
Abstract
Abstract
Our previous study demonstrated the direct involvement of the HIF-1α subunit in the promotion of cardiac differentiation of murine embryonic stem cells (ESCs). We report the use of cobalt chloride to induce HIF-1α stabilization in human ESCs to promote cardiac differentiation. Treatment of undifferentiated hES2 human ESCs with 50 μM cobalt chloride markedly increased protein levels of the HIF-1α subunit, and was associated with increased expression of early cardiac specific transcription factors and cardiotrophic factors including NK2.5, vascular endothelial growth factor, and cardiotrophin-1. When pretreated cells were subjected to cardiac differentiation, a notable increase in the occurrence of beating embryoid bodies and sarcomeric actinin-positive cells was observed, along with increased expression of the cardiac-specific markers, MHC-A, MHC-B, and MLC2V. Electrophysiological study revealed increased atrial- and nodal-like cells in the cobalt chloride-pretreated group. Confocal calcium imaging analysis indicated that the maximum upstroke and decay velocities were significantly increased in both noncaffeine and caffeine-induced calcium transient in cardiomyocytes derived from the cobalt chloride-pretreated cells, suggesting these cells were functionally more mature. In conclusion, our study demonstrated that cobalt chloride pretreatment of hES2 human ESCs promotes cardiac differentiation and the maturation of calcium homeostasis of cardiomyocytes derived from ESCs.
Introduction
The effects of direct reduced-oxygen treatment on cardiac differentiation of human ESCs are inconsistent and nonreproducible. Ezashi et al. (2005) showed that reduced oxygen tension prevents differentiation of human ESCs. On the other hand, Chen et al. (2009) demonstrated that 5% oxygen culture is not beneficial for the maintenance of human ESCs in an undifferentiated state with short splitting intervals. Despite the fact that reduced oxygen treatment may enhance cardiac differentiation of ESCs (Niebruegge et al., 2009), the number of consequent viable cardiomyocytes may be limited by poor cell survival during prolonged culture with reduced oxygen tension treatment (Ng et al., 2010). Other methods such as pharmacological treatment with divalent ions such as cobalt (II) and nickel (II) ions (Bunn and Poyton, 1996) to stabilize the HIF-1α subunit may thus provide an attractive alternative. In fact, cobalt chloride has been used in stabilizing the HIF-1α in both cell culture and animal models in various studies. For example, application of cobalt chloride has been shows to increase the HIF-1α stability in acute myeloid leukemic cells (Huang et al., 2003) as well as human nasal polyp fibroblasts (Lin et al., 2008), whereas short-term cobalt administration caused HIF-1 in the kidney tubules in a rat model (Matsumoto et al., 2003). The exact reason accounted for hypoxia mimicking property of cobalt chloride remains inconclusive. Because the degradation of HIF-1α requires prolyl hydroxalation and such process utilizes iron ion as cofactor, it is likely that cobalt chloride increases the HIF-1α stability via its iron antagonistic action (Kaelin, 2002; Kivirikko and Myllyharju 1998).
In the present study, we sought to investigate the use of cobalt chloride to stabilize the HIF-1α protein at normal oxygen levels to enhance cardiac differentiation and maturation of human ESCs. The results of this study provide new insight in the development of a novel strategy to improve the differentiation and functional property of cardiomyocytes derived from human ESCs.
Materials and Methods
Maintenance and cardiac differentiation of hES2 cells
Human ESC, the hES2 line, was obtained from the WiCell Research Institute (Madison, WI). The undifferentiated cells were maintained in a serum- and feeder-free environment using the StemPro hESC SFM system (Invitrogen, Carlsbad, CA) according to the manufacture's instruction. Where necessary, hES2 cells (at 60% confluence) were treated with cobalt chloride (at the final concentration of 50 μM) for 3 days prior to cardiac differentiation. For cardiac differentiation, when cells reached 80% confluence, cell colonies were treated with dispase (1 mg/mL) for 5 min at 37°C and scraped off into small aggregates. The cell aggregates were allowed to become embryoid bodies (EB) by being cultured in suspension in the StemPro hESC medium for 5 days. After suspension, EBs were plated onto a gelatin-coated plate in a basal medium [StemPro34, Invitrogen, containing 2 mM glutamine, 4 mM monothioglycerol (MTG), 50 μg/mL ascorbic acid, and 0.5 ng/mL bone morphogenetic protein-4 [BMP4]) according to previous protocol (Yang et al., 2008). The following concentrations of factors were used for primitive-streak formation and for mesoderm induction for cardiac differentiation: BMP4, 10 ng/mL; human basic fibroblast growth factor (bFGF), 5 ng/mL; activin A, 3 ng mL−1, and human DKK1, 150 ng/mL. Factors were added as follows: days 1–4, BMP4, bFGF, and activin A; days 4–8, DKK1; after day 8, DKK1 and bFGF. The bFGF was purchased from Peprotech (Rockyhill, NJ), DKK1 from Gibco (Grand Island, NY). and activin-A from Stemgent (San Diego, CA).
Western blot analysis
The total proteins from hES2 cells were isolated as described previously (Ng et al., 2010, 2011). Fifty micrograms of total soluble protein was resolved on 10% PAGE and transferred onto a PVDF membrane electrically. After incubating in the blocking solution (1×Tris-buffered saline, 0.05% Tween-20, 5% nonfat powdered milk), the membrane was incubated with the primary antibody as indicated in the figure at 4°C for 16 h. Unbound antibodies were removed by three washes in washing buffer (1×Tris-buffered saline, 0.05% Tween-20), and the membrane incubated with an HRP-conjugated antimouse IgG secondary antibody (GE Healthcare, Piscataway, NJ) at room temperature for 1 h. Free antibodies were removed by three washes in washing buffer, and the presence of the protein–antibody complex was visualized using the ECL Plus Western blotting detection reagents (GE Healthcare).
End-stage PCR and real-time quantitative PCR analysis
Total RNA was isolated and reverse transcribed using the illustra RNAspin Mini Kit (GE Healthcare): 0.5 μg was reverse-transcribed into first-strand cDNA using the QuantiTect Rev. Transcription Kit (Qiagen, Chatsworth, CA). Expression of the genes of interest was evaluated by end-stage or real-time quantitative amplification using 0.2 μL of the cDNA as a template using the respective oligonucleotides (see Tables 1 and 2). Quantitative PCR analysis was performed with DNA engine opticon 2 real-time PCR Detection System (Bio-Rad, Hercules, CA) using the iQ SYBR Green Supermix (Bio-Rad). For amplification, after initial holds for 5 min at 50°C, 50 cycles of 95°C for 15 sec, followed by 57°C for 30 sec and 72°C for 30 sec, melt curve analysis was performed. The relative quantification of the PCR products was performed according to the 2−ΔΔCt method, using ribosomal protein S16 (S16) as internal control. Where ΔΔCt=[(Cttarget gene − CtS16)CoCl2 treated group − (Cttarget gene −CtS16) control group]].
