
Abstract
Abstract
Because cloned dogs are genetically identical, variations among these animals can be a useful tool to elucidate mechanisms underlying phenotypic differences. To estimate the influence of genetic factors on phenotypic variation, changes in concentration patterns of growth hormone (GH), insulin like growth factor-1 (IGF-1), and IGF binding protein 3 (IGFBP-3) were compared among cloned and age-matched control dogs. In addition, the concentrations of GH and IGF-1 following administration of growth hormone releasing hormone (GHRH) and somatostatin (SRIF) were measured in both groups. In comparing hormone profiles, the control dogs had larger standard deviations from the means for GH, IGF-1, and IGFBP-3 than the clones. Also, the mean concentration of IGFBP-3 in clones was significantly lower than in the controls between 7 to 12 months of age, whereas the IGFBP-3 changes in clones and controls followed the same pattern. GHRH induced increased serum growth hormone concentration both in clones and controls. However, the concentration of IGF-1 was lower in clones than in controls, and larger standard variations were noted in the control group. In conclusion, the measured traits were more homogeneous in cloned animals than in controls, so cloned animals could be valuable for assessing effects of genotype and environment interactions.
Introduction
The somatotropic axis consists of growth hormone (GH), insulin-like growth factors (IGF-1 and -2), their associated carrier proteins, and receptors, and it plays a key role in control of growth and the regulation of metabolism (Breier, 1999; Radcliff et al., 2004; Renaville et al., 2002). The secretion of GH from somatotropes in the anterior pituitary gland is a central part of the somatotropic axis, and two hypothalamic peptides, GH-releasing hormone (GHRH) and somatostatin (somatotropin release-inhibiting factor, SRIF) are the primary regulators of GH secretion (Root and Diamond, 2000). It has been reported that the serum concentration of hormones associated with the somatotropic axis changes markedly during the first year of life and the ontogeny of alteration is known to involve various factors such as genetic variation, sex, age, and diet (Govoni et al., 2003, 2004; Skaar et al., 1994). Accordingly, comparing the effect of hormonal stimulation on the release of GH and IGF-1 in genetically identical versus wild-type individuals can be used to ascertain the influence of genetic and epigenetic factors on hormonal regulation.
Thus, the aim of this study was to study hormonal regulation in cloned dogs to elucidate the influence of genetic and epigenetic factors in response to hormonal stimulation. The changes in concentration of GH, IGF-1, and IGF-binding protein 3 (IGFBP-3) were compared during a period of growth. Also, the concentrations of GH and IGF-1 after GHRH and SRIF administration were measured in cloned dogs and age-matched control dogs.
Materials and Methods
Care and use of animals
Two cloned beagle dogs produced from canine fetal fibroblasts were used (Hong et al., 2009a). Briefly, isolated beagle fetal fibroblasts were used as donor cells and in vivo matured canine oocytes collected surgically from the oviducts of oocyte donor dogs were used as recipient oocytes for SCNT. After SCNT, the reconstructed embryos were surgically transferred into the oviducts of recipient female dogs. Two healthy beagle puppies were delivered from the same surrogate mother on day 60. Genetic identity of cloned dogs were confirmed by microsatellite genotyping and mitochondrial DNA sequences analysis (Hong et al., 2009a). Healthy age-matched beagles (n=4, Marshall Beijing Biotech Ltd., Beijing, China) were used as the control group in this study. The health status of animals was monitored regularly by measurement of hematological and serum biochemical parameters.
All animals were kept in conventional conditions (open system) on the Seoul National University campus under the following environmental conditions: temperature 25–27°C, relative humidity 50–70%, 12 h light:12 h dark cycle. All puppies received standard vaccinations (Vanguard Plus 5CV®, Pfizer Animal Health Korea ud., Korea), and received annual booster injections throughout their lives. None of the dogs was used for breeding during the study. The animal studies were conducted in accordance with recommendations described in “The Guide for the Care and Use of Laboratory Animals” published by the Institutional Animal Care and Use Committee (IACUC) of Seoul National University (approval number SNU-090508-5). Dog care facilities and the procedures performed met or exceeded the standards established by the Committee for Accreditation of Laboratory Animal Care.
