
Abstract
Abstract
A culture system capable of sustaining self-renewal of buffalo embryonic stem (ES) cell-like cells in an undifferentiated state over a long period of time was developed. Inner cell masses were seeded on KO-DMEM + 15% KO-serum replacer on buffalo fetal fibroblast feeder layer. Supplementation of culture medium with 5 ng/mL FGF-2 and 1000 IU/mL mLIF gave the highest (p<0.05) rate of primary colony formation. The ES cell-like cells' colony survival rate and increase in colony size were highest (p<0.05) following supplementation with FGF-2 and LIF compared to other groups examined. FGF-2 supplementation affected the quantitative expression of NANOG, SOX-2, ACTIVIN A, BMP 4, and TGFβ1, but not OCT4 and GREMLIN. Supplementation with SU5402, an FGFR inhibitor (≥20 μM) increased (p<0.05) the percentage of colonies that differentiated. FGFR1-3 and ERK1, K-RAS, E-RAS, and SHP-2, key signaling intermediates of FGF signaling, were detected in ES cell-like cells. Under culture conditions described, three ES cell lines were derived that, to date, have been maintained for 135, 95, and 85 passages for over 27, 19, and 17 months, respectively, whereas under other conditions examined, ES cell-like cells did not survive beyond passage 10. The ES cell-like cells were regularly monitored for expression of pluripotency markers and their potency to form embryoid bodies.
Introduction
FGF-2 plays an important role in the survival and maintenance of human ES cells in a pluripotency state (Amit et al., 2000). With high concentrations of FGF-2, human ES cells can be maintained in feeder-free conditions in combination with Activin-A or by suppression of BMP signaling (Vallier et al., 2005; Xiao et al., 2006; Xu et al., 2005). TGFβ-1/Activin/Nodal signaling has been observed to be present in human and rabbit ES cells (Honda et al., 2009; Vallier et al., 2005). Ludwig et al. (2006) developed a synthetic medium mTeSR1 for xenofree and feeder-free culture of human ES cells using high concentrations of these growth factors. Like TGFβ-1/Activin/Nodal signaling, Wnt signaling is also one of the conserved signaling pathways across different species (Dravid et al., 2005; Miyabayashi et al., 2007).
True pluripotent, germ-line competent ES cell lines are available for mice, chicken, and recently from the rat (Li et al., 2008). Attempts to derive germ-line competent ES cells from other mammalian species (apart from human) have met with failure. In case of farm animals, extensive attempts have been made to derive ES cells from bovine and porcine species. Although ES cell-like cells from farm animals are routinely cultured with LIF and FGF-2 or both (Munoz et al., 2008), to our knowledge there are no detailed studies on whether these growth factors are an absolute requirement for their survival. Also, the signaling pathways responsible for maintaining the self-renewal and pluripotency of ES cells in farm animal species, is not known. Although bovine ES cells have been cultured in the presence of either LIF or FGF-2 or both (Gong et al., 2010; Roach et al., 2006), neither FGF-2 nor LIF was reported to be required for their survival over a long-term culture of nearly 3 years (Mitalipova et al., 2001). Recently, Pant and Keefer (2009) used FGF-2, Noggin, Activin-A, and BMP-4 supplementation for bovine ICM culture and found that except for Noggin, no growth factor could sustain expression of OCT4, NANOG, and SOX-2.
Buffalo (Bubalus bubalis) is perhaps the most important farm animal in many developing nations of Asia in view of its immense contribution to milk, meat, and draft. The availability of ES cells from buffalo could help bring about its genetic modification through efficient gene transfer by providing an abundance of stem cells. Also, because the availability of human embryos and ES cells is restricted due to legal and ethical issues, the availability of ES cells from farm animals would permit the extension of studies in murine cells. Derivation and maintenance of buffalo ES cell-like cells from parthenogenetically produced buffalo blastocysts (Sritanaudomchai et al., 2007) or IVF blastocysts has been reported from our (Anand et al., 2009; Kumar et al., 2011; Verma et al., 2007) and other (Huang et al., 2010) laboratories. However, in none of these studies could the ES cell-like cells survive beyond passage 10. The aim of present study was to explore the possibility of developing a culture system capable of sustaining the self-renewal of buffalo ES cell-like cells in an undifferentiated state, over a long period of time. Parts of the results of this study have been published earlier as an abstract (Sharma et al., 2011).
Materials and Methods
All chemicals and media were from Sigma Chemical Co. (St. Louis, MO) and disposable plastic wares from Nunc (Roskilde, Denmark) unless otherwise mentioned.
In vitro embryo production
Cumulus–oocyte complexes (COCs) collected from abattoir-derived buffalo ovaries were subjected to in vitro maturation (IVM) and in vitro fertilization (IVF), as described earlier (Chauhan et al., 1998) with some modifications. Briefly, the COCs were washed several times with the IVM medium [M-199+10% fetal bovine serum (FBS) + 5 μg/mL pFSH + 1 μg/mL estradiol-17β + 0.81 mM sodium pyruvate + 10% buffalo follicular fluid + 50 μg/mL gentamicin sulphate], and groups of 15 to 20 COCs were placed in 100 μL droplets of the IVM medium, overlaid with sterile mineral oil in 35-mm Petri dishes, and were cultured in a CO2 incubator (5% CO2 in air) at 38.5°C for 24 h. For IVF, two straws of frozen-thawed buffalo semen, which had been tested for IVF earlier, were washed twice with the washing Bracket and Oliphant (BO) medium, containing 10 μg/mL heparin, 137.0 μg/mL sodium pyruvate, and 1.942 mg/mL caffeine sodium benzoate. The pellet was resuspended in 0.5 mL of the capacitation and fertilization BO medium (washing BO medium containing 10 mg/mL fatty acid-free BSA). The in vitro matured oocytes were washed twice with the fertilization BO medium and were then transferred to 50 μL droplets (15–20 oocytes/droplet) of the capacitation and fertilization BO medium. The spermatozoa in 50 μL of the capacitation and fertilization BO medium (2–4 million spermatozoa/mL) were then added to the droplets containing the oocytes, covered with sterile mineral oil, and were placed in a CO2 incubator (5% CO2 in air) at 38.5°C for 18 h for IVF. After this, the cumulus cells were washed off the presumed zygotes by gentle pipetting. The presumed zygotes were washed several times with modified Charles Rosenkrans medium with amino acids (mCR2aa) containing 0.6% bovine serum albumin (BSA) and were cultured in this medium for 48 h postinsemination. The embryos were then shifted to the IVC medium (mCR2aa + 0.6% BSA + 10% FBS) and were cultured in 100 μL droplets of this medium on original beds of granulosa cells for up to 8 days postinsemination in a humidified CO2 incubator (5% CO2 in air) at 38.5°C. The medium was replaced with 50% of fresh IVC medium every 48 h.
