
Abstract
Abstract
Demecolcine-assisted/induced enucleation has been used in nuclear transfer cloning procedures for many species, yet its mechanism of action remains unclear. Primarily because oocytoplasm protrusion induced by demecolcine is inhibited by the presence of cytochalasin, its use has had limited application. In this experiment, we investigated the microtubule and microfilament alterations in bovine oocytes after demecolcine and/or cytochalasin B (CB) treatments by immunocytochemical staining. We also examined mechanical enucleation of demecolcine-treated oocytes in cytochalasin-free medium. The results showed that demecolcine-treatment disrupts the balance between microtubule/microfilament interactions primarily by deleting microtubules and with little effect on the microfilaments that we believe accounts for the membrane protrusion. The CB treatment reduced the amount of microfilaments but had little effect on the microtubules. Most demecolcine-induced membrane protrusions disappeared when exposed to CB. Western blotting showed that CB treatment increases G-actin, which indicates a decrease in the microfilaments. High oocyte enucleation, survival, and developmental rates occurred when demecolcine-assisted enucleation was carried out in a CB-free solution. Higher blastocyst development rates and blastocyst cell numbers were achieved compared to control, indicating that CB is not necessary in the enucleation procedure of bovine oocytes. This study provides a clearer understanding of the mechanism for the demecolcine-induced oocyte membrane protrusion, and substantiates the practical use of demecolcine-assisted enucleation in a CB-free environment.
Introduction
Demecolcine treatment of MII oocytes to induce membrane protrusions has been reported in pigs (Li et al., 2006; Yin et al., 2002), cattle (Li et al., 2009; Russell et al., 2005; Saraiva et al., 2009; Tani et al., 2006; Vajta et al., 2005), mice (Costa-Borges et al., 2009; Gasparrini et al., 2003; Ibanez et al., 2003), sheep (Hou et al., 2006), goats (Lan et al., 2008), and hamsters (Wang et al., 2009). Once a protrusion has occurred, microtubule repolymerization will resume whenever demecolcine-treated oocytes are cultured in demecolcine-free medium (Costa-Borges et al., 2009; Saraiva et al., 2009). Although demecolcine-assisted enucleation has been widely used, the mechanism of its action is little understood. A recent study using goat oocytes showed that most demecolcine-induced protrusions will abate within 30 min of treatment when used with cytochalasin B (Lan et al., 2008). Cytochalasin B is widely used for mechanical enucleation of oocytes. The use of demecolcine with Cytochalasin B is self-limiting because it interferes with its mode of action.
Actin is one of the major proteins in mammalian oocytes. Most developmental events are dependent on the normal distribution of filamentous F-actin. Nonfilamentous or global G-actin is regularly polymerized into F-actin; the latter constitutes the microfilament, which is a major component of the cytoskeleton. Early studies have shown that cytochalasins can inhibit the polymerization of G-actin and induce rapid depolymerization of filamentous actin by binding to high-affinity sites associated with F-actin (Flanagan and Lin. 1980). Although cytochalasins have been widely used in the micromanipulation of mammalian oocytes, other methods such as demecolcine may be better suited particularly for the enucleation of farm animal oocytes. Demecolcine has had limited use, however, because cytochalasins counter effect the induced membrane protrusions. This limitation could potentially be dealt with if the enucleation procedure was performed in a cytochalasin-free medium.
In this experiment, we investigate the dynamic relationship of the microtubules and microfilaments that constitute the major components of the oocyte cytoskeleton by comparing the effects of demecolcine and cytochalasin B (CB). We also investigate the efficacy of using demecolcine alone for enucleation.
Materials and Methods
Chemicals
Unless otherwise noted, all chemicals were purchased from Sigma Chemical Co. (St. Louis, MO). Each experiment included at least three replicates.
Oocyte maturation in vitro (IVM)
Maturation of bovine oocytes was as described previously (Li et al., 2004). Briefly, bovine cumulus oocyte complexes (COC) were aspirated from 3–8-mm diameter follicles on ovaries obtained from a local slaughterhouse. Only COC with compact cumulus layers and a homogenous ooplasm were selected. The COC were matured in TCM 199 with Earle's salts, L-glutamine, and sodium bicarbonate (Gibco Inc., Grand Island, NY) supplemented with 10% FBS (HyClone, Logan, UT), 25 μg/mL gentamycin, 0.01 U/mL FSH (NIH-FSH-S17), 0.01 U/mL LH (USDA-bLH-6) in four-well plates with 0.6 mL medium and 30–50 oocytes per well at 38.5°C in humidified atmosphere of 5% CO2 and air.