Body weight (BW) measurement and blood sample collection
BWs were recorded monthly from 4 to 12 months of age for evaluating growth in the cloned dogs and age-matched controls. Blood samples were collected once per month (4–12 months of age) via jugular venipuncture from clones and controls, and transferred into a serum-separating tube (Becton Dickinson, Franklin Lakes, NJ). These blood samples (3 mL) were collected every 1 h for 6 h (six samples from each animal) and serum was harvested by centrifugation at 1000×g for 10 min and stored at −80°C until assayed.
GHRH and SRIF challenge
GHRH (1–44 amino acids, H-3695, Bachem, Natick, MA) and SRIF (H-4955, Bachem) were dissolved in sterile saline (0.9%) to concentrations of 10 and 5 μg/mL, respectively, and hormonal challenge test were performed at 6 months of age. In Experiment I, cloned animals and controls were challenged with a bolus intravenous infusion of GHRH (5 μg/kg BW) and blood samples were collected via jugular venipuncture in serum-separating tube (Becton Dickinson) at −30, −15, 0, 5, 10, 20, 30, 45, 60, 90, 120, and 180 min relative to GHRH administration. In Experiment II, animals were slowly infused with SRIF (10 μg/kg BW) during 60 min before the GHRH administration, and then GHRH (5 μg/kg BW) was injected intravenously. Blood samples were collected at −75, −40, −10, 0, 5, 10, 20, 30, 45, 60, 90, and 120 min relative to GHRH administration. Samples were centrifuged, and serum was separated and stored at −80°C until analysis.
Hormone analysis
Serum GH and IGF-1 were quantified in all serum samples. Serum GH concentrations were measured by enzyme-linked immunosorbent assay (ELISA) using a Canine growth hormone ELISA kit (Cat. No. E0044c, USCN LIFE Inc., _Wuhan, China), and the minimum detectable dose of canine GH is typically less than 0.26 ng/mL. Serum IGF-1 concentrations were measured by ELISA using a Canine IGF-1 ELISA kit (Cat. No. E0050c, USCN LIFE Inc.); the minimum detectable dose of canine IGF-1 is typically less than 1.56 ng/mL. Serum IGFBP-3 was quantified in two serum samples at each collection time from 4 to 12 months of age. Serum IGFBP-3 concentrations were measured by ELISA using a Canine IGFBP-3 ELISA kit (Cat. No. E0054c, USCN LIFE Inc.) in which the minimum detectable dose of canine IGFBP-3 is typically less than 0.18 ng/mL.
Statistical analysis
The data were subjected to Student's t-test and a protected least significant difference (LSD) test using general linear models (GLM) in a statistical analysis program (SAS Institute, version 9.1, Cary, NC) to determine differences among experimental groups. Differences among the groups were considered statistically significant when the p-values were less than 0.05.
Results
Measurement of BW and age-related changes of the hormones
Comparison of the growth parameters among cloned dogs was performed over the course of the study (Fig. 1). As the clones and controls grew, the estimated BWs increased and statistical differences were observed among two cloned dogs and age-matched controls, except at 8, 9, 10, and 12 months of age. The BWs of the controls were significantly heavier than the clones during 4 to 7 months of age (kg, controls vs. clones; 4 months 6.36±0.07 vs. 5.38±0.23; 5 months 6.47±0.1 vs. 5.8±0.27; 6 months 7.05±0.16 vs. 5.94±0.36; 7 months 7.51±0.1 vs. 6.36±0.47; p<0.05), and then the clones (9.02±0.33) became significantly heavier than controls (8.06±0.15) at 11 months of age (p<0.05).

Body weight change patterns of two clones and age-matched controls. (n=4). Data expressed as mean ±SEM and asterisked values denote a significant difference between the groups (p<0.05).
Profiles of mean plasma GH, IGF-1, and IGFBP-3 concentrations during growth of clones and controls are shown in Figures 2, 3, and 4. In comparing individual GH concentrations, clones demonstrated similar hormonal change patterns with small standard variations throughout the experimental period. Clones showed significant differences only at 4 months of age (Fig. 2A), whereas controls demonstrated significant differences from each other with large variations of GH concentration during the period of growth (Fig. 2B). There were no statistical differences in overall GH concentrations between clones and controls except at 9 months of age (Fig. 2C). As a results of changes in GH concentration profiles in clones and controls, similar patterns were observed in the result of serum IGF-1 concentration (Fig. 3). Also, as a result of changes in GH and IGF-1 concentrations, individuals in the control group demonstrated significantly different serum IGFBP-3 concentrations during the period of growth (Fig. 4B). Changes in IGFBP-3 concentration in clones and controls followed the same pattern, such that the concentration of IGFBP-3 increased from 4 to 5 months of age and exhibited a plateau from 7 to 10 months of age (Fig. 4C). There were significant differences in IGFBP-3 protein levels between clones and controls between 7 and 12 months of age.
