
Abstract
Abstract
Valproic acid (VPA), a histone deacetylase inbibitor, has been shown to generate inducible pluripotent stem (iPS) cells from mouse and human fibroblasts with a significant higher efficiency. Because successful cloning by somatic cell nuclear transfer (SCNT) undergoes a full reprogramming process in which the epigenetic state of a differentiated donor nuclear is converted into an embryonic totipotent state, we speculated that VPA would be useful in promoting cloning efficiency. Therefore, in the present study, we examined whether VPA can promote the developmental competence of SCNT embryos by improving the reprogramming state of donor nucleus. Here we report that 1 mM VPA for 14 to 16 h following activation significantly increased the rate of blastocyst formation of porcine SCNT embryos constructed from Landrace fetal fibroblast cells compared to the control (31.8 vs. 11.4%). However, we found that the acetylation level of Histone H3 lysine 14 and Histone H4 lysine 5 and expression level of Oct4, Sox2, and Klf4 was not significantly changed between VPA-treated and -untreated groups at the blastocyst stage. The SCNT embryos were transferred to 38 surrogates, and the cloning efficiency in the treated group was significantly improved compared with the control group. Taken together, we have demonstrated that VPA can improve both in vitro and in vivo development competence of porcine SCNT embryos.
Introduction
However, the efficiency of porcine SCNT was still very low, not only because that the majority of SCNT embryos were lost during different stages of pregnancy, but also because that many born piglets suffered from serious developmental abnormalities and died within a few days. The major reason perhaps lies on the incomplete or faulty epigenetic reprogramming in differentiated donor cells (Ng and Gurdon, 2005; Rideout et al., 2001). In genome reprogramming, posttranslational modifications by acetylation of the N-terminal portion of histones are involved. These modifications alter the higher order chromatin structure to make the DNA either accessible or inaccessible to the regulatory and transcriptional machinery (Lee et al., 1993; McGraw et al., 2003). Therefore, histone acetylation is taken as one of the key epigenetic markers in both cells and SCNT embryos, which would affect the subsequent development of SCNT embryos (Yamanaka et al., 2009); changing the histone acetylation in donor cells and/or SCNT embryos can probably alter the in vitro and in vivo developmental competence of reconstructed embryos. Previous studies have reported that increasing the histone acetylation level in donor cell and/or SCNT embryos can enhance their development ability (Giraldo et al., 2007; Martinez-Diaz et al., 2010; Yamanaka et al., 2009; Zhao et al., 2010). In particular, the HDACi valproic acid (VPA), a cell-permeable, short-chain fatty acid, has been shown to induce reprogramming of mouse fibroblasts with a significantly higher efficiency by three factors: Oct4, Sox2, and Klf4 (Huangfu et al., 2008). In addition, VPA induced reprogramming of primary human fibroblasts with only Oct4 and Sox2, without the need for the oncogenes c-Myc and Klf4 (Huangfu et al., 2008). Therefore, it is necessary to determine whether the VPA treatment has beneficial effects on porcine somatic cell nuclear transfer, especially the effect on the full-term development.
Here we examined the effect of VPA on reprogramming of the pig somatic nucleus in SCNT embryos in terms of histone modification, gene expression, and full-term development.
Materials and Methods
All chemicals were purchased from Sigma Chemical Co. (St. Louis, MO) unless otherwise stated, and all of the solutions and medium were filtered by a 0.22-μm filter. The methods for production of SCNT embryos were largely based on those described by Lai and Prather (2003).
Ethics statement
Animal experiments were done in accordance with the guidelines on animal care and use established by the Jilin University Animal Care and Use Committee.