Derivation, maintenance, and characterization of ES cell-like cells
Inner cell masses (ICMs) were isolated mechanically from in vitro produced day 8 hatched blastocysts using Microblades™ (MicroBlades, MTB-05; Micromanipulator Microscope Company, Inc., Washington, DC) under a zoom stereomicroscope and were cultured on mitomycin C (10 μg/mL) inactivated buffalo fetal fibroblast feeder layers in ES cell medium (SCM), which is comprised of Knockout DMEM™ (Invitrogen Corporation, Carlsbad, CA) + 15% Knockout serum replacement™ (Invitrogen) + 2 mM L-glutamine + 50 μg/mL gentamicin sulphate + 1% MEM nonessential amino acids + 0.1 mM β-mercaptoethanol. The SCM was supplemented with 1000 IU/mL mLIF and/or FGF2 at different concentrations (0, 5, 10, 20, and 40 ng/mL) for initial derivation of primary colonies and standardization of optimal FGF2 concentration. The colonies were mechanically passaged every 5 to 6 days in a split ratio of 1:2 or 1:3 depending upon the colony size. ES cell-like cells were regularly checked for their pluripotency by studying the expression of transcription-based markers like OCT4, NANOG, and SOX-2 and/or, surface based-markers (SSEA-1, SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81). The experiments related to effect of FGF-2 on colony survival, proliferation, and expression of pluripotency markers of buffalo embryonic stem cell-like cells were carried out both at lower (15–40) and higher passages (60–90); however, the data is presented from one of the passages only as the observations were similar among both the passages.
The transcription-based markers of pluripotency were examined by reverse transcriptase (RT)-PCR for which, total RNA was isolated from ES cell-like cells using Tri® Reagent. The quality and integrity of RNA was checked by Nanoquant (Teccan, Salzburg, Austria), and agarose gel electrophoresis. The RNA was reverse transcribed by Revertaid first-strand cDNA synthesis kit (K1621, Fermentas Life Sciences, Hanover, MD), as per manufacturer's protocol. The annealing temperature and PCR conditions of the target genes are given in Supplementary Table 1 (Supplementary Data are available online at www.liebertonline.com/cell). For immunofluorescence staining, the ES cell-like cells' colonies were fixed in 4% paraformaldehyde in DPBS for 30 min, washed three times with DPBS, and then permeabilized by 0.1% Triton X-100 in DPBS for 30 min. After thorough washing with DPBS, the colonies were incubated with the blocking solution (4% normal goat serum) for 30 min, and then with the primary antibody at a dilution of 1:10 to 1:20 for 1 h. The addition of primary antibody was omitted in the respective controls. After washing three times with DPBS, the ES cell-like cells' colonies were incubated with the appropriate FITC-labeled secondary antibody (goat antirat IgM or goat antirabbit IgG or antimouse IgG or IgM, diluted 1:100 to 1:200) for 2 h. The colonies were then examined under a fluorescence microscope (Diaphot, Nikon, Tokyo, Japan). The primary antibodies for SSEA-1, SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81 were purchased from Chemicon® International and the FITC-conjugated secondary antibodies were from Sigma Chemical Co. Alkaline phosphatase staining of ES cell-like cells' colonies was performed using a kit (Catalog No. 86C, Sigma Chemical Co.) according to the manufacturer's protocol. The differentiation potential of the ES cell-like cells was examined by generation of embryoid bodies via the hanging drop method followed by spontaneous differentiation and also by detecting the expression of the markers of the three germ layers in embryoid bodies as described earlier (George et al., 2011).
Quantitative real-time PCR (qRT-PCR), wherever carried out, was performed on CFX 96 I Cycler (Biorad, Hercules, CA) in 10-μL reaction volume containing 5 μL of SYBR Green master-mix (Maxima SYBR Green Mastermix, Fermentas), 0.5 μM of each primer, and 5×diluted c-DNA. Thermal cycling conditions consisted of initial denaturation at 95°C for 5 min, followed by 40 cycles of 10 sec at 95°C, 15 sec at the corresponding annealing temperature (Supplementary Table 1), and 15 sec at 72°C followed by 95°C for 10 sec and melt curve. All primer pairs used were confirmed for their PCR efficiency, and specific products were checked by melt curve analysis and for the appropriateness of size by agarose gel electrophoresis. Primer sequences are provided in the supplementary data (Supplementary Table 1). The expression data were normalized to the expression of GAPDH and were analyzed with CFX Manager software (BioRad). Relative mRNA levels were presented as percentage of calibrator used in the each experiment. In all experiments, three trials were carried out, each with three replicates. Wherever required, statistical analysis was carried out using ANOVA, and the level of significance was kept at p≤0.05.
Experimental design
Experiment 1 was aimed at examining the effect of supplementation of SCM with FGF-2 on the primary colony formation rate to determine the optimal FGF-2 concentration. ICMs were divided into four groups and were seeded on feeder layer in SCM in the presence of 0, 5, 10, 20, or 40 ng/mL FGF-2. The concentration that gave the maximum percentage of colony survival was chosen for the rest of the experiments.
Because in the first experiment FGF-2 was added with LIF, the FGF-2 role alone could not be determined. Hence, the objective of Experiment 2 was to investigate the effects of supplementation of SCM with FGF-2 (5 ng/mL), alone or in combination with LIF on primary colony formation rate. ICMs were divided into the following four groups for seeding on feeder layer in SCM. Group 1: SCM alone; Group 2: SCM + 1000 IU/mL LIF; Group 3: SCM + FGF-2; and Group 4: SCM + 1000 IU/mL LIF + FGF-2.
In Experiment 3, in which the effects of supplementation of SCM with FGF-2 (5 ng/mL), alone or in combination with LIF on colony survival rate and self-renewal were examined, the ES cell-like cells' colonies were divided into the following four groups. Group 1: SCM alone; Group 2: SCM + 1000 IU/mL LIF; Group 3: SCM + FGF-2; and Group 4: SCM + 1000 IU/mL LIF + FGF-2. For determining the colony survival rate, the colony morphology was examined after 6 days of culture in the respective medium. The colonies were considered to be normal when they were observed to be rounded or dome shaped, compact, with defined edges, and with no apparent signs of cell death or differentiation (Supplementary Fig. 1A). The ES cell like cells' colonies were considered to be differentiated when they were found to be flattened fragmented, with loosened and scattered cells with apparent signs of cell death or differentiation (Supplementary Fig. 1B–F). The colonies in different groups showed different types of proliferation and differentiation patterns. The self renewal capacity of colonies was determined by the maximum number of passages for which the colonies could survive in culture without dying or getting differentiated.