Flow cytometry analysis
The percentage of ESC-derived cardiomyoyctes was quantified by FACS analysis. Briefly, EBs were dissociated with 1 mg/mL collagenase B (Roche Applied Sciences, Penzberg, German), washed with differentiated medium, and the cells permeabilized by Cytofix/Cytoperm permeabilization kit (BD Biosciences, San Diego, CA). Cells were then stained with monoclonal antisarcomeric actinin antibody (A7811, Sigma, St. Louis, MO) using antimouse IgG H+L-PE as a secondary antibody stain (Beckman Coulter, Fullerton, CA). Analysis was performed with a Beckman Coulter FC500 flow cytometer in which at least 1000 events were counted.
Isolation of hES2-derived cardiomyocytes
Isolation of hES2-derived cardiomyocytes has been described previously (Au et al., 2009; Lieu et al., 2009; Liu et al., 2007, 2009). Briefly, beating outgrowths were microsurgically dissected from hES2-derived EBs (25 days) with a glass knife, followed by incubation in collagenase B (1 mg/mL, Roche Applied Sciences) with DNase I (60 U/mL, Roche Applied Sciences) at 37°C for 30 min with occasional rocking. Isolated cells were recovered in Kraftbrühe (KB) solution containing (mM) 85 KCl, 30 K2HPO4, 5 MgSO4, 1 EGTA, 2 Na2-ATP, 5 pyruvic acid, 5 creatine, 20 taurine, and 20 D-glucose at room temperature for 1 h. The cells were subsequently plated on 0.1% gelatin coated glass cover slips in 24-well culture plates with corresponding differentiation medium. Immunohistochemistry, electrophysiological study, and confocal calcium imaging of isolated cells or cell clusters were performed within 2 days.
Immunohistochemistry
The hES2-derived cardiomyocytes were fixed with 2% paraformaldehyde for 15 min at 4°C followed by two washes with phosphate-buffered saline (PBS). Prior to overnight incubation with murine monoclonal anticardiac Troponin-T antibody (NeoMarker, Fremont, CA) or monoclonal antisarcomeric actinin antibody (A7811, Sigma) at 4°C, fixed cells were blocked in 2% bovine serum alubmin (BSA), PBS, and 0.1% Triton X-100 for 2 h. After removal of unbound antibodies with washing buffer, cells were incubated with Alexa-488 conjugated antimouse secondary antibodies (Invitrogen, Carlsbad, CA) (1:100 dilution in washing buffer) for 1 h at room temperature. Cells were then mounted on glass slides with a mounting medium containing DAPI. The presence of the immune complex was examined by fluorescent microscopy.
Electrophysiological study
For electrophysiological characterization of hES2-derived cardiomyocytes, the standard whole-cell patch-clamp recording was performed at 37±0.5°C to record the action potential phenotypes (HEKA Instruments Inc., Southboro, MA) as previously described (Chan et al., 2009). In brief, patch pipettes were prepared from 1.5-mm thin-walled borosilicate glass tubes using a Sutter micropipette puller P-97 and had typical resistances of 3–4 MΩ when filled with an internal solution containing (mM): 110 K+ aspartate, 20 KCl, 1 MgCl2, 0.1 Na-GTP, 5 Mg-ATP, 5 Na2-phosphocreatine, 5 EGTA, 10 HEPES, and pH adjusted to 7.3 with KOH. The external Tyrode's bath solution consisted of (mM): 140 NaCl, 5 KCl, 1 MgCl2, 0.4 KH2PO4, 1.8 CaCl2, 10 Glucose, 5 HEPES, with pH adjusted to 7.4 with NaOH. For those hES2-derived cardiomyocytes that were electrically quiescent, a stimulation of 0.1–1 nA for 5 msec was given to elicit an action potential. The sampling frequency was 2.00 kHz, and data were corrected for the liquid junction potentials of +20.3 mV.
Confocal calcium imaging
Confocal calcium imaging was performed with the protocol described previously (Ng et al., 2010, 2011). In brief, the isolated hES2-derived cardiomyocytes were loaded with 5 μM Fluo-3 AM (Invitrogen) for 25 min at 37°C in Tyrode solution containing 140 mM NaCl, 5 mM KCl, 1 mM MgCl2, 1.8 mM CaCl2, 10 mM glucose, and 10 mM HEPES at pH 7.4. Spontaneous calcium transient of single or clusters of ESC-derived cardiomyocytes was recorded with a confocal imaging system (Olympus Fluoview System version 4.2 FV300 TIEMPO) mounted on an upright Olympus microscope (IX71). To record the caffeine-induced calcium transient, 20 μL of 10 mM caffeine was applied onto the cell surface during measurement. To evaluate the effect of ryanodine, spontaneous calcium transients were recorded before and after the incubation (5 min) of Tyrode solution containing 10 μM ryanodine. The calcium transient was quantified as the background subtracted fluorescence intensity changes normalized to the background subtracted baseline fluorescence. Data were fed into the Felix 32 (PTI) software for analysis.
Statistical analysis
Continuous variables are expressed as mean±SEM. Statistical comparisons were performed using a Student's t-test. Calculations were performed with GraphPad Prism 5 (GraphPad, LaJolla, CA). A p-value <0.05 was considered statistically significant.
Results
Cobalt chloride treatment increases the protein levels of HIF-1α in hES2 human ESCs
To evaluate the possibility of using cobalt chloride to activate the HIF-1 pathway in hES2 cells, expression of the HIF-1 pathway components in undifferentiated hES2 cells was evaluated using end-stage PCR analysis. As shown in Figure 1A, both HIF-1α and HIF-1β subunits as well as HIF-2α were expressed after cobalt chloride treatment but there was no detectable HIF-3α. These findings suggest that cobalt chloride treatment activates expression of the HIF-1α and HIF-2α signal in hES2 cells. The expression of other components of the HIF-1 pathway, including the prolyl hydroxylases termed oxoglutarate-dependent proline 4-hydroxylases, EGLN-1 to 3 were also observed.