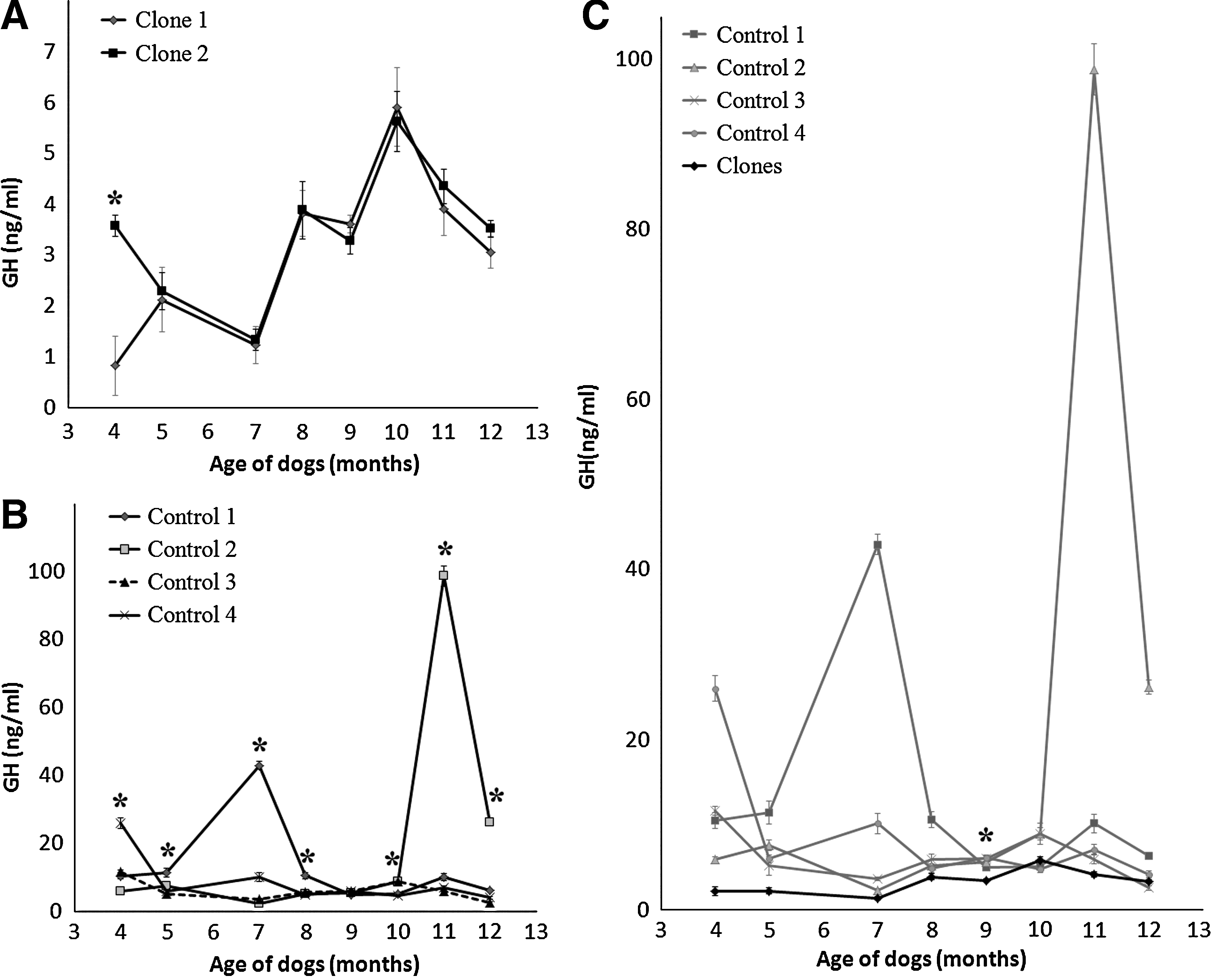
GH concentration profiles in clones and controls measured from 4 to 12 months of sampling. (
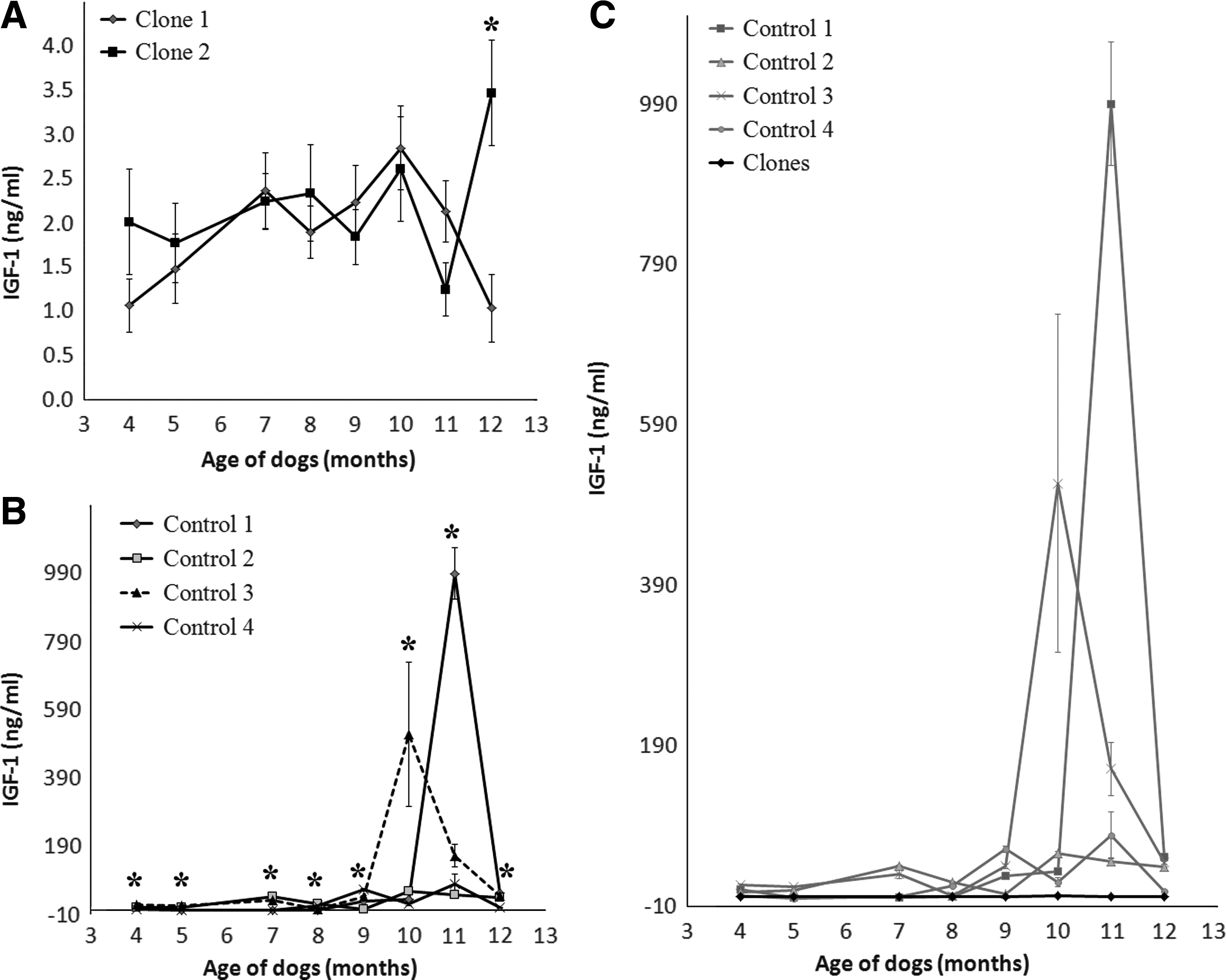
IGF-1 concentration profiles in clones and controls measured from 4 to 12 months of sampling. (

IGFBP-3 concentration profiles in clones and controls measured from 4 to 12 months of sampling. (
GHRH and SRIF challenge
Profiles of mean serum GH and IGF-1 concentrations during intravenous injection of GHRH (Experiment I) or infusion of SRIF followed by intravenous injection of GHRH (Experiment II) are shown in Figures 5 and 6. Relative to pre-GHRH administration, both GHRH and SIRF + GHRH stimulation induced increased GH release in clones and controls as early as 5 min after stimulation. The GHRH stimulation test showed that GH returned to basal levels around 60 min, and the SIRF + GHRH stimulation test demonstrated that the increased GH release lasted approximately 45 min. Overall, serum concentrations of IGF-1 in clones and controls followed the same pattern but the hormonal concentrations in clones were lower than in controls.