Donor cell culture and preparation
The donor fibroblast cell line (Holzer 064) for nuclear transfer (NT) was established from an ear biopsy and is maintained as a resource cell line in our lab (Aston et al., 2006). The donor cells were cultured in DMEM/F12 (1:1) supplemented with 10% fetal bovine serum (FBS) and antibiotics at 38.5°C in a humidified atmosphere of 5% CO2 and air. Passage 6–12 cells at 80–100% confluence were cultured in DMEM/F12 (1:1) supplemented with 0.5% FBS for 20–24 h and used for NT. Before nuclear transfer, the cells were dissociated with 0.25% trypsin and resuspended in HEPES-buffered SOF (HSOF) medium (Gandhi et al., 2000). Only small (10–12 μm), round cells were used as donor cells.
Treatments of recipient oocytes with demecolcine and CB
Oocytes matured for 20–21 h were used in this study. Cumulus cells were removed by pipetting COC in HSOF medium containing 0.1% hyaluronidase. Only oocytes with first polar bodies (PB1) were used in the experiments. In preliminary experiments, 70–80% of in vitro matured oocytes exhibited a PB1under the culture conditions described above. The oocytes were incubated in TCM-199 supplemented with 0.6 μg/mL demecolcine for 30–40 min. The demecolcine concentrations selected for this study have been verified to induce membrane protrusions containing a maternal chromosome mass efficiently (∼90%) (Li et al., 2004; Tani et al., 2006; Vajta et al., 2005). To determine the effect of CB treatment on the demecolcine-induced membrane protrusions, demecolcine-treated oocytes were incubated in HEPES-buffered SOF supplemented with 0.6 μg/mL demecolcine and 7.5, 3.25, 2.5, 1, 0.5, or 0 μg/mL CB at room temperature for 30 min. The status of the membrane protrusion was observed at 0, 10, 20, and 30 min postincubation in CB solution.
Enucleation
To determine the effects of demecolcine or CB treatment on the enucleation of bovine oocytes, oocyte enucleation was carried out as follows: (1) enucleation was performed by traditional mechanical enucleation with the CB method, in which the PB1 and surrounding cytoplasm were removed with a piezo-actuated micropipette in HSOF supplemented with 7.5 μg/mL CB (Dominko et al., 2000; Keefer et al., 1994). About 20% of the cytoplasm is removed with this method (CB-ENU). (2) The enucleation procedure was performed by mechanical enucleation as described in method 1, but it was carried out in HSOF without CB or demecolcine (D/CB-free-ENU). (3) Enucleation was performed by a new chemically assisted enucleation without the CB method. Briefly, oocytes were pretreated with 0.6 μg/mL demecolcine for 30–40 min as describe above. Enucleation was performed by aspirating the cytoplasmic protrusions induced by demecolcine treatment in HSOF without CB (DT-ENU). A small amount of cytoplasm (<1%) is removed with this method (Fig. 1). In all groups, the micromanipulation was carried out in HSOF containing 10 μg/mL Hoechst 33342. The aspirated cytoplasts were stained with Hoechst 33342 and assessed by UV illumination to determine enucleation accuracy. The enucleation accuracy and oocyte survival rates were recorded.

Enucleation of demecolcine-treated oocyte in cytochalasin B-free solution. Democolcine treatment induced a cytoplasmic protrusion (arrow-pointed) on the oocyte membrane (
Immunocytochemical staining of oocytes
Oocytes were fixed and extracted with 2% paraformaldehyde in PBS containing 0.5% Triton X-100, 1 mM dithiothretol and 0.3% BSA overnight at 4°C. After washing twice with PBS containing 0.01% Triton X-100, oocytes were blocked in PBS containing 150 mM glycine, 0.02% Sodium azide, 0.1% Triton X-100 and 2% bovine serum albumin (BSA) for 1 h at 37°C. The oocytes were then incubated in a FITC-conjugated mouse anti-α-tubulin (1:100) and Texas Red-X phalloidin (Molecular Probes, Eugene, OR; 1:200) in PBS for 2 h at 37°C. DNA was stained with 10 μg/mL of Hoechst 33342. Oocytes were mounted on slides in 50% glycerol in PBS and then examined under a Zeiss epifluorescent microscope (Carl Zeiss Optical, Inc., Chester, VA). Images were captured by digital camera with Axio Vision LE Program (Carl Zeiss, Thornwood, NY). Four groups of oocytes were used for immunocytochemical (ICC) staining. In group 1, oocytes were treated with 0.6 μg/mL demecolcine for 30–40 min as described above, and fixed for ICC staining (DT group). In group 2, oocytes were incubated in HSOF supplemented with 7.5 μg/mL CB at room temperature for 30 min, and then fixed for ICC staining (CT group). In group 3, oocytes were first treated with demecolcine, then incubated in CB medium, and fixed for ICC staining (DT + CT group). In group 4, untreated oocytes were fixed and used as control (control group).