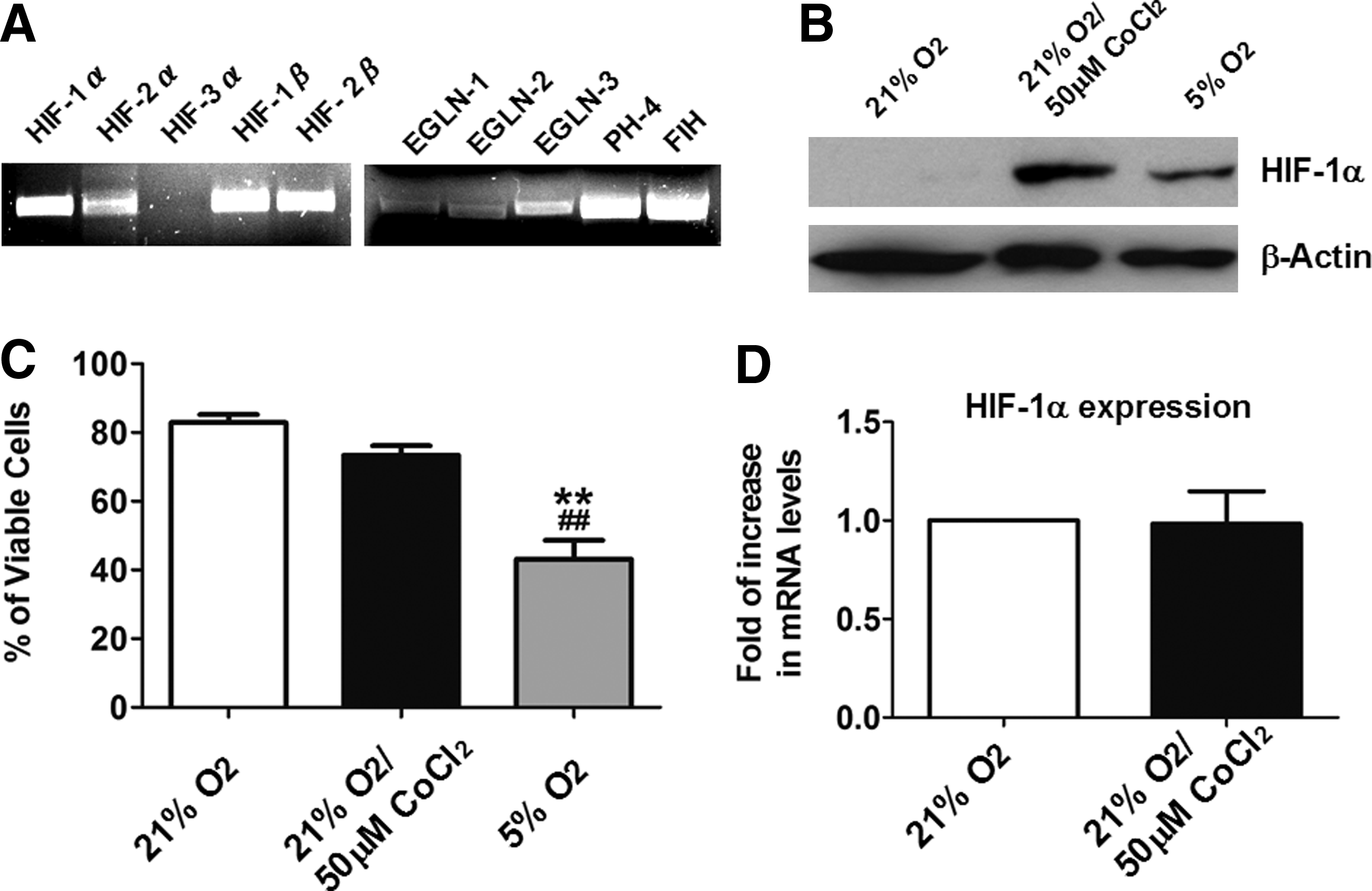
Effect of cobalt chloride on undifferentiated hES2 ESCs. (
Undifferentiated hES2 cells were then subjected to treatment with cobalt chloride (50 μM) or reduced oxygen (5% O2) followed by Western blot analysis to evaluate the protein levels of HIF-1 α. As shown in Figure 1B, both 72 h of cobalt chloride treatment and 24 h of reduced oxygen treatment remarkably increased cell HIF-1α protein levels. Nonetheless, the cell viability of the reduced oxygen-treated groups was thus significantly reduced to about 50% (Fig. 1C). There was no such reduction in cell viability in the control (21% O2) or cobalt chloride-treated group (Fig. 1C) and no significant differences in the mRNA levels of HIF-1α between the two groups (Fig. 1D). This suggests that cobalt chloride prevents the breakdown of HIF-1α protein rather than increasing the gene expression of HIF-1α in the undifferentiated hES2 cells.
Cobalt chloride treatment initiates the differentiation of undifferentiated hES2 cells
To evaluate the effects of cobalt chloride-induced HIF-1α stabilization on initiating cardiac differentiation, cultured hES2 cells were harvested immediately following cobalt chloride treatment, and the expression of early cardiac-specific transcription factors [GATA-binding protein 4 (GATA4) and NK2 transcription factor related locus 5 (Nkx2.5)] and cardiotrophic factors [vascular endothelial growth factor (VEGF) and cardiotrophin-1 (CT1)] were compared with controls using quantitative real-time PCR analysis. Cobalt chloride pretreatment significantly increased mRNA levels of Nkx2.5 (1.8-fold, p<0.05), VEGF (1.6-fold, p<.05), and CT1 (4.3-fold, p<0.005), but not GATA4 (Fig. 2A). Interestingly, in addition to the early cardiac markers, the expression of the early neuronal marker, neurogenin1 (NGN) was also increased significantly (3.1-fold, p<0.005) upon cobalt chloride treatment, whereas those governing the development of skeletal muscle (MYOG) and erythropoietin lineage (GATA1 and SPI1) did not show significant changes (Fig. 2A).