In vitro maturation
Ovaries in this study were collected from gilts at a local abattoir and transported to the laboratory at 32 to 37°C within 2 to 3 h. The oocyte–cumulus complexes (COCs) were aspirated from ovarian follicles (3–6 mm in diameter) with an 18-gauge needle attached to a 10-cc syringe. Only the COCs with three to five layers of intact cumulus cells and uniform cytoplasm were selected and rinsed three times in TL–HEPES containing 0.1% (w/v) polyvinyl alcohol (PVA), then cultured in vitro in the maturation medium. Maturation media consisted of TCM 199, 0.1% polyvinyl alcohol, cysteine (0.1 mg/mL), epidermal growth factor (10 ng/mL), 0.91 mM Na-pyruvate, 3.05 mM D-glucose, follicle-stimulating hormone (0.5 mg/mL), luteinizing hormone (0.5 mg/mL), penicillin (75 mg/mL), and streptomycin (50 mg/mL).
Preparation of donor cells for SCNT
Briefly, 33- to 35 day-old fetuses were recovered and rinsed in 60, 70, and 80% alcohol in turn, then rinsed three times with phosphate-buffered saline (PBS). After removal of the head, intestine, liver, limbs, and heart, the remaining tissues were finely minced into pieces (1 mm3) using scissors in PBS. Minced tissue were then digested with Collagenase (200 IU/mL) and Dnase I (25 KU/mL) in Dulbecco's Modified Eagle Medium (DMEM; Gibco, Grand Island, NY) plus 20% fetal bovine serum (FBS; PAA, Austria) for 2 to 3 h at 39°C and 5% CO2 in air. After digestion and washing, the cell pellet was seeded in a 10-cm culture plate (Corning, Corning, NY) and cultured until confluence. Then cells were frozen with FBS containing 10% DMSO. One or 2 days before nuclear transfer, cells were thawed and cultured.
Production of SCNT embryos
Briefly, COCs were pipetted repeatedly in 0.1% hyaluronidase to remove the cumulus cells, at 42 to 44 h after maturation. Oocytes having an extruded first polar body with round-shape and intact cytoplasm were selected. Metaphase II (MII) oocytes were placed in the manipulation medium containing 7.5 mg/mL cytochalasin B, and enucleation was accomplished by aspirating the polar body and MII chromosomes with a small amount of surrounding cytoplasm using a beveled glass pipette. Consequently, a single donor cell with proper size and good morphology was introduced into the perivitelline space and placed adjacent to the recipient cytoplasm. The fusion and activation of cell-cytoplast complexes were accomplished with 2 DC pulses of 1.2 kV/cm for 30 μsec on a BTX Electro Cell Manipulator 2001 (BTX, San Diego, CA) in a chamber full filled with fusion medium (0.3 M mannitol, 1.0 mM CaCl2·2H2O, 1.0 mM MgCl2·6H2O, and 0.5 mM HEPES, pH adjusted to 7.0–7.4). Embryos were then washed and incubated in porcine zygote medium-3 (PZM3) supplemented with 3 mg/mL bovine serum albumin (BSA), and the fusion rate was evaluated 30 to 40 min later. Approximately 50 reconstructed embryos were then transferred into each droplet containing 100 μL PZM3 with 3 mg/mL BSA covered with mineral oil, and cultured at 39°C, 5% CO2 in air. Each experimental group, 24 h, 48 h cleavage, and blastocyst rates were evaluated, and the number of cells in blastocyst was examined under UV light after staining with 10 μg/mL of Hoechst 33342 on day 5.
Preparation of VPA
Valproic acid was dissolved in PZM3 at 40 mM, and the concentrated stock solution was stored at 4°C (no more than 2 weeks). The VPA stock solution was added to the PZM3 at different concentrations according to the experimental procedure. After treatment, the culture medium was changed to PZM3 without VPA.