Nuclear transfer
To investigate the in vitro development of cloned embryos reconstituted with cytoplasts produced by the demecolcine treatment minus CB, two groups of nuclear transfer experiments were carried out. In group 1, the demecolcine-treated oocytes were enucleated and PB1s were removed in HSOF supplemented with demecolcine without CB as described above. In group 2, oocytes were enucleated by the conventional method utilizing CB. PB1s and surrounding cytoplasm were removed in HSOF supplemented with 7.5 μg/mL CB. Prior to cell insertion, the resulting cytoplasts were allowed to recover from enucleation in SOFaa medium for 1.5 h. A cell was inserted into the perivitelline space of an enucleated oocyte and the cytoplast–cell constructs were induced to fuse by a single 22 sec 2.8 kV/cm DC pulse in electrofusion medium (0.28 M sorbitol in water, supplemented with 0.5 mM HEPES, 0.1 mM Ca(CH3COO)2, 0.5 mM Mg(CH3COO)2 and 1 mg/mL bovine serum albumin). The fused embryos were activated with 5 μM ionomycin for 5 min, followed by treatment with 1 mM 6-dimethylaminopurine (6-DMAP) and 5 μg/mL cycloheximide (CHX) in SOFaa for 4.5 h at 38.5°C in 5% CO2 in air.
In vitro culture of nuclear transfer (NT) embryos
After activation, NT embryos were cultured under mineral oil in 50 μL droplets of SOFaa + 5% FBS with a monolayer of bovine cumulus cells at 38.5°C in 5% CO2 and air for 8 days. The medium was changed at days 3, 5, and 7 from culture. The cleavage and blastocyst development were evaluated on days 3 and 8 after activation (day 0). Blastocyst cell number was determined by fluorescent microscopy after staining with Hoechst 33342.
G-actin level analysis by Western blot
To examine the G-actin level of demecolcine or cytochalasin-treated oocytes, equal numbers of untreated, demecolcine, or cytochalasin-treated oocytes were lysed and extracted and the resulting protein was transferred to a nitrocellulose membrane by using commercial kits according to their prescribed protocols. Briefly, the oocytes were lysed and extracted with cell lysis buffer (Fermentas, Hanover, MD). The extract was then frozen at −80°C until use. The total proteins were separated by electrophoresis with a 10% separating gel (Pierce Precise Protein Gels) for 45 min at 110 V and 115 mA. After electrophoresis, the gel was washed with transfer buffer for 3 min and then the total protein was transferred electrophoretically onto a nitrocellulose membrane for 45 min at 20 V (Biorad, Hercules, CA; Trans-Blot SD Semi-Dry electrophoretic transfer cell). The β-actin level of samples was then analyzed with a WesternBreeze® Chromogenic Kit (Anti-Mouse, Invitrogen, Carlsbad, CA). The nitrocellulose membrane was blocked and incubated in the first antibody (mouse anti-β-actin, 1:1000, Sigma) solution and then processed according to the instructions of the kit. The Western blot images were scanned into a computer and analyzed with AxioVision software (Zeiss).
Statistical analysis
Differences in enucleation survival, enucleation accuracy rates, cell numbers, blastocyst rates, and β-actin levels between groups were analyzed by one-way ANOVA with SPSS software. Values of p<0.05 were considered statistically significant.
