded) before being subjected to IVM and SCNT doubles the number of cloned blastocysts and offspring produced. Moreover, prematuration provides extra flexibility to the process of embryo production allowing adjustments to the laboratory routine. Finally, because prematuration represents a brief period before IVM to increase oocyte competence, development of better prematuration media that increases even further their development competence would have a direct implication on the efficiency of SCNT.
In contrast to other drugs that are broad-spectrum inhibitors, butyrolactone I is a selective inhibitor of MPF activity through CDK1 inhibition. Moreover, butyrolactone I is among the drugs mostly used to arrest oocyte in GV (Adona et al., 2008a, 2008b; Ferreira et al., 2009a). Nonetheless, although butyrolactone I specifically inhibits MPF activity, in oocytes it has been shown to also indirectly inhibit MAPK activity (Kubelka et al., 2000) and to affect transcription and translation (Tomek et al., 2002a. 2002b; Vigneron et al., 2004). On the other hand, the use of butyrolactone I to block meiotic progression has resulted in developmental rates that were not different from nontreated controls, both in vitro (Adona et al., 2008b; Lagutina et al., 2002; Lonergan et al., 2000; Mermillod et al., 2000; Ponderato et al., 2001) and in vivo (Coy et al. 2005a, 2005b; Ponderato et al., 2002). Therefore, although we cannot exclude the possibility that butyrolactone I could have unknown side effects, our results and those of others did not cause a decrease in developmental rates.
In conclusion, the present study provides clear evidence that prematured oocytes can be used to produce viable calves by SCNT. Moreover, we show a positive effect of prematuration on embryo quality as judged by expression of POU5F1 and IFNT2 genes, but this positive effect was not observed during postimplantation development, resulting in no augmentation of birth rate. MtDNA copy number was disturbed in immature oocytes by prematuration, but this alteration did not reflect in mature oocytes and cloned blastocysts. To our knowledge, this is the first report on the birth of cloned animals using prematured oocytes. These findings have significant implications for laboratories of in vitro embryo production, mainly those working on SCNT, where prematuration can be used to increase the flexibility of the procedure. Prematuration has also the advantage of allowing the use of surplus oocytes in a different SCNT routine, which could double the number of cloned offspring.
Footnotes
Acknowledgments
The authors appreciate the assistance provided by Clonest Company, Pirassununga, SP, Brazil. This research was supported by research grants (06/58536-0 and 05/59694-5) from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), SP, Brazil.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.