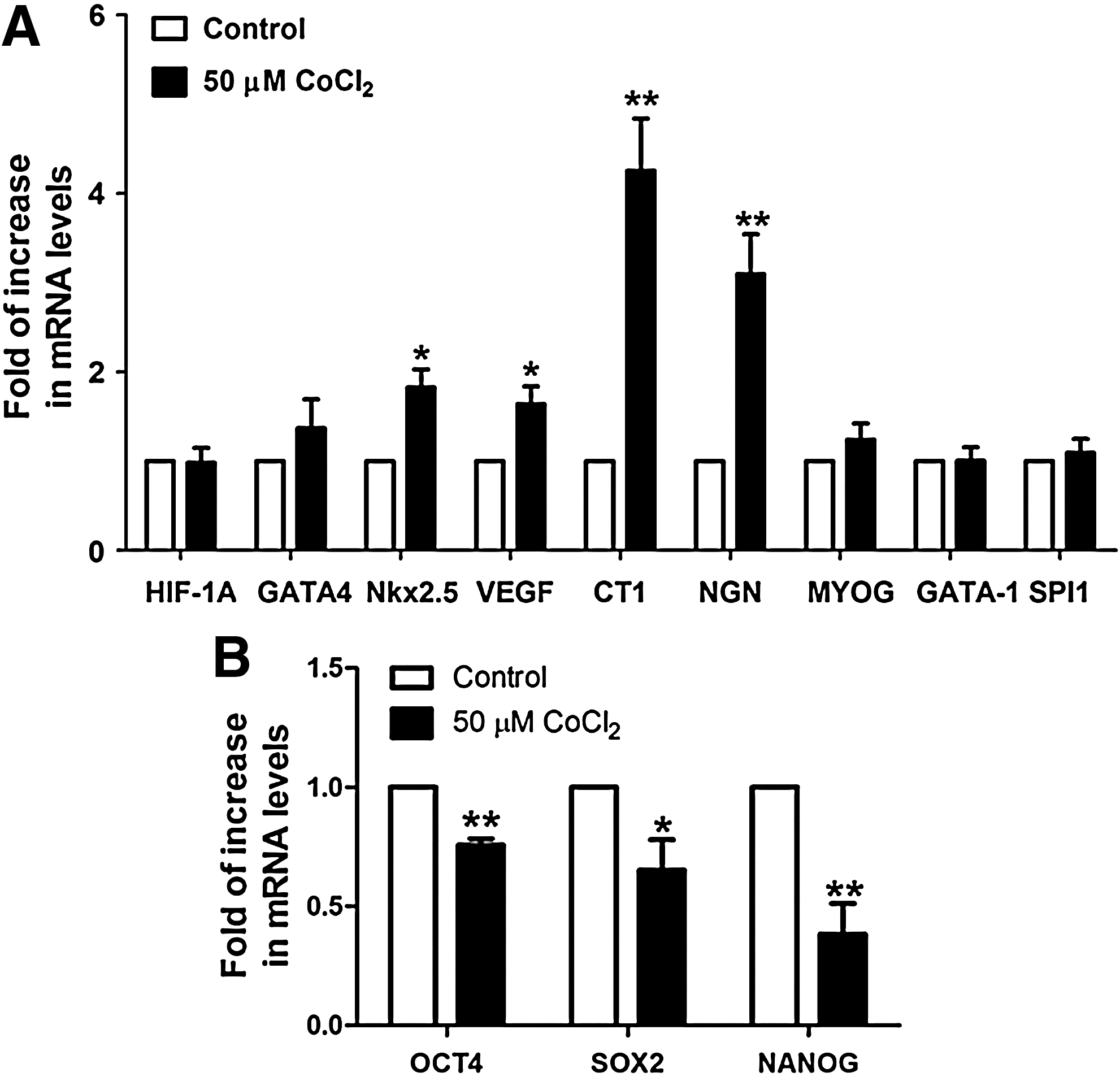
Cobalt chloride treatment increased the mRNA levels of cardiogenic transcription factors, cardiotrophic factors, and neuronal transcription factor. (
On the other hand, the expressions of the pluripotency markers, octamer-binding transcription factor 4 (OCT4), sex determining region Y-box 2 (SOX2), and homeobox protein NANOG (NANOG) were significantly reduced by 0.76-fold (p<0.005), 0.65 (p<0.05), and 0.38 (p<0.005), respectively, after cobalt chloride treatment (Fig. 2B).
Cobalt chloride pretreatment facilitate the cardiac differentiation of hES2 human ECSs
The control and cobalt chloride-pretreated cells were then allowed to undergo cardiac differentiation. Comparison at the 20th day after plating revealed that the percentage of beating embryoid bodies (EBs) (Fig. 3A; p<0.05) and sarcomeric–actinin-positive cells (Fig. 3B; p<0.005) was significantly higher following cobalt chloride treatment. The mRNA level of the cardiac specific genes, that is, myosin heavy chain (MHC)-A (1.7-fold, p<0.05); MHC-B (1.8-fold, p<0.05), and myosin light chain 2V (MLC2V) (1.5-fold, p<0.05) in the EBs differentiated from the cobalt-treated hES2 cells was also significantly higher (Fig. 3C). Immunohistochemistry demonstrated that the cardiomyocytes (cardiac troponin-T positive and sarcomeric–actinin-positive cells) differentiated from the cobalt chloride-treated group were generally more elongated (a significantly higher length/width ratio) than those from the control group (Fig. 4A and B). At higher magnification, a more organized pattern of myofibrils could be observed in the cobalt chloride-pretreated group. These results suggest that cobalt chloride pretreatment facilitates cardiac differentiation of the undifferentiated hES2 cells.
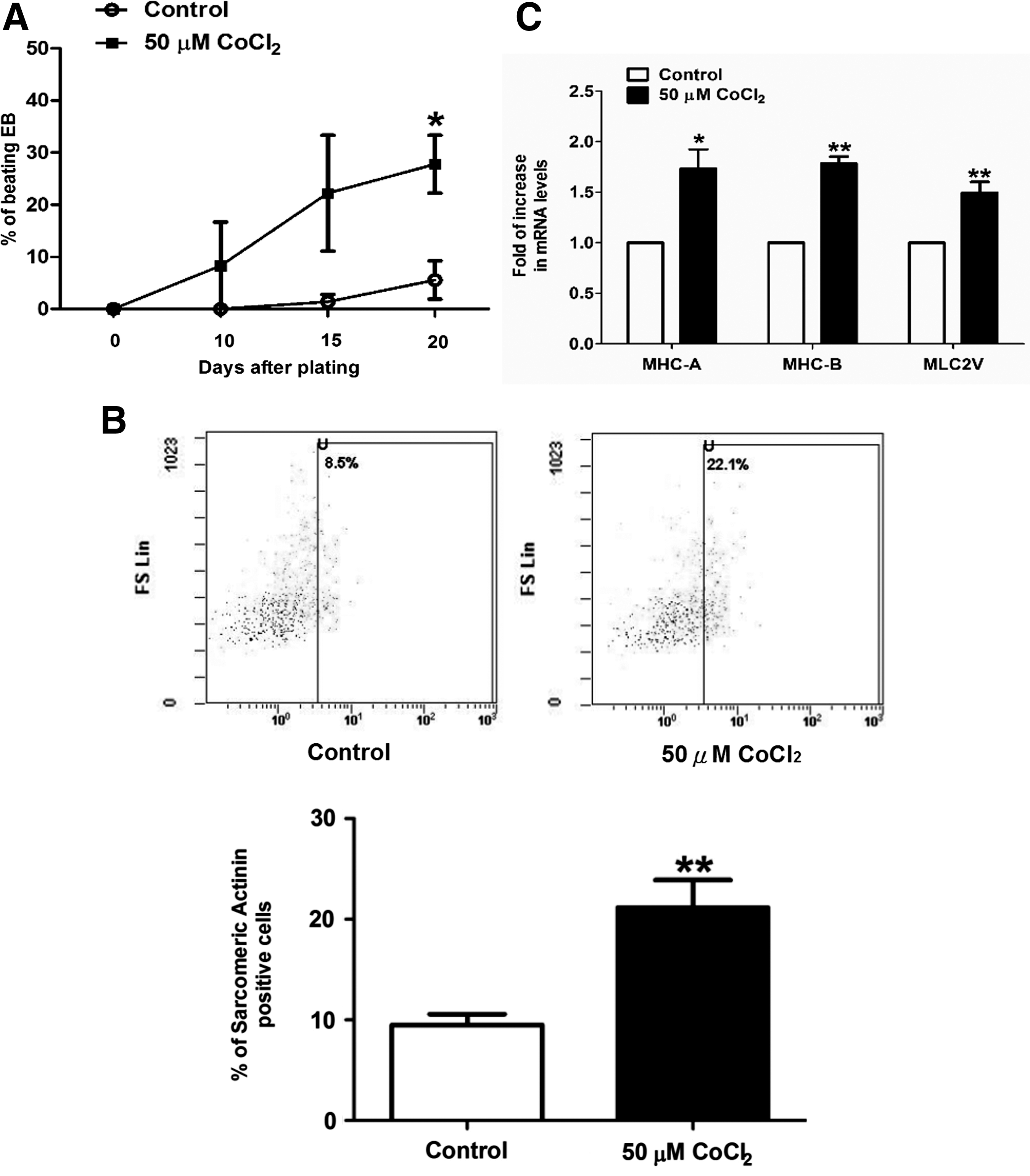