Results
Effect of CB treatment on the duration of demecolcine-induced membrane protrusion
Demecolcine treatment induced membrane protrusions on bovine oocytes with high efficiency (86.7–100%). Membrane protrusions gradually became indistinguishable when oocytes were exposed to CB. In the 7.5 μg/mL CB solution, 96.7% of the demecolcine-induced membrane protrusions disappeared within 30 min. The percentage of membrane protrusions decreased from 96.8 to 16.1% within 30 min in 0.5 μg/mL CB solution. In CB-free medium, 88.9% of oocytes had recognizable membrane protrusions at 30 min postincubation (Fig. 2). These results indicate that CB will counter affect the benefits of demecolcine-assisted enucleation.
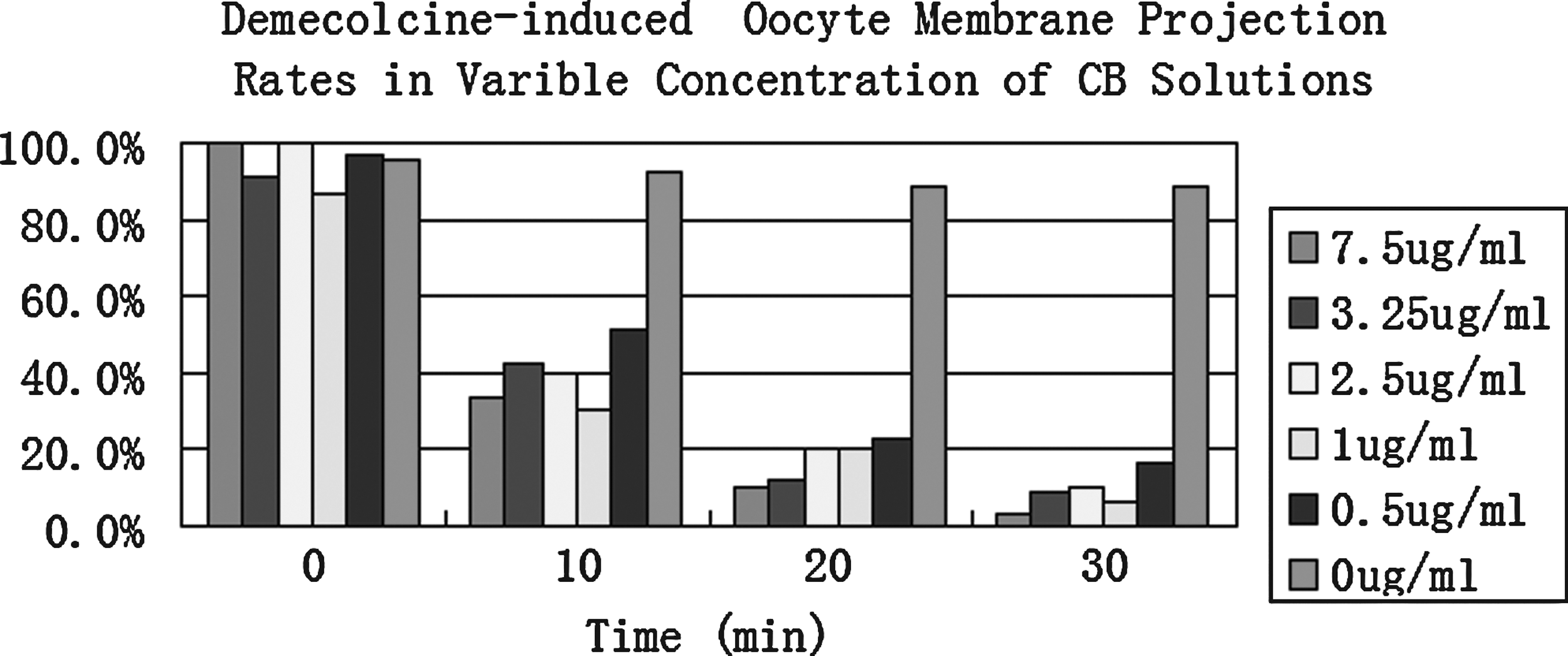
Demecolcine-induced membrane projection rates in variable concentrations of CB. Demecolcine treatment (0.6 μg/mL, 30 min)-induced membrane protrusion rates of 86.7–100%. These protrusion rates decreased to 3.3–16.1% within 30 min in 0.5–7.5 μg/mL CB solutions. In the CB-free medium, 88.9% of oocytes still had recognizable membrane protrusions at 30 min postincubation (n=27–33 in each treatment).