Detection of histone acetylation
An indirect immunofluorescence analysis was performed to determine histone acetylation. Briefly, samples were rinsed in PBS and then fixed in 4% (w/v) paraformaldehyde in PBS for 30 min at room temperature. After permeabilization with 0.2% Triton X-100 in PBS for 30 min, samples were blocked in 5% goat serum in PBS for 30 min at room temperature. Consequently, the samples were incubated with a rabbit polyclonal primary antibody to acH3K14 (Abcam, diluted in 1:50) or acH4K5 (Abcam, diluted in 1:250) overnight at 4°C. After washing intensively with 0.2% (v/v) Tween-20 in PBS, the samples were incubated with Alexa Fluor 594 goat antirabbit IgG (Invitrogen, Carlsbad, CA) diluted in 1:800 for 1 to 2 h at 39°C. After washing with PBS, embryos and cells were mounted on slides in mounting medium containing Hoechst 33342 for 5 min. Samples were analyzed under an epifluorescent microscope (Nikon, Tokyo, Japan) equipped with a digital camera. Images were captured with the same exposure times and adjustments of the microscope and the quantification of images was done with an image analyzer system, Image-Pro Plus (Media Cybernetics, Bethesda, MD). Then, the average ratios of acH4K5 signals were used for the comparison between the different groups.
Total RNA isolation and real-time PCR amplification
Single blastocyst's total RNA isolation and reverse transcription was done according to previous report (Kurimoto et al., 2007). The quantification of all gene transcripts was carried out in three replications using BioEasy SYBR Green I Real-Time PCR Kit. Briefly, the reaction mixture of the total 50 μL volume was consisted of 25 μL 2×SYBR Mix (including PCR buffer, Mg2+, dNTP mixture, SYBR Green I, etc.), 1 μL of each primer, 0.3 μL Taq polymerase, 4 μL template, and 18.7 μL ddH2O. The primer sequences for each gene were shown as follows:
Oct4 Forward, 5′-AAGCAGTGACTATTCGCAAC-3′ Reverse, 5′-CAGGGTGGTGAAGTGAGG-3′ Sox2 Forward, 5′-CGCAGACCTACATGAACG-3′ Reverse, 5′-TCGGACTTGACCACTGAG-3′ Klf4 Forward, 5′-GTTCTCATCTCAAGGCACACC-3′ Reverse, 5′-TCGCACTTCTGGCACTGG-3′ β-actin Forward, 5′-GAACCCCAAAGCCAACCGT-3′ Reverse, 5′-CCTCGTAGATGGGCACCGT-3′
Embryo transfer
The SCNT embryos were reconstructed with normal fibroblasts and were treated or nontreated with 1 mM VPA for 14 to 16 h after activation. Then the SCNT embryos were transferred to the oviducts, on the day of, or 1 day after the onset of estrus. Pregnancy was diagnosed on days 23–27 (the day of SCNT was taken as day 0).
Statistical analysis
All data were obtained in at least three replicates. Developmental rates were analyzed by a chi-squared test; other data were analyzed by analysis of variance (ANOVA), and a probability of <0.05 was considered statistically significant.
Results
Optimization of VPA treatment for pig SCNT embryos
In order to determine the optimized VPA concentration in the treatment medium, SCNT embryos using normal Landrace fetal fibroblasts as donor cells were cultured in modified PZM3 supplemented with 0, 0.1, 1, 4, or 10 mM VPA for 14 to 16 h. The rates of blastocysts formation were significantly higher in the 1 mM-treated group compared with the 0 mM group (31.8 vs. 11.4%; Table 1). This optimized concentration and treated time duration was used in further experiment.
Values with different superscripts are significantly different.