Experiment 4 was aimed at examining the effects of supplementation of SCM with FGF-2 (5 ng/mL), alone or in combination with LIF, on quantitative expression of some transcription markers of pluripotency, that is, OCT4, NANOG, and SOX-2 in ES cell-like cells. ES cell-like cells' colonies were divided into the following four groups, that is, Group 1: SCM alone; Group 2: SCM + 1000 IU/mL LIF; Group 3: SCM + FGF-2; and Group 4: SCM + 1000 IU/mL LIF + FGF-2.
The aim of Experiment 5 was to examine the effects of supplementation of SCM with FGF-2 (5 ng/mL), alone or in combination with LIF, on the colony size. ES cell-like cells' colonies (n=25 each) were divided into four groups; that is, Group 1: SCM alone; Group 2: SCM + 1000 IU/mL LIF; Group 3: SCM + FGF-2; and Group 4: SCM + 1000 IU/mL LIF + FGF-2. ES cell colonies were cultured for 24 h to enable them to get attached to the feeder layer following which their size was recorded at 0 h (day 0), 72 h (day 3), and 144 h (day 6) of culture.
Experiment 6 was aimed at examining the effects of SU5402, an FGFR inhibitor on colony morphology and differentiation. ES cell colonies were divided into the following supplementation groups and were cultured for 6 days, Group C: SCM + LIF + FGF-2; Group F: SCM + FGF-2; Group SU10: SCM + LIF + SU5402 (10 μM); Group SU20: SCM + LIF + SU5402 (20 μM); Group SU30: SCM + LIF + SU5402 (30 μM). The percentage of colonies that were found to be differentiated as indicated by a change in their morphology, that is, those that either became flattened or turned into embryoid body-like structure, was recorded.
The objective of Experiment 7 was to examine the effects of FGF and LIF on the quantitative expression of growth factors such as Activin-A, BMP-4, TGF-β1, and Gremlin. ES cell colonies were cultured for 6 days in the presence of FGF and/or LIF following which the expression of these growth factors was determined by qRT-PCR.
Results