Effects of cytochalasin and demecolcine on bovine oocyte enucleation
To determine cytochalasin treatment on the survival of bovine oocyte enucleation and to evaluate our new demecolcine-assisted CB-free enucleation method, enucleation was carried out with CB-ENU, (D/CB-free-ENU) and DT-ENU) as described above. High survival rates (98.2 or 96.3%) were achieved when enucleation was carried out in CB-free medium with (DT-ENU) or without (D/CB-free-ENU) demecolcine, which was comparable to the result obtained by CB-ENU (100%). Enucleation accuracy was higher in the DT-ENU group than in either the CB-ENU or D/CB-free-ENU group (100% vs. 77.8%, p<0.001; 100% vs. 70.1%, p<0.001) (Fig. 3). These results demonstrate that cytochalasin is not necessary for mechanically or chemically assisted enucleation of bovine oocytes and the demecolcine-assisted CB-free enucleation is very efficient for bovine oocytes.
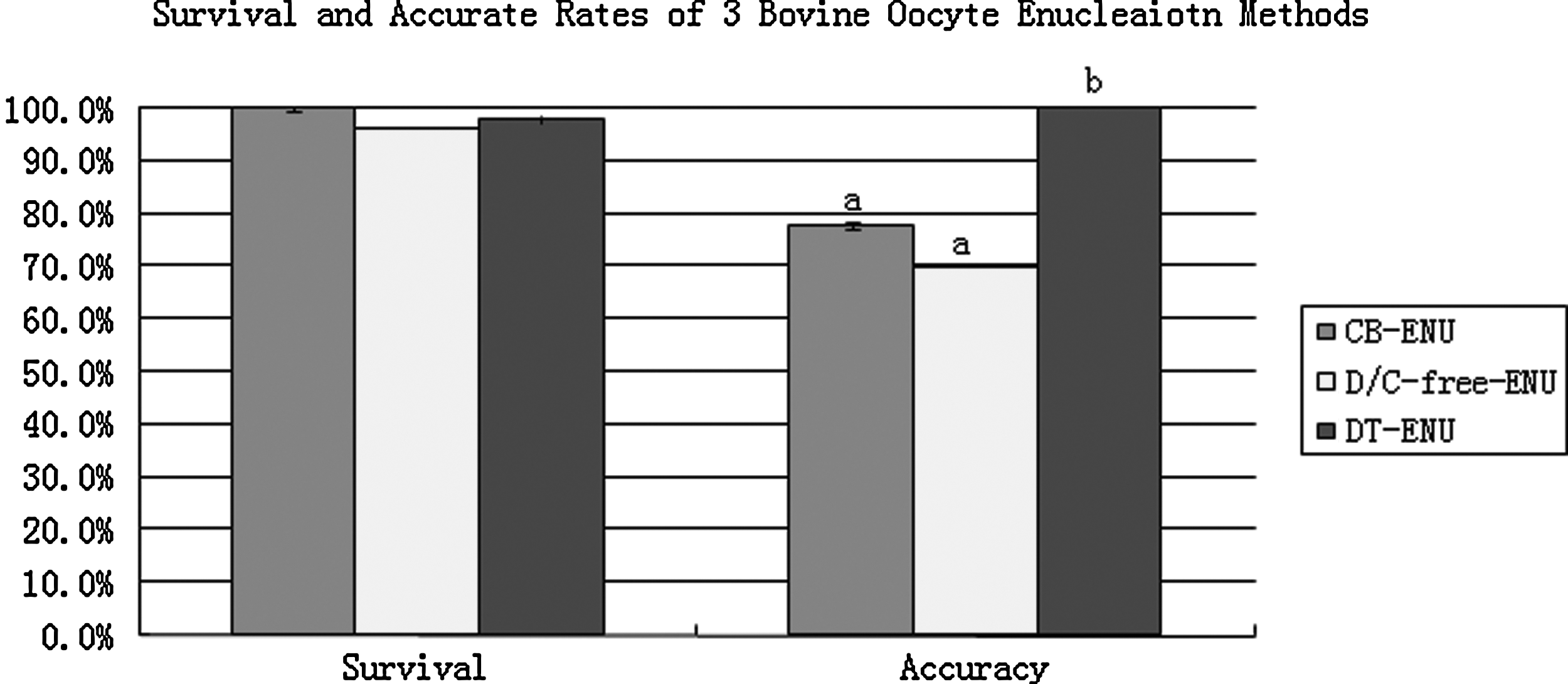
Survival and accuracy rates of CB-ENU (n=63), D/CB-free-ENU (n=80), and DT-ENU (n=55), enucleation methods. Detailed description of the above enucleation methods are in the Materials and Methods section.