Effect of cobalt chloride pretreatment on the cardiac differentiation of hES2 human ESCs. (
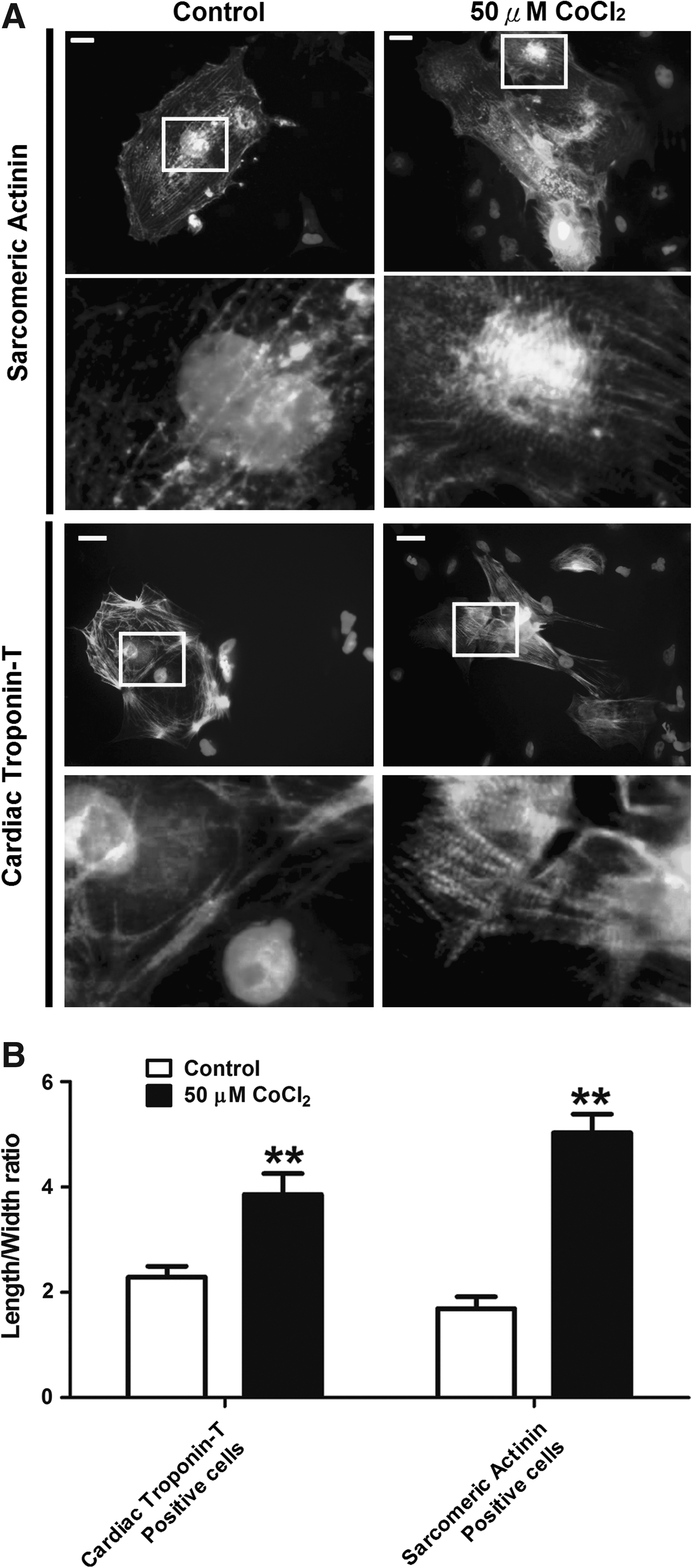
Immunohistochemistry of hES2-derived cardiomyocytes. (
Electrophysiology of hESC-derived cardiomyocytes
To test whether cobalt-induced cardiac differentiation influences the maturation of hES2-derived cardiomyocytes, their electrophysiological properties were characterized by whole-cell patch clamp experiments. Figure 5 depicts the electrophysiogical characteristics (i.e., ventricular-like, atrial-like, and nodal-like) and the corresponding representative action potential recordings of cardiomyocytes derived from the control and cobalt chloride-treated hES2. In the control group, out of seven cells analyzed, 71% showed ventricular-like phenotype and 29% exhibited the atrial-like phenotype (Fig. 5A, left panel). Despite the limited number of cells being studied, this observation is inline with the previous study (Moore et al., 2008). In the cobalt chloride-pretreated group, of 24 cardiomyocytes measured, 67%, 25%, and 8%, respectively demonstrated atrial-, ventricular-, and nodal-like phenotype (Fig. 5A, right panel). When the action potentials at 90% (APD90s) and 50% (APD50s) repolarization of both groups were compared, ventricular-like cells in particular showed a decrease in both APD90s (p<0.05) and APD50s (p<0.01) on cardiomyocytes of the cobalt chloride-pretreated group (Fig. 5B and C). In addition, maximum rise of the action potential upstrokes (dV/dtmax) for cardiomyocytes showing atrial- and ventricular-like phenotypes were both significantly increased (p<0.05) (Fig. 5D). On the other hand, the maximum diastolic potentials (MDPs) of both atrial-like and ventricular-like cells showed no significant difference between the two groups (Fig. 5E).