Fluctuation in acetylation level of histone H3 lysine 14 and histone H4 lysine 5 in SCNT embryos
To determine the effect of the VPA treatment, we compared the acetylation levels of histone H3 lysine 14 and histone H4 lysine 5 in fibroblasts and SCNT embryos (Fig. 1). To confirm the difference, the average optical intensity of acH3K14 and acH4K5 was measured using Image-Pro Plus. In the first experiment, fibroblasts were treated with or without 1 mM VPA for 14 to 16 h. Results indicated that the acetylation levels of both histone H3 lysine 14 and histone H4 lysine 5 were significantly higher in the VPA-treated group in comparison with the nontreated control. Next, the SCNT embryos, reconstructed with untreated fibroblasts, were treated or untreated with 1 mM VPA for 14 to 16 h. At both pronuclear or pseudopronuclear and two-cell stages, the acetylation levels of both histone H3 lysine 14 and histone H4 lysine 5 were significantly higher in the VPA-treated groups than that in the control groups (Fig. 2). However, at the blastocyst stage, no significant difference in both acH3K14 and acH4K5 acetylation level was observed among the groups (Fig. 3). This is in the agreement with the results of the immunofluorescence analysis (Fig. 4).
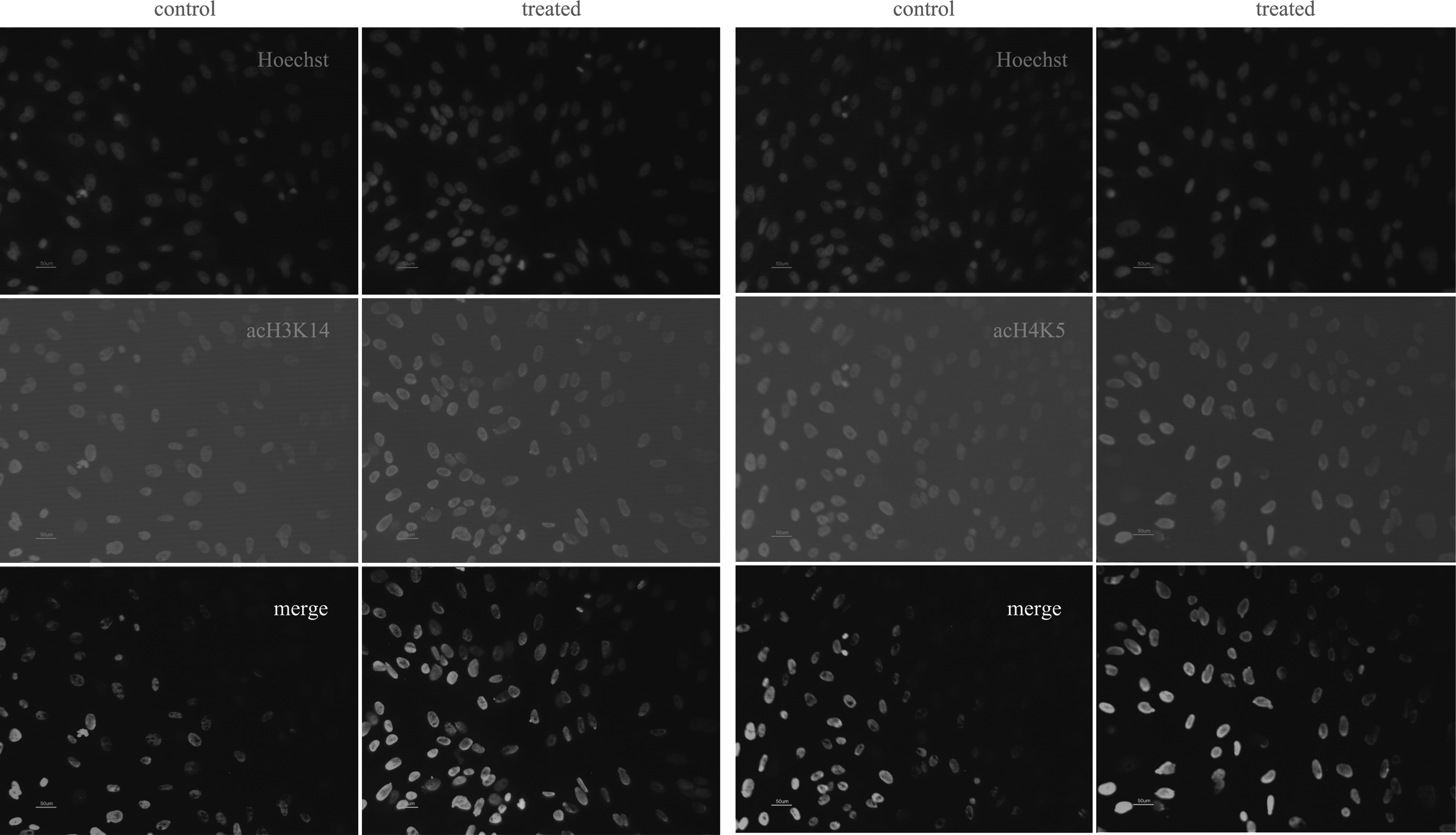
Acetylation levels of histone H3 lysine 14 and histone H4 lysine 5 in fetal fibroblasts. The fibroblasts were labeled for acH3K14 or acH4K5 and DNA (Hoechst 33342). Control, nontreated fibroblasts; treated, fibroblasts treated with VPA.