Cytoskeleton changes of demecolcine- and cytochalasin-treated bovine oocytes
The cytoskeleton status in the demecolcine- or CB-treated oocytes provided the probable mechanism for the demecolcine-induced cytoplasmic protrusion. In the untreated control MII oocytes, the microtubules were concentrated to form the normal spindle structure with microfilaments distributed uniformly in the cytoplasm (Fig 4a). In the demecolcine-treated oocytes, the spindle microtubules were nearly depleted by demecolcine, but the microfilaments were hardly affected (Fig. 4b). The demecolcine treatment seemed to have altered the balance between the interaction of microtubules and microfilaments in the treated oocytes, which appears to be the result of the cytoplasmic protrusion. Without the aid of the microtubules, the chromosomes were compressed by the microfilaments to form a protrusion on the oocyte membrane. We believe this hypothesis can be substantiated by the results observed in CB-treated and demecolcine + CB-treated oocytes. In the CB-treated oocytes, the filamentous actin (microfilament) was partially depolymerized, although the spindle structure was not seriously affected (Fig. 4c). In the oocyte group that was first treated by demecolcine and then by CB, microtubules were deleted and microfilaments were partially depolymerized (Fig. 4d). In this situation, the protrusion induced by demecolcine treatment would withdraw from the oocyte membrane as the microfilaments depolymerized by CB treatment and the protrusion could no longer be maintained.

Cytoskeletal status of oocytes treated with demecolcine and cytochalasin B (CB). The shape of the cytoskeleton was well maintained by its microtubules and filaments in the untreated control oocyte (
Cytoplasts produced with demecolcine, without CB enucleation supported better in vitro development of cloned embryos
To examine in vitro developmental capacity of cloned embryos reconstituted with the demecolcine-assisted, CB-free enucleation method, blastocyst development rate and blastocyst cell number were compared between this newly proposed enucleation method and the conventional blind aspirating method (control). The blastocyst rate and average cell numbers were significantly higher in the demecolcine-assisted, CB-free method than in the controls (Table 1).
Enucleation was performed by aspirating PB1 and surrounding cytoplasm in CB solution (CB-enucleation).
Enucleation was performed by aspirating the demecolcine-induced cytoplasmic protrusion in CB-free solution.
Values in the same column with different superscripts are significantly different.
Effects of CB- or demecolcine-treatment on the G-actin level
The comparison of G-actin levels between the control, demecolcine, and CB-treated oocytes with the Western blot method showed that the G-actin level of CB-treated oocytes was significantly higher than that of demecolcine-treated or control oocytes (Fig. 5). This observation indicates that the cytochalasin treatment method will increase G-actin level of oocytes.