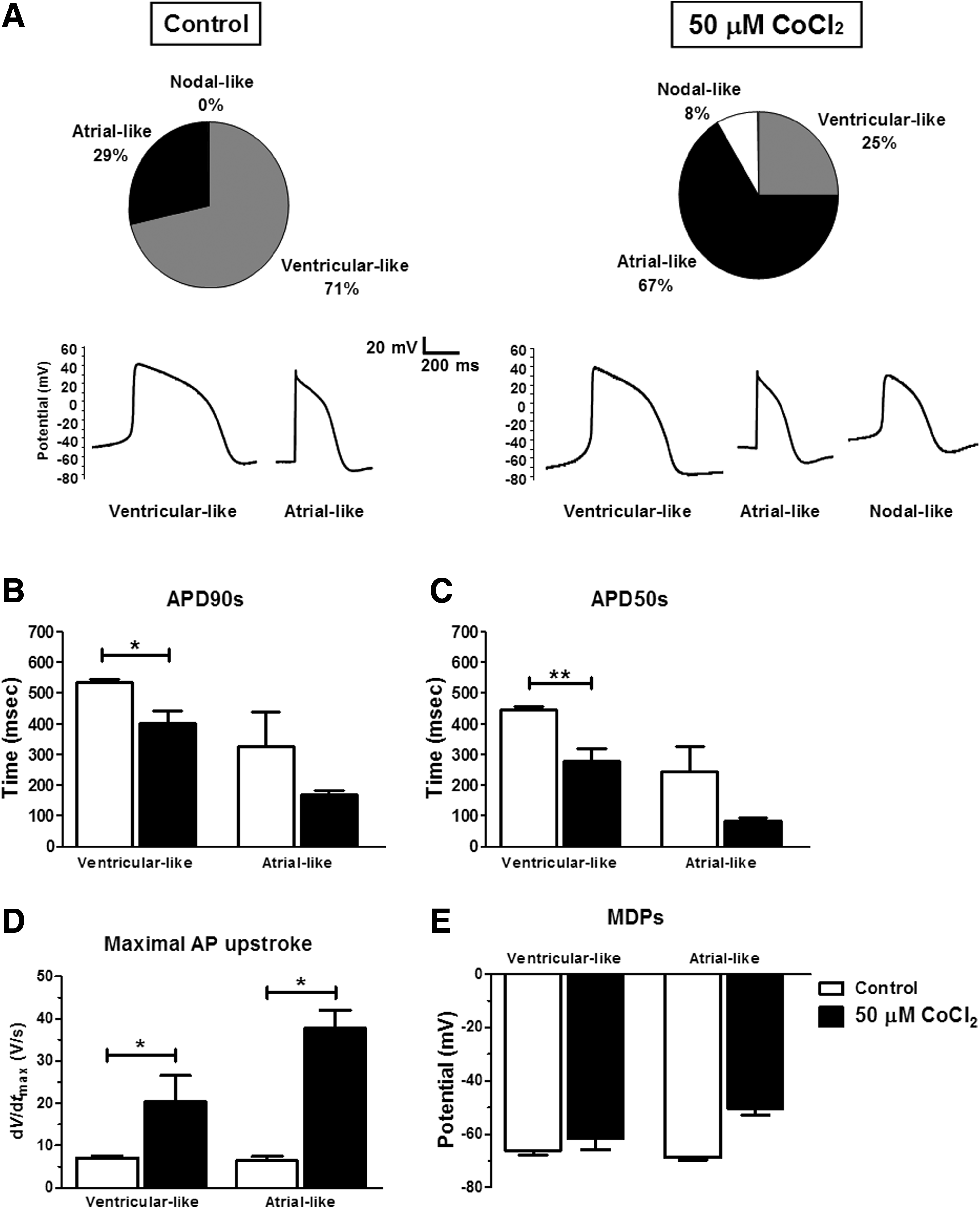
(
Cardiomyocytes differentiated from cobalt chloride-pretreated cells exhibit improved calcium-handling ability
Spontaneous intracellular calcium (Ca2+) transients in the hES2-derived cardiomyocytes were assessed using confocal laser microscopy 25 to 30 days following onset of beating. In both control and cobalt chloride-pretreated groups, Ca2+ flux was observed in their derived cardiomyocytes. Fluorescence intensity values measured against time revealed that the amplitude of Ca2+ transient measured in cardiomyocytes derived from the cobalt chloride-pretreated hES2 cells was approximately 2.5-fold higher than that in the untreated hES2-derived cardiomyocytes (Fig. 6A and B; p<0.05). Similarly, kinetic analysis revealed that cardiomyocytes derived from cobalt chloride-pretreated hES2 cells displayed faster calcium release and reuptake rates as indicated by the significant increase in the upstroke (Vmaxupstroke) and decay (Vmaxdecay) velocities (Fig. 6C and D; p<0.05).
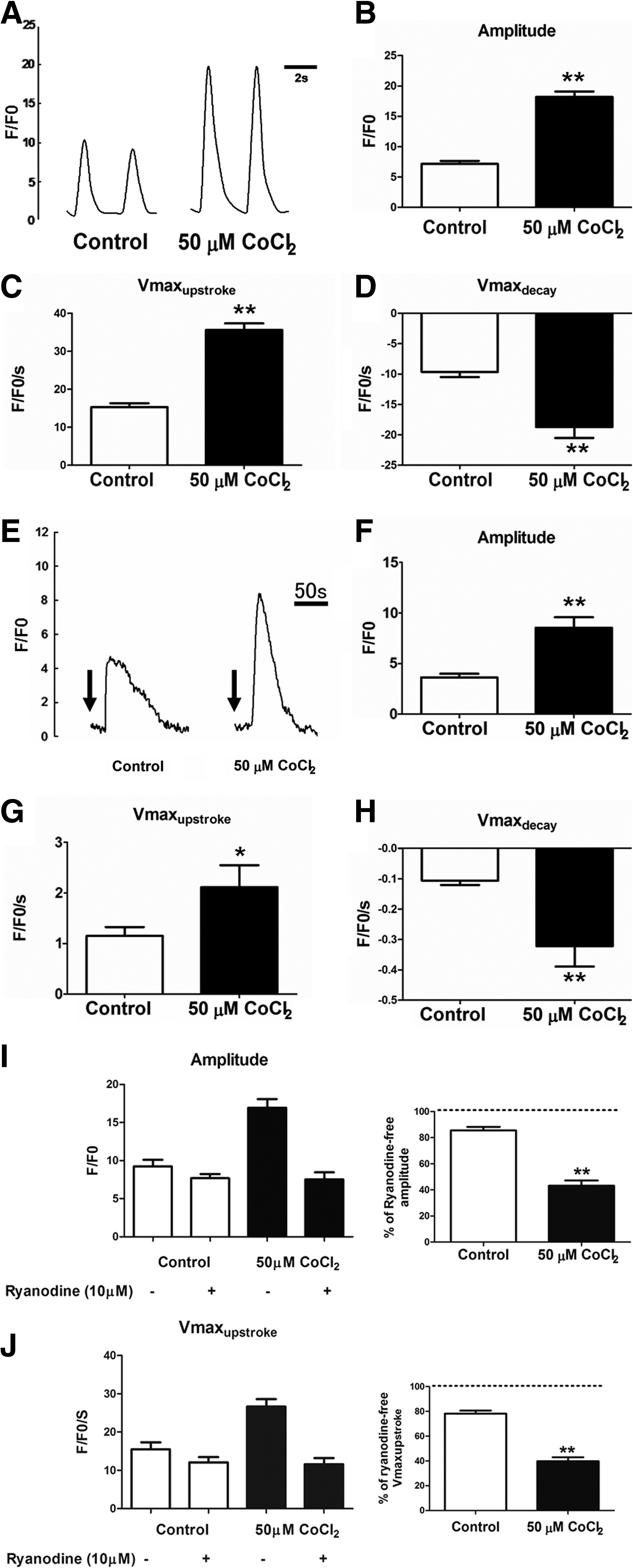
(
To evaluate the sarcoplasmic reticulum performance, the caffeine-induced Ca2+ transient was also recorded. As shown in Figure 6E, brief exposure to 10 mM caffeine induced a surge in cytosolic Ca2+ that subsequently decayed back to baseline in the cardiomyocytes derived from both control and cobalt chloride-pretreated hES2 cells. Similar to the observations in spontaneous Ca2+ transient, the amplitude of the caffeine-induced Ca2+ from the cobalt chloride-pretreated group was significantly higher compared with the control group (Fig. 6F, approximately 2.4-fold increase; p<0.01). This finding suggests that sarcoplasmic reticulum Ca2+ storage is increased in cardiomyocytes derived from cobalt chloride-pretreated hES2 cells. In the kinetic analysis, cardiomyocytes derived from cobalt chloride-pretreated hES2 cells also displayed faster upstroke and decay velocities during caffeine-induced Ca2+ transient (Fig. 6G and H). In addition, application of 10 μM ryanodine significantly reduced the amount and release rate of intracellular calcium in these cardiomyocytes, but not those from the control group (Fig. 6I and J).