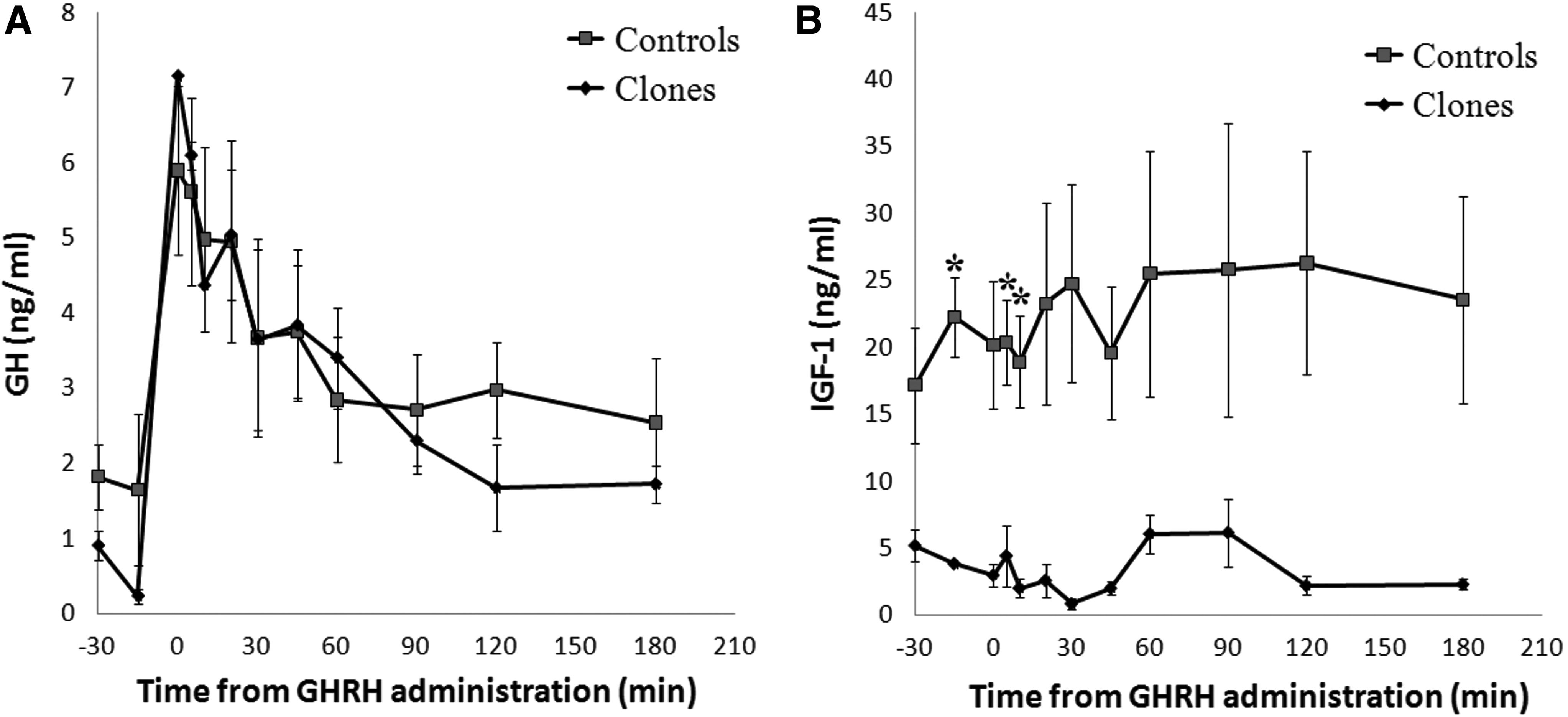
Concentrations of GH and IGF-1 in clones and controls (n=4) after GHRH (5 μg/kg BW) challenge. Data expressed as mean ±SEM and astrisked values denote a significant difference between the groups (p<0.05).