In the present study, buffalo ES cell-like cells were isolated and maintained for an extended period of time under in vitro conditions. The ES cell-like cells expressed OCT4, NANOG, SOX-2, REX-1, FOX-D3, NUCLEOSTAMIN, and STAT-3 (Fig. 1A) as examined by RT-PCR. The colonies also exhibited high alkaline phosphatase activity (Fig. 1B) and were found to be positive for SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81, but not for SSEA-1 as studied by immunofluorescence staining (Fig. 1C–J). Also, the presence of OCT4 was detected in the central core of ICM derived cells and in the trophectodermal layer by immunofluorescence staining (Supplementary Fig. 2). OCT4 expression could also be detected by RT-PCR in both ICM and trophectoderm (data not shown). The karyotype of the cells, which was monitored regularly, was found to be normal (data not shown). The embryoid bodies formed through the hanging drop method exhibited the presence of three germ layers (Fig. 2A and B) and different cell types were observed when embryoid bodies (Fig. 2C) were subjected to spontaneous differentiation (Fig. 2D–H).
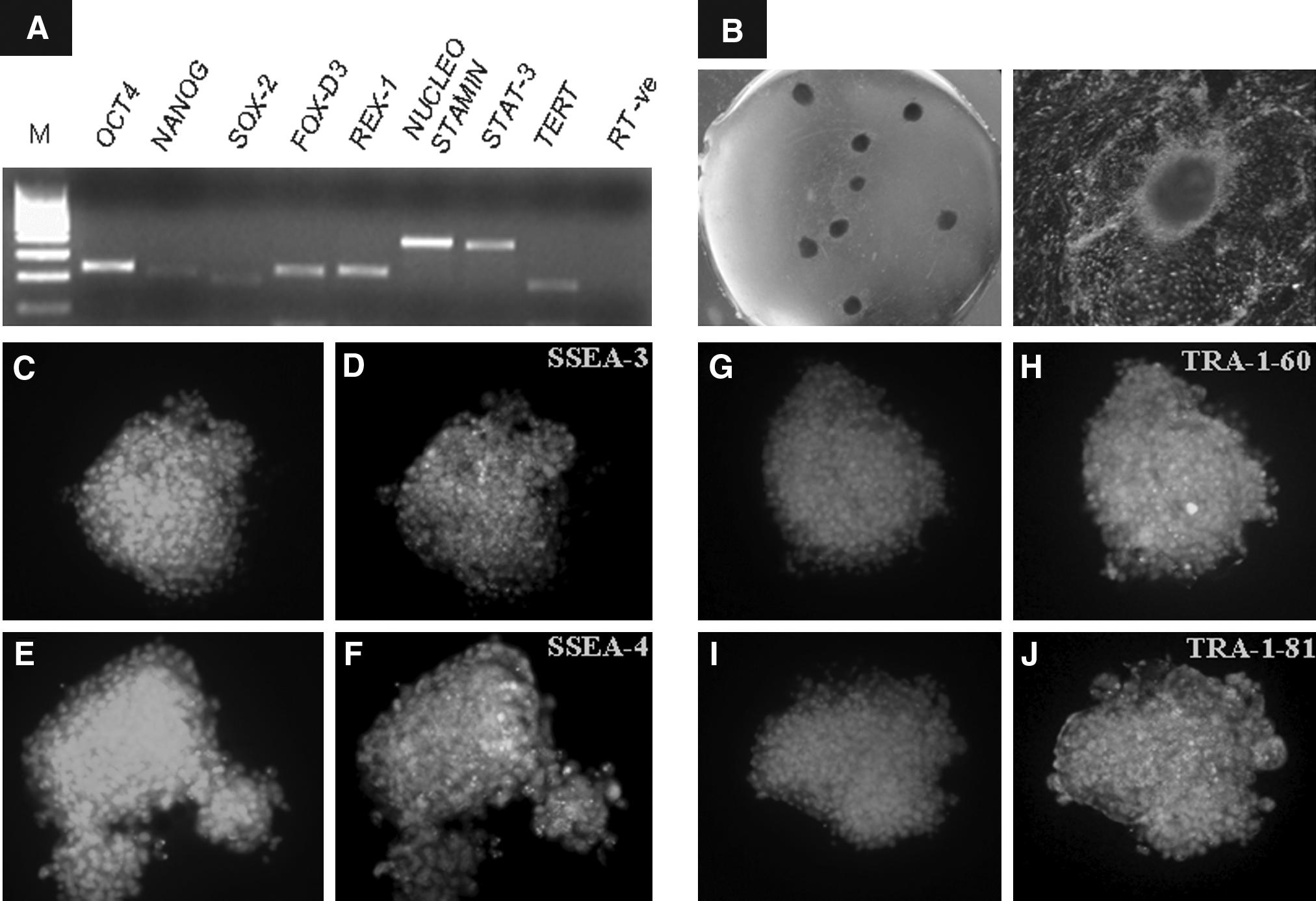
Expression of transcription-based markers OCT4, NANOG, SOX-2, REX-1, FOX-D3, NUCLEOSTAMIN, and STAT-3 examined by RT-PCR (
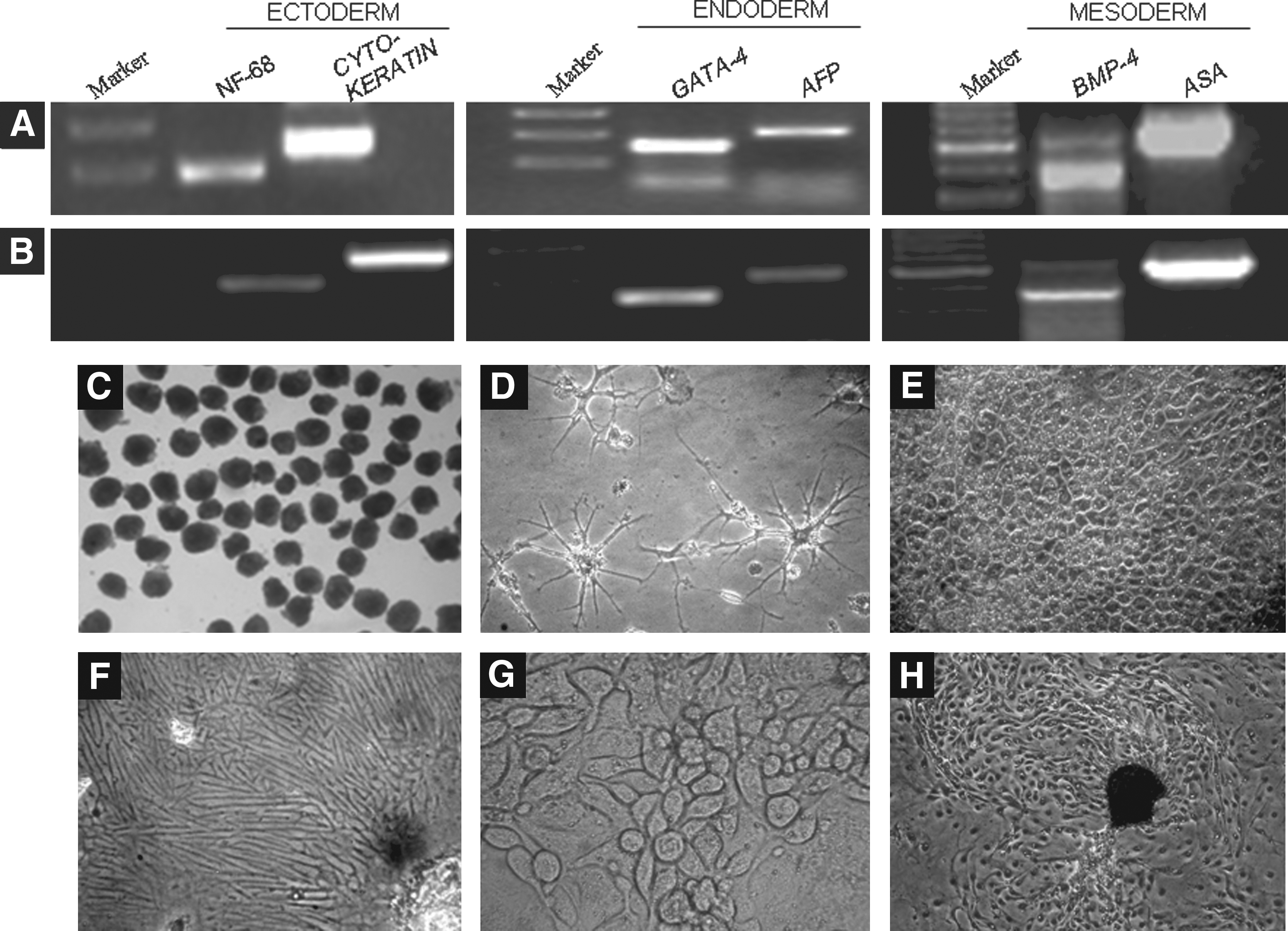
Presence of three germ layer markers in embryoid bodies formed by hanging drop method from lower passage (
Effect of supplementation with FGF-2 on primary colony formation
Following seeding of ICMs isolated mechanically from in vitro produced day 8 hatched blastocysts on buffalo fetal fibroblast feeder layer, primary colonies were formed 6 to 10 days later. The basic organization of a primary colony included a central mass of ICM cells (Fig. 3) followed by a layer of polar trophoblast cells, which were further surrounded by a layer of mural trophoblast cells. The primary colony formation rate was significantly higher (p<0.05) following supplementation with FGF-2 at concentrations ≥5 ng/mL compared to that of control (Table 1).