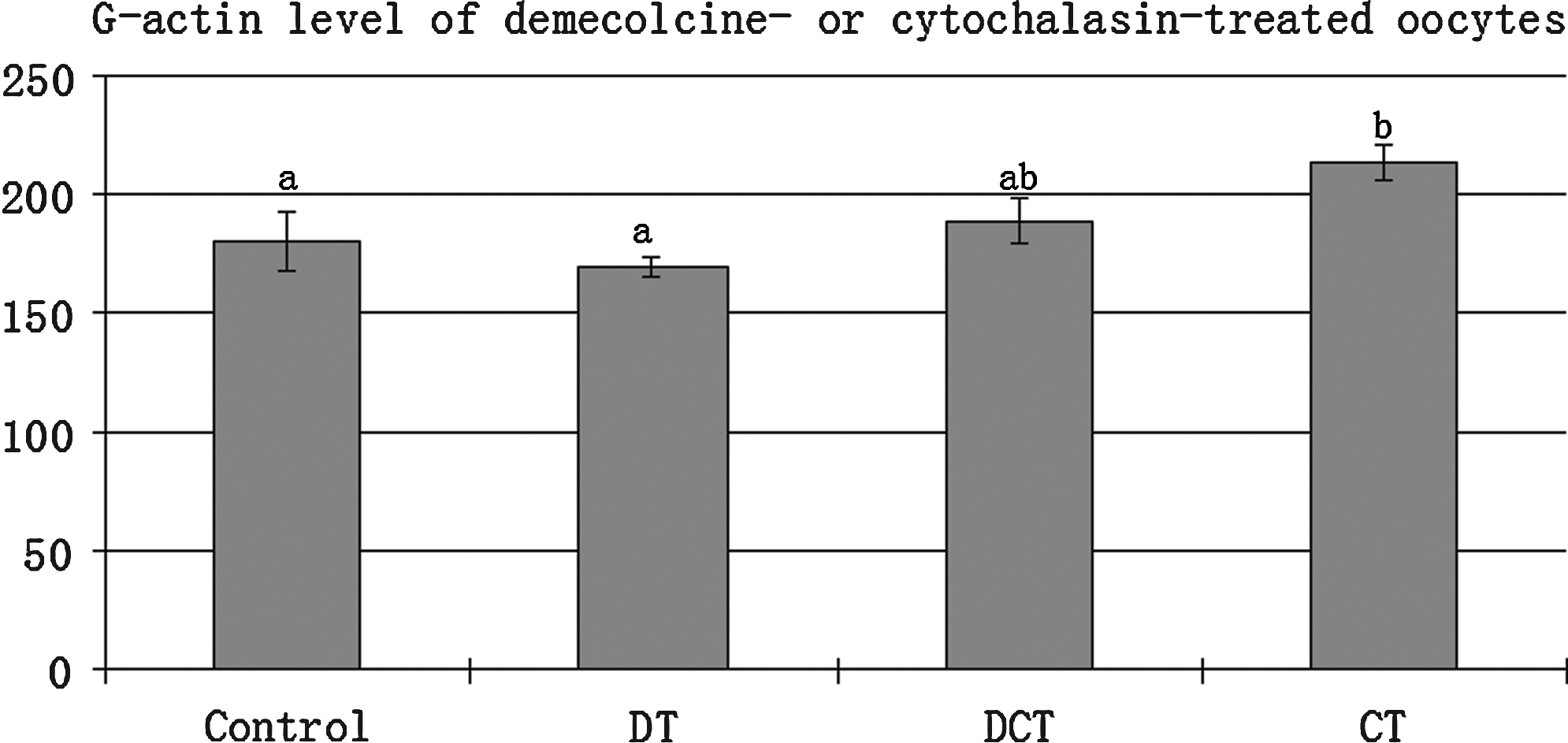
Effects of demecolcine or cytochalasin B treatment on G-actin level in bovine oocytes. G-actin level was increased (p<0.05) in the cytochalasin B-treated (CT) oocytes compared to the demecolcine-treated (DT) or untreated (control) oocytes. G-actin level of Demecolcine + CB-treated oocytes was also higher than that of DT or control groups, but the difference doest not reach statistical level (n=170).
Discussion
Although demecolcine-assisted enucleation in animal cloning has been used in many species, its mode of action still remains unclear. In this study, we investigated the interaction of microtubules and microfilaments, the two major cytoskeleton components in mammalian oocytes, after bovine oocytes were treated with demecolcine. Immunochemical staining of intact oocytes revealed a nuclear area with chromosomes, microtubules, and microfilaments. Our findings and reports from other laboratories indicate (Hou et al., 2006; Lan et al., 2008; Tani et al., 2006) that demecolcine will alter the spindle structure causing a unique cytoplasmic protrusion that contains the chromosomes. We observed that the protrusion comes about with little change to the microfilaments. Our hypothesis is that the membrane protrusion is induced by demecolcine interrupting the interaction between microtubules and microfilaments. The interruption appears to squeeze or compress the nuclear area causing the membrane to protrude, and occurs once the microtubule spindle is dismantled. If our hypothesis is correct, then the demecolcine-induced membrane protrusions withdraws once the demecolcine-treated oocytes have been treated with G-actin polymerization inhibitors. The inhibitors induce a rapid decrease of filamentous actin (microfilaments) and the reduced microfilaments are no longer capable of maintaining the membrane protrusion. The results we observed indicate that the microfilaments were greatly reduced with subsequent disappearance of the demecolcine-induced membrane protrusions. When the demecolcine-treated oocytes were treated with the actin polymerization inhibitor CB, the induced protrusion rapidly vanished. A previous study on goat oocytes also indicated that most demecolcine induced protrusions disappeared within 30 min after exposure to CB (Lan et al., 2008). The G-actin/F-actin dynamics can be transformed between each other by polymerization /depolymerization. Wang et al. (2000) reported in their study that F-actin formation resulted from the polymerization of G-actin into F-actin and was not a result of protein synthesis (Wang et al., 2000). Our Western blot data indicated that cytochalasin treatment increases the G-actin level of oocytes (Fig. 5), which is also consistent with the observation that cytochalasin decreases the F-actin level in oocytes. We conclude that the increase of the global actin level in cytochalasin-treated oocytes is due to the depolymerization of filamentous actin. The cytoskeleton alterations in demecolcine-/cytochalasin-treated oocytes shows an imbalance/interruption between the microtubulin/filament interactions, which is the primary reason for the formation of the membrane protrusion containing the chromosomes.