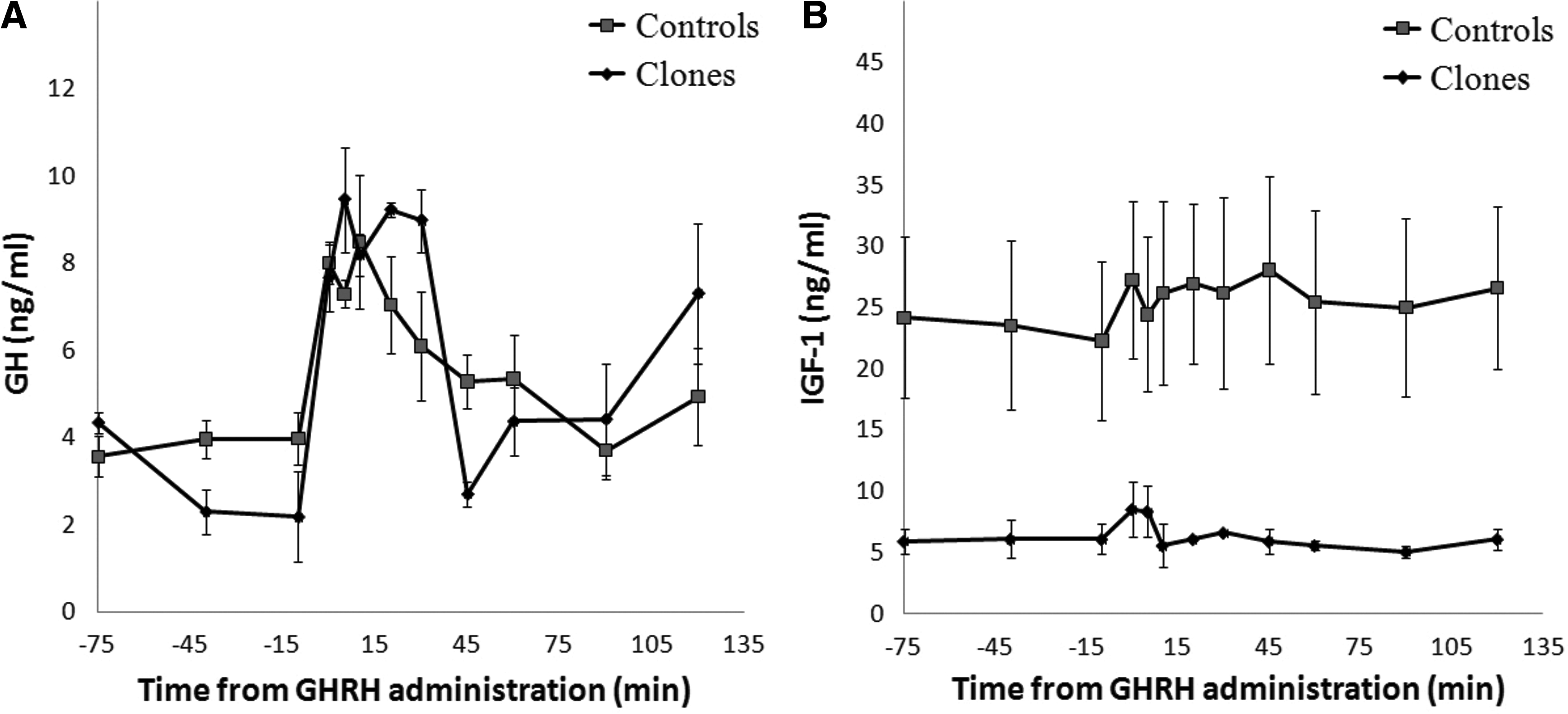
Concentrations of GH and IGF-1 in clones and controls (n=4) after SRIF (10 μg/kg BW) and GHRH (5 μg/kg BW) challenge.
Discussion
Since the birth of the first mammal successfully cloned from an adult somatic cell (Wilmut et al., 1997), production of genetically identical animals is known to be feasible, even though somatic cloning has a low success rate and still needs technical improvement. Nevertheless, the wide application areas for somatic cell cloning have attracted a great deal of scientific interest.