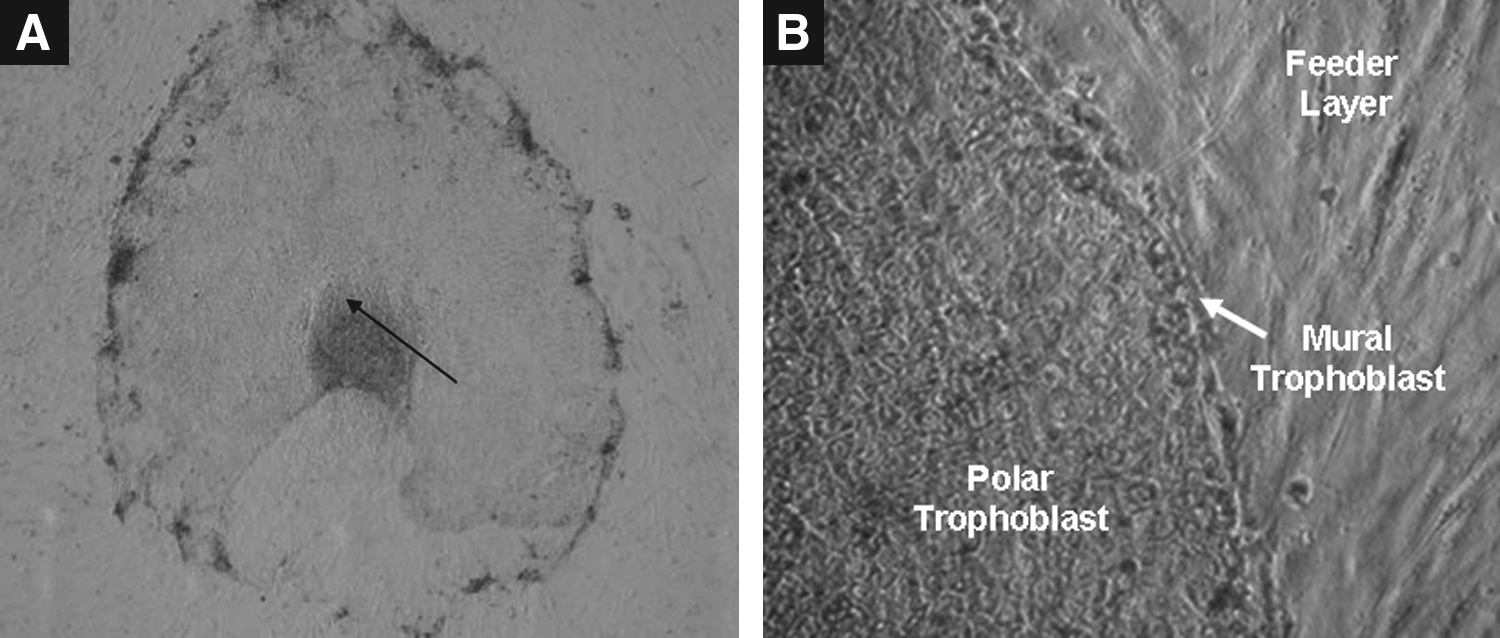
A typical primary colony of ES cell-like cells formed 6 to 10 days after seeding of ICMs isolated mechanically from in vitro produced day 8 hatched blastocysts on buffalo fetal fibroblast feeder layer. The arrow indicates proliferating ICM cells surrounded by trophectoderm (
Data from 10 trials.
Values are mean±SEM.
Values with different superscripts in the same column differ significantly (p<0.05).
Effect of supplementation with FGF-2 and/or LIF on primary colony formation
In Experiment 2, aimed at examining the effect of supplementation of the culture medium with 5 ng/mL FGF-2, alone or in combination with 1000 IU/mL LIF, the primary colony formation rate was found to be significantly higher (p<0.05) following supplementation with both FGF-2 and LIF than that after supplementation with LIF alone or that of the controls (Table 2). The primary colonies formed were observed to be with well-defined edges, and their morphology was similar among all the groups (Supplementary Fig. 3) and had the similar basic organization as discussed above.
Data from 11 trials.
Values are mean±SEM.
Values with different superscripts in the same column differ significantly (p<0.05).
Effect of supplementation with FGF-2 and/or LIF on colony survival and self-renewal
In Experiment 3, among the four groups, the percent survival after 6 days of culture of colonies at passage 15 to 20 was highest when the colonies were cultured in the presence of both FGF-2 and LIF (Table 3). Similar results were obtained when this experiment was repeated with ES cell-like cells' colonies at passage 60 to 90 (data not shown). The self-renewal capacity, determined by the maximum number of passages for which colonies could survive in an undifferentiated state, was also found to be the highest in this group. Out of four experimental groups, only LIF + FGF-2 group could support the self renewal of buffalo ES cell-like cells, as in rest of three groups, colonies either got differentiated or died within 50 to 60 days of culture (10 passages).
Data from four trials.
Values are mean±SEM.
Values with different superscripts in the same column differ significantly (p<0.05).
Effect of supplementation with FGF-2 and/or LIF on expression of pluripotency-related genes