Because cytochalasins inhibit the polymerization of G-actin and induce rapid depolymerization of filamentous actin (Casella et al., 1981), it has been widely used for the mechanical enucleation of mammalian oocytes in nuclear transfer technology. The findings in this study agree with a fairly recent report by Lan et al. (2008) on enucleated sheep oocytes. Both studies conclude that CB inhibits the formation of demecolcine-induced membrane protrusions. In addition, our results showed that CB could effect oocyte membrane protrusions at concentrations as low as 0.5 μg/mL. The relevance of these observations is that CB hinders the practical application of demecolcine, even when oocytes are only exposed to low concentrations. To overcome the confounding effects of CB, we investigated the potential of mechanical enucleation of bovine oocytes without cytochalasin. We were somewhat surprised at the high survival rate (98.2%) achieved when enucleation was carried out in CB-free medium, which was comparable to CB enucleation (100%). This observation is contrary to the enucleation procedures used for the mouse and rabbit where cytochalasin is necessary (Chesne et al., 2002; Kishigami et al., 2006; Meng et al., 2008, 2009). The difference between these species is more than likely a consequence of the biologic uniqueness of the oocyte. Compared with rodent oocytes, which have fewer lipids and are transparent, the farm animal oocyte has more lipids and appears dark under the light microscope. This unique characteristic makes it easier to operate on the oocyte during enucleation because the oocytes appear to be softer and easier to manipulate with a micropipette. Therefore, based on the results of our study and the unique characteristics of the bovine oocyte, we conclude that the cytochalasin-free demecolcine-assisted enucleation method can be a very efficient method for the enucleation of bovine oocytes.
In our study, enucleation accuracy was higher in the demecolcine-assisted, cytochalasin-free enucleation group than in the traditional blind aspiration cytochalasin group. Blastocyst development rate and blastocyst cell number were also compared between these two enucleation methods. The blastocyst rate and average cell number were significantly higher in the CB-free, demecolcine-assisted group. We believe this was due to the demecolcine-assisted method being more precise, resulting in minimal cytoplasmic loss compared to the traditional blind aspirating method. Previous reports have shown that by reducing the loss of oocyte cytoplasm, blastocyst developmental rates, and blastocyst cell numbers increase (Meng et al., 2003; Westhusin et al., 1996). In addition, demecolcine treatment of oocytes has been reported to enhance in vitro and in vivo development of nuclear transfer embryos (Li et al., 2009).
Demecolcine-assisted/induced enucleation has been widely described; however, its mode of action has not been very well clarified. To provide a better awareness of possible mechanisms involved, we used immunocytochemical staining to investigate the microtubule and microfilament dynamics in bovine oocytes after demecolcine and/or CB treatments and confirmed the microfilament alterations by Western blotting. The results showed that demecolcine-treatment disrupts the balance between microtubule/microfilament; of which, we believe is the primary cause for the demecolcine-induced membrane protrusion. This explanation may also apply for oocyte cytoplasm protrusions induced by other microtubule polymerization inhibitors. Our results also indicate that cytochalasin is not necessary for the enucleation of bovine oocytes, and that demecolcine–enucleation is very efficient in a CB-free medium. Our findings may not only improve upon the efficiency of bovine NT cloning, but may also have application to other farm animal species.
Footnotes
Acknowledgments
This research was supported by the Utah Agriculture Experiment Station (UAES manuscript number 8269) and USDA.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.