Acetylation levels of histone H3 lysine 14 and histone H4 lysine 5 at pronuclear or pseudopronuclear and two-cell stages. The SCNT embryos were labeled for acH3K14 or acH4K5 and DNA (Hoechst 33342). Control, nontreated SCNT embryos; treated, SCNT embryos treated with VPA.

Acetylation levels of histone H3 lysine 14 and histone H4 lysine 5 at the blastocyst stage. The blastocysts were labeled for acH3K14 or acH4K5 and DNA (Hoechst 33342). Control, nontreated blastocysts; treated, blastocysts treated with VPA.
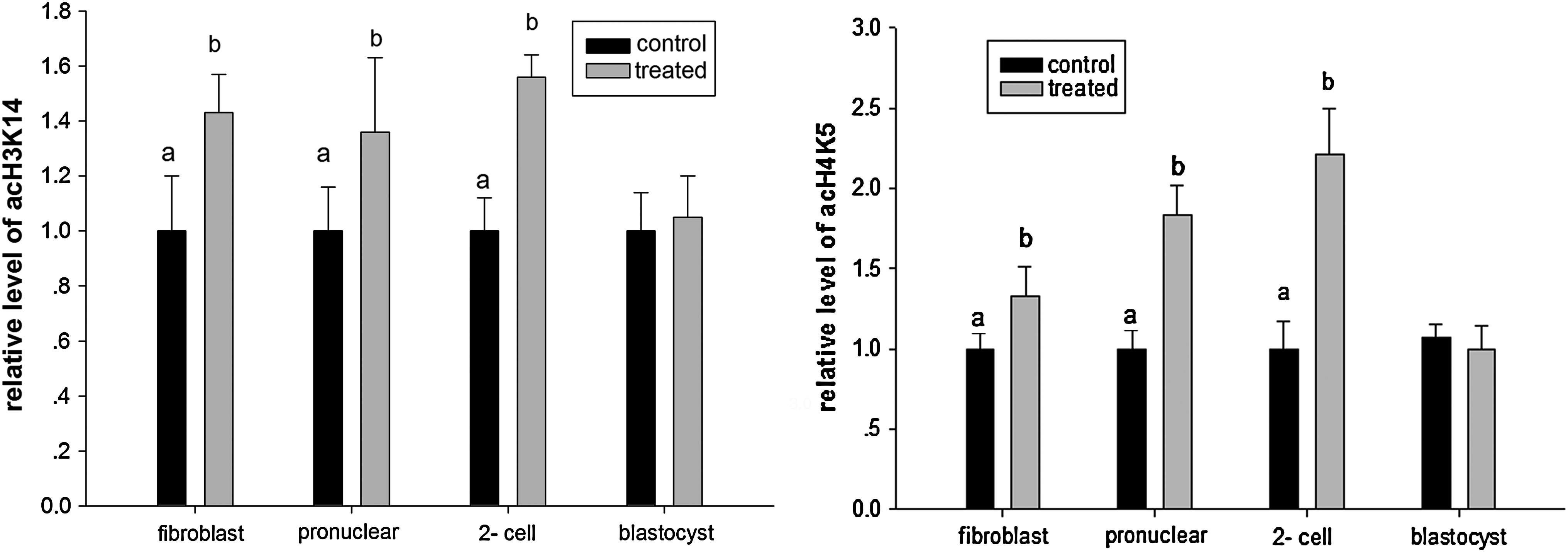
Relative level of acH3K14 and acH4K5 in fetal fibroblast cells or SCNT embryos without (control) or with (treated) VPA treatment;
Investigation of pluripotency-related gene expression in SCNT embryos with or without VPA treatment
To determine whether the increased blastocyst rate with VPA treatment was correlated with the altered pluripotency-related gene expression, the expression levels of Oct4, Sox2, and Klf4 were studied using real-time PCR at the blastocyst stage. As shown in Figure 5, there are no significant differences in the relative expression levels of Oct4, Sox2, and Klf4 between the nontreated and treated embryos. Taken together, the gene expression was probably not affected with VPA treatment at the blastocyst stage.

Relative expression level of Oct4, Sox2, and Klf4 in blastocysts derived from SCNT embryos without (nontreated) or with (treated) VPA treatment. Each bar represents the relative fold change between treated and untreated group.
Study on the effect of VPA on the full-term development of SCNT embryos