For the biologist, genetically identical individuals have been considered as an important resource for exploring the etiology of disease and for understanding the role of genetic and environmental factors in determining phenotypes (Wong et al., 2005). To elucidate the effects of genetic and environmental factors on phenotypic variation, monozygotic versus dizygotic twins were compared to estimate the degree of genetic contribution to a given phenotype, however monozygotic twins often demonstrate phenotypic discordance (Petronis et al., 2003; Wong et al., 2005). Although it has been reported that the degree of genetic sharing and the degree of phenotypic similarity are correlated (Boomsma et al., 2002; Yang et al., 2002), the substantial discordance between monozygotic twins indicates that chromosomal DNA sequences alone cannot completely determine phenotypic variations such as disease susceptibility (Chakravarti and Little, 2003). The term “epigenetics” is classically used to explain phenotypic events that cannot be described by genetic mechanisms alone and it is known to play a critical role in regulating a variety of genomic functions (Fernandez-Morera et al., 2010). To explore the relative contributions of genes, the environment and other factors to phenotypic variance, genetic sources of variations were minimized by using cloned animals derived from identical somatic cell donors and it can be a useful tool for elucidating underlying mechanism of epigenetic regulation. In this study, two cloned dogs were used for evaluation of genetic effects on growth and hormonal regulation.
In the growth data, a larger variation of BWs was observed in cloned dogs than in controls. Although statistical differences in growth were detected, all animals were healthy throughout the study and grew at a rate that was expected for this breed (Aleman et al., 2000; Swanson et al., 2004). In a study of cloned afghan hounds, the increases in BW and height were similar to each other but the estimated variances of the BW were greater than those of the body height (Park et al., 2010). In addition, Landary et al. (2005) reported that cloned goats tended to be more variable in growth patterns than was expected and they suggested that a possible reason for the variance arose from differences in birth type such as uterine environments among the cloned does and the influence of environment on growth. In the present study, possible effects of different surrogate mothers can be eliminated because the two clones were born as twins to the same surrogate mother. Thus, cloned dogs can be applied in studying other factors such as the influence of environment on postnatal growth and development.
In comparing hormone profiles between the clones and controls, the controls had larger standard deviations from the means for GH, IGF-1, and IGFBP-3 than the clones. Also, in the altered patterns of serum IGF-1 concentrations following stimulation with GHRH between clones and controls, the control dogs demonstrated larger standard variations than the cloned animals, from which we infer that genetic factors are associated with regulation of serum IGF-1 concentration. In accordance with our results, cloned goats exhibited less variation than the controls in circulating IGF-1 and T4 concentrations (Landry et al., 2005). Regarding genetic effects on the variation of phenotypic traits, such as variability of litter size, days to slaughter and backfat thickness, behavior, and female gamete developmental competence were compared between clones and unrelated animals (Heyman et al., 2003; Lamberson, 1994; Savage et al., 2003). It was demonstrated that the overall measured traits were more homogeneous in cloned groups than in controls; thus, cloned animals would be valuable for assessing effects of genotype and environment interactions.
The IGFBP-3 concentrations in the clones were significantly lower than in the controls during 7 to 12 months of age, whereas the IGFBP-3 changes in clones and controls followed the same pattern. In a study of cloned calves, the levels of serum GH and IGFBP-3 did not differ from the controls, but clones had significantly different levels of IGF-1 and IGFBP-3 (Chavatte-Palmer et al., 2002). Because there were no differences in health status, based on hematology and serum biochemistry parameters (data not shown), the low serum IGFBP-3 level in the cloned dogs cannot be considered solely as an adverse effect of SCNT and further study will be needed to more clearly identify the reason for the lower serum IGFBP-3 in clones.
In conclusion, the hormonal regulation profiles of clones were similar to controls and larger standard variations were noted in the control group. The clones demonstrated phenotypic traits with smaller standard variations compared to controls. Thus, genetically identical clones can be a model system for studies on the mechanism of genetic and epigenetic regulation of hormone regulation by controlling for effects of genotype and environment interactions.
Footnotes
Acknowledgments
We thank Dr. Barry D. Bavister (Wayne State University) for his valuable editing of the manuscript. This study was financially supported by NRF (#20100025515, #20100002068), RNL BIO (#0468-20110001), MKE (Grant #10033805-2010-12/ #10033839-2010-12), TS Corporation, the Research Institute for Veterinary Science, and BK21 program for Veterinary Science.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.