In cardiomyocytes, the sodium calcium exchanger (NCX1), cardiac sarcoplasmic/endoplasmic reticulum calcium ATPase (SERCA2A), and cardiac specific ryanodine receptor (RYR2) are the key components that mediate calcium transport (Graham et al., 2004). Calsequestrin (CaSQ), triadin, and junctin play a role in regulating the sarcoplasmic reticulum function via their binding to RYR2 (Bers, 2004; Fan et al., 2008; Knollmann, 2009). Real-time quantitative PCR analysis revealed that the expression of SERCA2A (3.6-fold, p<0.05), RYR2 (4.1-fold, p<0.05), and triadin (2.5-fold, p<0.05) was significantly increased in EBs derived from the cobalt chloride-pretreated hES2 cells compared with control cells (Fig. 7). Nevertheless, there was no significant change in the mRNA level of NCX1, CaSQ, or junctin in the two groups. These findings suggest that improved sarcoplasmic reticulum function of cardiomyocytes derived from cobalt chloride-pretreated hES2 cells is likely related to increased expression of the Ca2+-handling components.
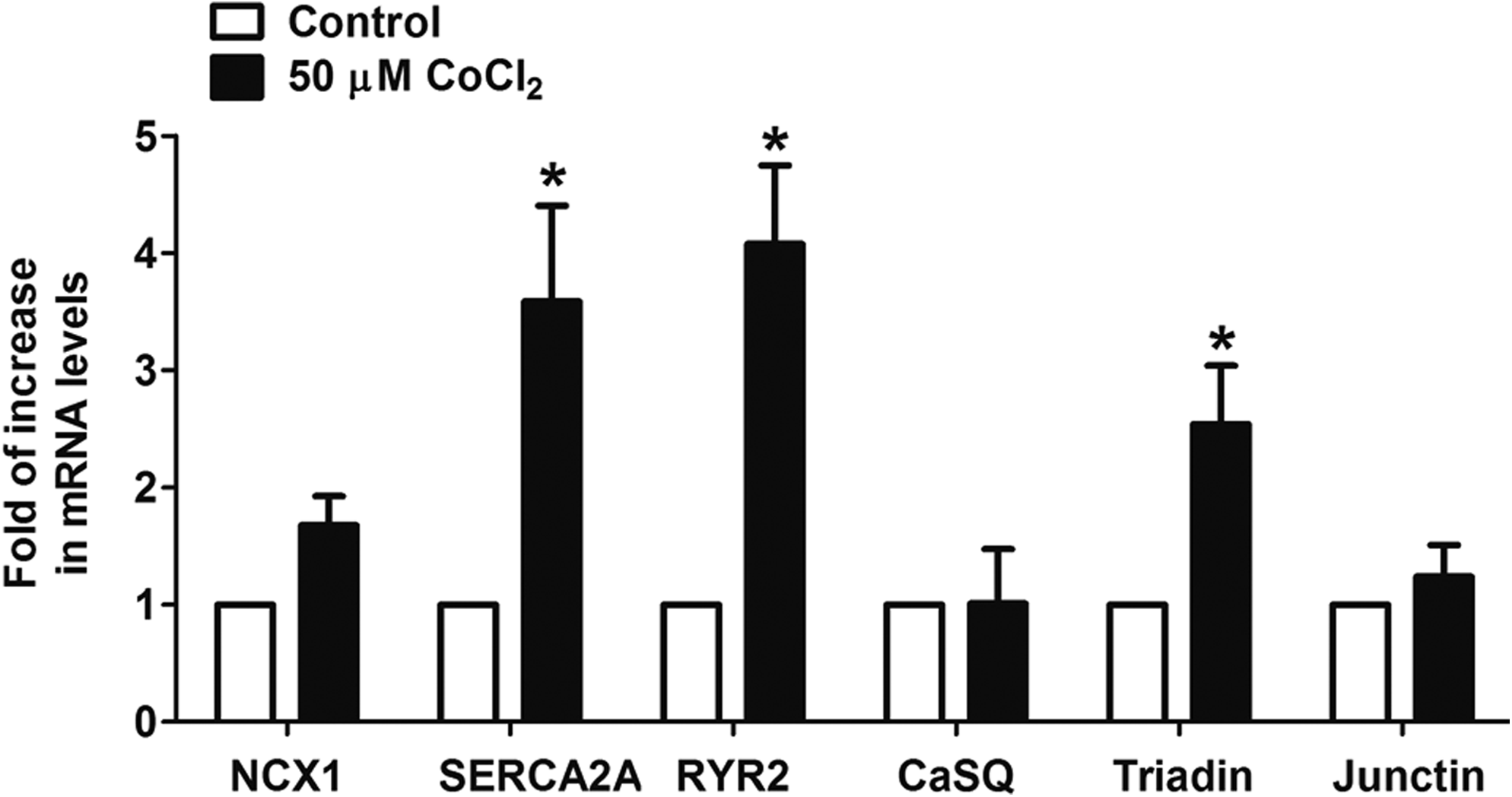
The expression of calcium-handling components in cardiomyocytes. The mRNA levels of calcium-handling components NCX1, SERCA2A, RYR2, CaSQ, Triadin, and Junctin in the EBs derived from the control or hES2 cells pretreated with 50 μM cobalt chloride (CoCl2) were evaluated by real-time quantitative PCR analysis using GATA4 as an internal reference. Data shown as mean±SEM from at least three independent experiments; *P<0.05.