In Experiment 4, the expression of transcription-based pluripotency markers was determined in the ES cell-like cells' colonies belonging to the four supplementation groups by qRT-PCR SYBR green assay. The fold change in mRNA expression normalized to GAPDH was compared to that of the controls (SCM alone). NANOG and SOX-2 expressions were found be decreased (p<0.05) following supplementation with FGF-2 and LIF, respectively, but when the medium was supplemented with both these, the expression of NANOG and SOX-2 was similar to that in the controls. OCT4 expression was not affected following supplementation with FGF-2 or LIF or both (Fig. 4).
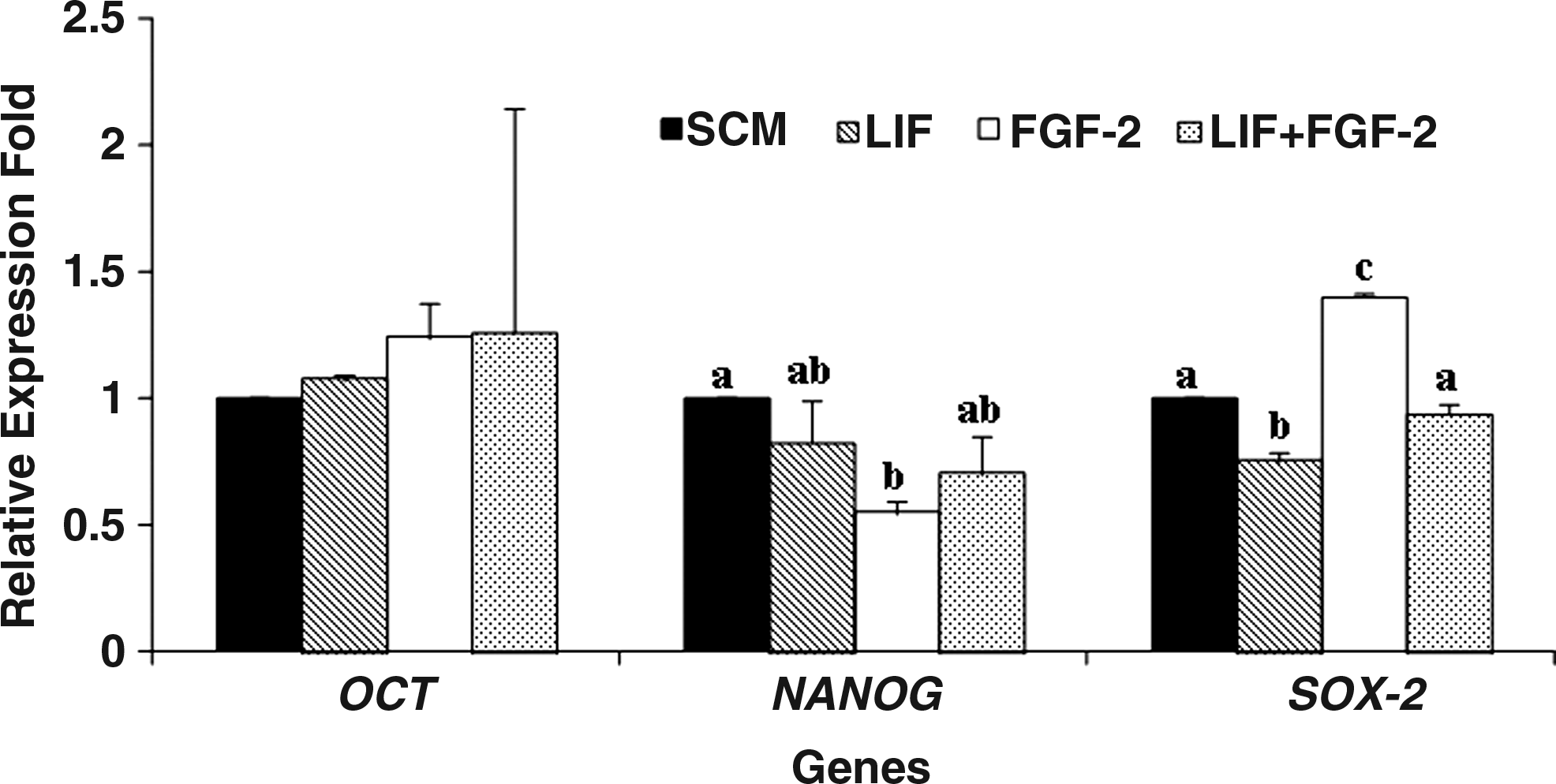
Relative expression levels of pluripotency genes in ES cell-like cells colonies cultured under different conditions. Group 1: stem cell culture (SCM) medium alone; Group 2: SCM + LIF; Group 3: SCM + FGF-2; and Group 4: SCM + LIF + FGF-2.
Effect of supplementation with FGF-2 and/or LIF on colony size
In Experiment 5, among the four groups, the highest (p<0.05) increase in the colony size was observed following supplementation with both FGF-2 and LIF between days 3 and 6 and days 0 and 6 of culture (Fig. 5). The increase in colony size was significantly higher (p<0.05) following supplementation with FGF-2 compared to that after supplementation with LIF or that in the control between days 0 and 6 of culture.

Effect of supplementation of stem cell culture medium (SCM) with FGF-2, alone or in combination with LIF, on the size of ES cell-like cell colonies following culture for 144 h.
Effect of FGFR inhibitor on colony survival
Out of the four FGF receptors, FGFR1, 2, and 3 were detected in the ES cell-like cells. Also, some key signaling intermediates of FGF signaling such as ERK1, K-RAS, E-RAS, and SHP-2 could be detected in the ES cell-like cells by RT-PCR (Fig. 6). In Experiment 6, the percentage of ES cell-like cells' colonies that were found to be differentiated was significantly higher (p<0.05) following supplementation of SCM with SU5402 (Table 4), an FGFR inhibitor, at concentrations of ≥20 μM (Fig. 7).
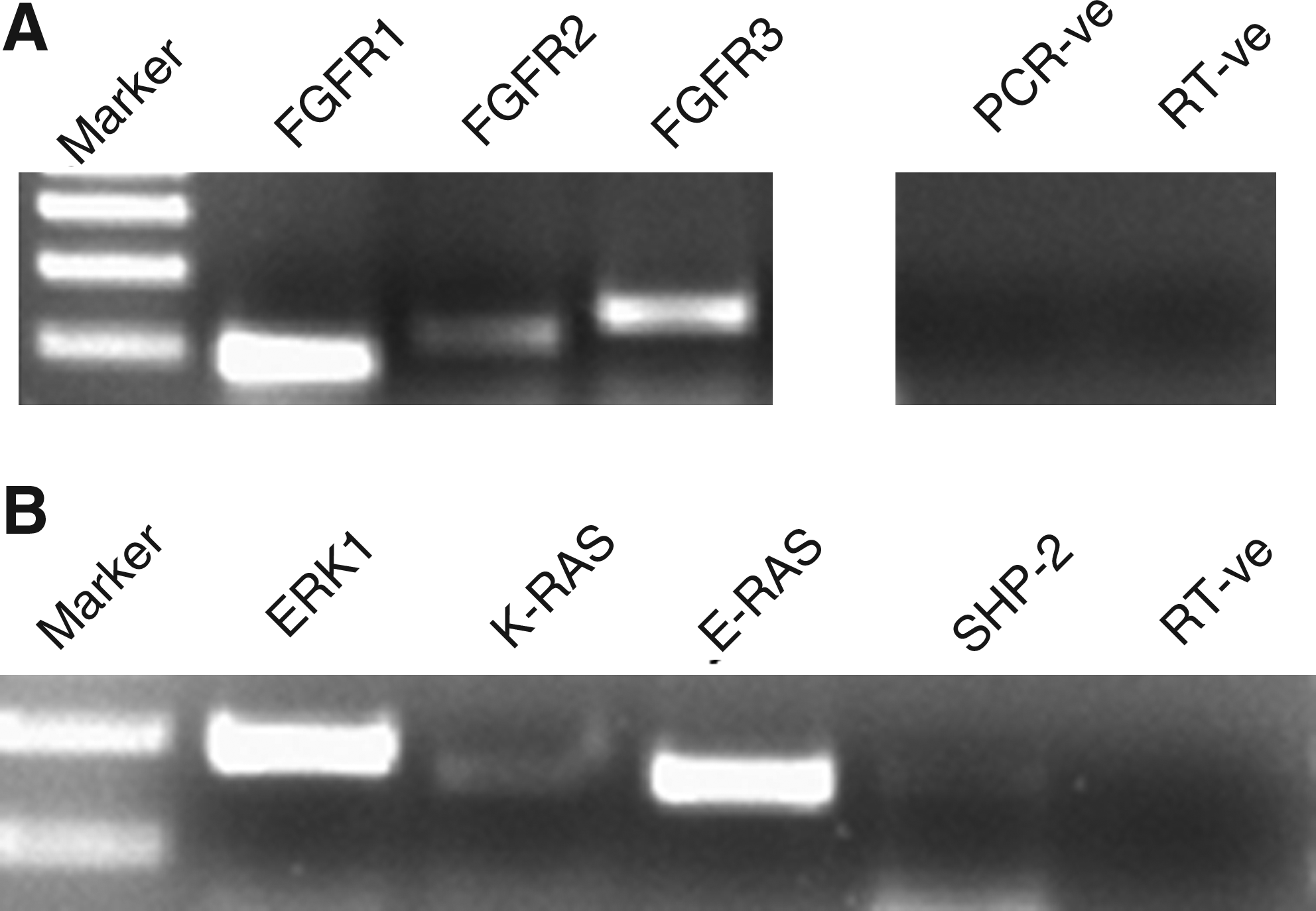
Expression of FGF receptors (

The effect of inhibiting FGF-2 signaling by addition of SU5402 in SCM + LIF. The colonies cultured in SCM + LIF + FGF-2 showed normal morhpology (
Data from five trials.