In order to better understand the effect of VPA on the in vivo developmental competence of porcine SCNT embryos, the embryo transferring was performed. As shown in Table 2, pregnancies in 14 of 20 surrogates (70%) were detected in the VPA-treated group, and 19 piglets were born from seven litters with cloning efficiency of 0.40%. As for the nontreated group, 6 of 18 surrogates (33%) became pregnant and eight piglets were delivered with cloning efficiency of 0.18%. It should be noted that, the pregnant rate and/or cloning efficiency of both male and female cell strains were higher in the VPA-treated group than the control group, respectively. Abortion and fetal resorption was taken place during gestation; however, the ratio of the abortion and fetal resorption number to the pregnant number was the same (7/14 vs. 3/6). Taking together, VPA significantly improved the cloning efficiency of porcine SCNT embryos in this study.
+: Pregnant; −: not pregnant.
Abortion or fetal resorption.
Average numbers of embryo transferred.
Cloning efficiency: No. of piglets/No. of embryos transferred.
Discussion
Because of several similarities with humans, porcine SCNT to generate cloned embryos is of great importance in many biomedical applications (Prather et al., 2003). Many efforts have been placed on genetically modifying the pig genome through SCNT, to make the pig organs immunologically more compatible to humans (Dai et al., 2002; Lai et al., 2002; Liang et al., 2011; Shimizu et al., 2008). However, the generation of pigs through SCNT has been proven to be not so easy. As we known, the breeding of piglets is a long and costly process, because of the long breeding cycle. Furthermore, possibly due to the inefficiencies of in vitro oocyte maturation and/or embryo culture, the cloning efficiency is still low. In the present study, we demonstrated that the VPA, a histone deacetylase inbibitor, had a beneficial effect on both preimplantation and full-term development of porcine SCNT embryos.