Discussion
Our recent study has demonstrated that the cardiac differentiation and maturation of murine ESCs can be promoted by exogenous expression of the full-length HIF-1α cDNA under a constitutive EF1α promoter (Ng et al., 2010). These observations highlight the potential importance of the HIF-1α subunit in directing cultured ESCs to differentiate into more mature cardiomyocytes. Nevertheless, the therapeutic application of this strategy is limited by the requirement of a viral vector. The present study aimed to evaluate the use of cobalt chloride-induced HIF-1α stabilization to promote cardiac differentiation of cultured human ESCs. The use of divalent ions (such as cobalt, iron, or nickel compounds) has a potential advantage over the reduced oxygen tensions required in stabilizing the HIF-1α subunit. First, the substantial cost involved in the setup and operation of a reduced oxygen culture system is avoided. Second, the use of a pharmacological treatment that includes the defined concentration of drugs or chemicals is also anticipated to provide more robust and reproducible results between different laboratories. This is especially important given that the actual partial pressure of oxygen that cells experience may differ widely among different culture setups and sources of gas. As shown in this study, treatment of undifferentiated hES2 human ESCs with cobalt chloride for 3 days was associated with remarkably increased protein levels of HIF-1α. Although increased HIF-1α expression was also observed in cells cultured under reduced oxygen conditions, a decrease in cell viability was also evident in this study. In contrast, previous studies showed that reduced low oxygen tension can enhance the culture and maintenance of H1 human ESCs in undifferentiated states (Ezashi et al., 2005). The reasons for this discrepancy remain unclear, but may be related to different ESC lines and culture systems. In this study, the HES2 human ESC line was cultured in a feeder-free serum-free culture system. Previous studies (Ezashi et al., 2005) have used a mouse embryonic fibroblast as feeder layer supplemented with 20% KnockOut serum replacer to culture H1 human ESC line.
In this study, cobalt chloride pretreatment notably increased the occurrence of beating EBs as well as expression of cardiac specific structure components (MHC-A, MHC-B, and MLC2V) over the 25-day period of cardiac differentiation compared with controls. As cardiac differentiation of ESCs was performed in the absence of cobalt chloride, it is reasonable to speculate that cobalt chloride help initiate the early cardiac differentiation process before embryoid body formation, and this could be evidenced by the downregulation of the pluripotent markers (OCT4, SOX2, and NANOG), and the upregulation of early cardiac differentiation factors Nkx2.5, VEGF, and notably CT1 gene expression during the pretreatment period. It is interesting that the expression of NGN, an early neuron-specific marker, was also significantly increased after cobalt chloride treatment, while the expressions of the skeletal muscle specific (MYOG) and hematopoietic markers (GATA1 and SPI1) shows no significant difference. The reasons account for such observation is not clear but deserve further investigation that would be out of the scope of the present study.
HIF-1α plays an important role in the embryonic heart development (Iyer et al., 1998; Nagao et al., 2008; Ramirez-Bergeron and Simon, 2001), yet the direct action of HIF-1α on the promoters of many cardiac genes remains to be determined. Recently, Nagao et al. (2008) described that the knockdown of HIF-1α causes significant reduction in the nkx2.5 expression in a Xenopus model (Nagao et al., 2008) and this may explain, in part, our observations that cobalt chloride increase NKX2.5 expression in the undifferentiated hES2 cells.
The importance of VEGF and CT1 in the mediation of cardiac differentiation from pluripotent stem cells has been highlighted in various studies (Ateghang et al., 2006; Chen et al., 2006; Xinyun et al., 2010). The appearance of a population of kinase insert domain protein receptor (KDR)-positive progenitor cells during the very initial period of differentiation dictates the subsequent cardiac differentiation events of the ESCs (Yang et al., 2008). Accordingly, increased expression of VEGF during cobalt chloride pretreatment may play a role in activation of the KDR-positive cardiac progenitor cells to increase the occurrence of beating EBs and the percentage of sarcoplasmic actinin-positive cardiomyocytes. It should be noted that attempts have been made to supplement cobalt chloride throughout the whole differentiation period, but such an approach appears to reduce the number of beating EBs (data not shown). The underlying reason is unclear but may be related to the oxidative toxicity that arises from prolonged cobalt chloride treatment.
When the cobalt chloride-pretreated groups was compared with the control group, the former showed a notable increase not only in the differentiation of atrial- and nodal-like cardiomyocytes, but also the maximum rise of the action potential upstroke (dV/dtmax). Such a significant increase in the dV/dtmax reflects an improved electrical maturity after cobalt chloride treatment. Nonetheless, the limited sample size means further work is required to identify whether cobalt chloride pretreatment determines the fate of cadiomyogenesis. In addition to these electrophysiological observations, calcium homeostasis of the hES2-derived cardiomyocytes was also studied. Those derived from the cobalt chloride-pretreated group exhibited significant increases in the amplitude, upstroke rate, and decay rates of the calcium transient compared with the control group. This indicates that the former may more effectively handle the release and reuptake of intracellular calcium during excitation-contraction coupling. In addition, the increased performance of caffeine-induced calcium transient and increased ryanodine sensitivity in the cobalt chloride-pretreated cells suggests these cardiomyocytes have improved sarcoplasmic reticulum function. This assumption was further supported by the observation that both SERCA2A and RYR2, the two key components that mediate the sequestration and release of calcium from the sarcoplasmic reticulum, respectively, were significantly upregulated in the cobalt chloride-pretreated cells. In addition to SERCA2A and RYR2, the expression of RYR2 regulatory components dictates the maturity of the ESC-derived cardiomyocytes (Liu et al., 2007, 2009). Although there was no observable change in the expression of CaSQ and junctin between the two groups, a significant increase in the mRNA levels of triadin was also found in the cobalt chloride-pretreated group: this might contribute to the improved sarcoplasmic reticulum-mediated calcium transient in the treated cells. With increasing evidence of the importance of sarcoplasmic reticulum calcium load and transient in contractile function of ESC-derived cardiomyocytes (Fu et al., 2006; Liu et al., 2007), it has become clear that the proper calcium-handling property of the ESC-derived cardiomyocytes is crucial to functional integration of the transplanted cells into the host tissue (Liu et al., 2007).
In conclusion, our present study provides evidence that cobalt chloride-induced HIF-1α stabilization facilitates the cardiac differentiation of human ESCs in a serum- and feeder-free system. Unlike low oxygen culture systems, no sophisticated or costly facilities are required, and there is no reduction in viability of cobalt chloride-treated human ESCs. Cardiomyocytes derived from cobalt chloride-treated cells also appear to have a more advanced calcium-handling ability. This may be due to increased expression of the sarcoplasmic reticulum calcium-handling components. These data provide a novel strategy to encourage maturation of calcium handling of cardiomyocytes from ESCs for therapeutic application.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China/Research Grants Council Joint Research Scheme (HKU 722/05 to M.L.F. and H.F.T.), the HKU Small Project Funding (201007176133 to K.M.N., C.W.S., and H.F.T.); Collaborative Research Fund of Hong Kong Research Grant Council (HKU 8/CRF/09 to C.W.S and H.F.T)
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.