Values are mean±SEM.
Values with different superscripts in the same column differ significantly (p<0.05).
Effect of supplementation with FGF-2 and/or LIF expression on expression of growth factors
In Experiment 7, buffalo ES cell-like cells were cultured in the presence of FGF and/or LIF, following which the relative expression of growth factors such as Activin-A, BMP-4, TGF-β1, FGF-2, and Gremlin was determined by qRT-PCR. The expression of Activin-A, which was decreased (p<0.05) by FGF-2 was further decreased (p<0.05) by simultaneous supplementation with LIF, although LIF alone had no effect (Fig. 8). BMP-4 expression was not altered by FGF-2 or LIF but was decreased (p<0.05) when medium was supplemented with both of them. The expression of TGF-β1 was increased (p<0.05) following supplementation with FGF-2 or LIF but was decreased (p<0.05) after the medium was supplemented with both of them. Expression of FGF-2 by the buffalo ES cell-like cells was not affected following supplementation with FGF-2, alone or in combination with LIF, whereas supplementation with LIF alone decreased (p<0.05) FGF-2 expression. Gremlin expression was not affected by FGF-2 and LIF, alone or in combination.

Relative expression levels of Activin-A, BMP-4, TGF-β1, FGF-2, and Gremlin in ES cell-like cells' colonies cultured under different conditions. Group 1: stem cell culture medium (SCM) alone; Group 2: SCM + LIF; Group 3: SCM + FGF-2; and Group 4: SCM + LIF + FGF-2.
Discussion
To our knowledge, this is the first report on the development of a culture system that could support sustained self-renewal of buffalo ES cell-like cells in an undifferentiated state over a long period of time. We have been able to derive three ES cell lines that, to date, have been maintained in culture for 135, 95, and 85 passages for over 27, 19, and 17 months, respectively. These cells were monitored for the expression of pluripotency markers OCT4, NANOG, SOX-2, REX-1, FOX-D3, NUCLEOSTAMIN, and STAT-3 by RT-PCR at passages 17, 65, 90, and 110. The expression of SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81 by these cells was confirmed at passages 9, 20, 37, 56, 80, and 110 by immunofluorescence. The ability of these cells to form embryoid bodies and to spontaneously differentiate to various cell types was confirmed at passages 17, 35, 75, and 120. One of the cell lines was successfully used as nucleus donor and a live-cloned calf was produced through HMC (George et al., 2011).
Derivation of ES cell-like cells has been reported from many farm animal species including cattle (Mitalipova et al., 2001), pig, sheep (Piedrahita et al., 1990), and horse (Saito et al., 2002). However, there is only a single report on long-term survival of bovine ES cells in culture (Mitalipova et al., 2001). Use of suboptimal culture conditions, which are generally based empirically on those standardized for mouse or human ES cells, is probably the major reason behind the lack of long-term survival of ES cells in culture, in view of the very high species specificity of the culture requirements. The buffalo ES cell-like cells derived from blastocysts produced parthenogenetically (Sritanaudomchai et al., 2007) or by IVF (Anand et al., 2009; Huang et al., 2010; Kumar et al., 2011; Verma et al., 2007) have not been reported to survive beyond passage 10. Assuming that the primary reason for this could be suboptimal culture conditions employed, the focus of our work was on improving the culture system.
The basic culture medium used in these studies included DMEM or DMEM/F12 (Sritanaudomchai et al., 2007) supplemented with 15 to 20% FBS. In the present study, a combination of KO-DMEM and 15% KO-SR was used instead. Although DMEM has been used for culturing ES cells in a large number of studies, it was developed for general cell culture and may not, therefore, be suitable for the culture of ES cells. KO-DMEM, a propriety preparation, has been found to be superior to DMEM for culturing ES cells in human (Amit et al., 2000) and bovine (Wang et al., 2005). Also, because inclusion of serum as a supplement offers many disadvantages such as promotion of differentiation of ES cells, lack of full characterization, high batch-to-batch variation, bio-safety concerns associated with use of animal sera, etc. A number of supplements that are alternative to serum, known as serum replacers, are now available which, besides being chemically defined, and being formulated on the basis of the specific needs of a particular cell, do not suffer from most of the disadvantages of serum mentioned above. Among these, KO-SR, which is a chemically defined serum replacer, has been found to be suitable for the culture of ES cells in a large number of studies across a wide variety of species. In the present study, the ES cell like cells were cultured on a mitomycin C-inactivated buffalo fetal fibroblast feeder layer, because the homologous feeder layers were found to be superior to heterologous feeder layers such as sheep and goat fetal fibroblast feeder layers for supporting self-renewal of buffalo ES cell-like cells in our earlier study (Kumar et al., 2011).
FGF-2 had been in use for a long time, but its role has not been extensively studied in long-term maintenance of ES cells of farm animals. The major difference between our culture system and those used earlier was the use of FGF-2 at an optimal concentration, in combination with LIF. In the present study, supplementation of SCM with FGF-2 and LIF could support the survival of colonies for more than 135 passages (over a 27-month period), whereas in the absence of both these supplements, the colonies could not survive in an undifferentiated state beyond passage 10. Although LIF (Brevini et al., 2005) or FGF (Munoz et al., 2008) alone or in combination (Gjørret and Maddox-Hyttel., 2005; Miyoshi et al., 2000; Tian et al., 2006) have been used in earlier studies for the culture of bovine, porcine, and caprine ES cell-like cells, the results have been contradictory probably due to high species-specificity in the growth factors and signaling pathways responsible for maintaining pluripotency.