In the miniature pig, a treatment at 4 mM VPA for 48 h significantly increased the blastocyst formation rate of SCNT embryos compared with the control (Miyoshi et al., 2010). In mice, a treatment at 2 mM for 2 to 3 h after nuclear transfer and continued 6 h following activation was considered as an optimal condition for preimplantation development of SCNT embryos (Costa-Borges et al., 2010). The optimal conditions determined in these studies presented a higher concentration of VPA compared with the present study. Here, it showed that VPA treatment at 1 mM for 14 to 16 h from the start of activation was sufficient to significantly improve both porcine blastocyst formation rate and full-term developmental competence. These results probably suggested that VPA can be effective in a wide range of concentration and time duration, and there possibly be different in the VPA treatment among different species. Certainly, even to the same specie, the optimal VPA treatment may be differed due to different SCNT protocols and/or culture medium in different laboratories. Previous studies have reported that nuclear transfer protocol would affect the messenger RNA expression, and the different culture condition can also affected the developmental potential (Wrenzycki et al., 2001). As shown in Figure S1 and Figure S2 (Supplementary Data are available online at www.liebertonline.com/cell), the morphology of both fibroblasts and blastocysts between VPA-treated and nontreated groups did not differ largely. Also, the proliferation rates of fibroblasts in both treated and control groups were not significantly different (Table S1). From these aspects, VPA treatment was safe for porcine SCNT.
The acetylation levels of both histone H3 lysine 14 and histone H4 lysine 5 in the VPA-treated group in SCNT embryos at pronuclear or pseudopronuclear and two-cell stages, but not the blastocyst stage, were significantly higher than those in the control groups, suggesting that VPA treatment is possibly mainly effective in the early preimplantation stages. As we known, the four-cell stage is a genome activating period in the pig. Therefore, a higher acetylation level at the early stages may contribute to go through the period for most embryos much more completely and easily. Because both histone acetylation and deactylation played very important roles in reprogramming and contribute to the success of SCNT (Kishigami et al., 2006), and hyperacetylation level of histones facilities some factors access to nuclesomes (Lee et al., 1993). It should be noted that, although the levels of both acH3K14 and acH4K5 were significantly higher in the treated groups than that in the control groups, the levels between them were different. It was probably suggested that the acetylation levels were varied in different acetylation sites of histones.
Embryos lacking Oct4 gene expression cannot form normal blastocysts (Nichols et al., 1998), so a promoted expression of the Oct4 gene in the SCNT embryos at a certain range was expected to improve the quality of the blastocyst. However, in the present study, the real-time PCR on Oct4, Sox2, and Klf4 showed that a slight fluctuation existed, but was not significant higher or lower. Therefore, the enhanced implantation developmental competence was perhaps not through changing the quantity of pluripotency-related gene expressions. Combined with the results of actylation detection, VPA treatment may possibly mainly rise the population level in contrast to a single embryo quantity at the blastocyst stage. It should be noted that, in the VPA-treated groups, the first blastocyst appeared as early as on day 4, which was earlier than that in the nontreated groups. The promoted developmental process may also lead to the fluctuations in the experiment. At any rate, it has to be acknowledged that in vitro culturing is not equal to the in vivo development system, and the cloning efficiency should be taken as stricter criteria.
Although abortion and fetal resorption was taken place during gestation, the positive effects on the in vivo development of porcine SCNT embryos with VPA treatment should not be neglected because abortion and fetal resorption is a multifactorial event. As there is a minimal necessity for at least four good-quality embryos to maintain a pregnancy (Jiang et al., 2008), suggesting that more good-quality embryos were harvested after VPA treatment. It has to be pointed out that the sample size was still not large enough and the generation of piglets through somatic cell nuclear transfer is far from being a stable production line. As a result, some uncertain factors probably contribute to the fluctuation of cloning efficiency and a further exploitation should be performed.
Taken together, VPA played a positive role on the in vitro and in vivo developmental competence of porcine SCNT embryos. Further studies will need to be performed to determine much more exact effects of VPA on the regulation of gene expression and postnatal development, as well as the combined effects on the pig SCNT embryo production with other small compounds to harvest a much higher cloning efficiency.
Footnotes
Acknowledgments
The authors express their gratitude to all the members of our laboratory for technical support and helpful discussion, and the staff at Embryo Engineering Center for technical assistance and taking care of the animals. This work was supported by the National Key Basic Research and Development Program of China (973 Program) (No. 2009CB941000).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.