It is now established that in addition to some signaling pathways like Activin/Nodal signaling, which are common across many species like human, murine, rat, rabbit, etc., some signaling pathways are highly species specific. For example, LIF is needed by mouse ES cells for activation of STAT3 via LIF receptor signaling for self-renewal, and LIF works in synergism with BMP-4 for maintaining these cells under feeder-free conditions (Ying et al., 2003). In contrast, LIF/STAT3 signaling fails to maintain the self-renewal of human ES cells (Daheron et al., 2004; Humphrey et al., 2004), and FGF-2 supplementation and suppression of BMP signaling sustained undifferentiated proliferation of human ES cells (Xu et al., 2005). Our results show that buffalo ES cell-like cells resemble human ES cells in terms of the requirement for FGF-2 (Amit et al., 2000; Vallier et al., 2005; Xiao et al., 2006; Xu et al., 2005), but are probably closer to rabbit stem cells because, like them, not only both FGF-2 and LIF are necessary for self-renewal and proliferation but they also are more effective when acting concomitantly (Hsieh et al., 2011). In our study ES cell-like cells' colonies were able to proliferate only in the presence of FGF-2; LIF alone was not able to support the proliferation of ES cell-like cells, as also suggested by previous reports (Anand et al., 2009; Huang et al., 2010; Verma et al., 2008). Also, despite the availability of multiple growth factors like FGFs, activin, transforming growth factor (TGF)-β, Wnts, etc., from feeder layer, buffalo ES cell-like cells cannot survive over a long term in the absence of additional FGF-2. The buffalo ES cell-like cells were found to express the key intermediates of FGF signaling. It was found that the three FGFR kinases, and some key signaling intermediates of FGF signaling such as ERK1, K-RAS, E-RAS, and SHP-2 could be detected in the ES cell-like cells by RT-PCR. The downstream mechanism of FGF in buffalo, therefore, may involve ERK pathway, as in human ES cells (Kang et al., 2005; Li et al., 2007).
The requirement of FGF-2 for the self-renewal of buffalo ES cell-like cells was further confirmed by the observation that SU5402, an FGFR inhibitor, caused the ES cell-like cells' colonies to differentiate. The cell death could be observed at every concentration of SU5402 as single cells were seen floating in culture after every 24 h during medium change. Further, the medium collected was observed for the presence of cells and trypan blue staining was carried to see the viability of cells (data not shown) and more than 95% cells were seen to be dead. In addition to it the colonies were observed to follow two different differentiation patterns, colonies cultured in the presence of 10 μM SU5402 differentiated from the center maintaining the round shape (Supplementary Fig. 1B and C), while colonies cultured with 20 μM and 30 μM SU5402 got flattened with more uniformity (Fig. 7 and Supplementary Fig. 1D–F). The survival rate was most severely affected in the presence of SU5402 at a concentration of 30 μM as merely after 2 to 3 days of exposure the colony size got significantly reduced along with significant loss of alkaline phosphatase activity compared to control colonies (data not shown). Human ES cells exhibits a similar differentiation pattern when treated with SU5402 and were observed to differentiate into neuroepithelium and primitive endodermal cells (Dvoark et al., 2005).
Next, the effects of supplementation of FGF-2 alone or in combination with LIF on quantitative expression of signaling molecules such as Activin-A, BMP-4, TGF-β1, FGF-2, and Gremlin were determined by qRT-PCR. Our results indicate that FGF-2 appears to have a relationship with TGF-β/Activin/Nodal signaling pathway, although further studies are required to find out if there is a cooperation between Activin/Nodal and FGF pathways for the maintenance of pluripotency, as reported earlier in human (Vallier et al., 2005).
Primary colonies could be formed in the present study even in the absence of both LIF and FGF-2, probably because these growth factors were secreted by feeder layer. But the percentage of recalcitrant ICMs (Supplementary Fig. 4A and B{SUPPLFIG4}) was higher in SCM group, which could be the reason of lower primary colony formation rate. Apart from that, the ICMs cultured in SCM and LIF groups exhibited more percentage of ICM cells giving rise to balloon-like structure comprising of trophectodermal cells (Supplementary Fig. 4C and D) or when the cultures were left unattended without passaging. Contrary to Kwon et al. (2009), who reported that the peeling off such balloon-like structures revealed the pure epiblast colony concealed beneath, in our study we could not observe any such colony-like structure although the monolayer of cells could be seen beneath the balloon (Supplementary Fig. 4D). Moreover, the peeling step itself is tedious and requires great precision and accuracy, which further complicates the process. The significant increase in primary colony formation rate following supplementation with both FGF-2 and LIF indicates that there could be a synergistic effect between the two. Despite use of FGF-2 for initial derivation of ES cells in many studies in humans (Camarasa et al., 2010; Inzunza et al., 2005; Strelchenko et al., 2004), and use of a combination of FGF-2 and LIF for deriving ES cell-like cells in cattle (Roach et al., 2006), goat (Tian et al., 2006), and pig (Li et al., 2003), little information is available on the effects of FGF-2 on primary colony formation rate. The primary colony formation rate in our study (20–65%) was in the range as reported in earlier studies (Du Pay et al., 2010; Gong et al., 2010; Stice et al., 1996). In an another report, addition of hLIF (1000 IU/mL) to the culture medium had no significant effect on the ability of porcine ICMs to form primary ES cell colonies, rather there was no positive effect of exogenous supplementation of any of the growth factors studied (Du Pay et al., 2011).
In bovine ICM cultures, NANOG and SOX-2 expression was not detected beyond the initial passage (P-0); however, OCT4 continued to express through P-2. The authors suggested that the loss of expression of pluripotency markers in the ICM explants during the initial days of culture is because of suboptimal culture conditions (Pant and Keefer, 2009). In porcine day 6 embryos, the expression of OCT4 was observed in both ICM and trophectoderm, whereas NANOG and SOX-2 could not be detected in blastocysts containing the ICM (Hall et al., 2009; Kuijk et al., 2008). Comparative analysis of OCT4, NANOG, and SOX-2 within trophectoderm and epiblast of day 11 embryos indicated that these are exclusively expressed in the epiblast (Hall et al., 2009). In our study supplementation with FGF-2 alone decreased (p<0.05) NANOG and increased (p<0.05) SOX-2 expression but simultaneous supplementation with LIF reversed this trend and was able to bring the expression of these transcription factors back to the levels seen in the controls. In our study, with the supplementation of LIF expression of SOX-2 was observed to decrease (p<0.05). The expression of OCT4 was not affected by supplementation with FGF-2 or LIF or both. In human ES cells, members of the TGF-β family such as activin A are also believed to support self-renewal, by collaborating with FGF, via activation of Smad2/3, leading to upregulation of OCT4 and NANOG (Vallier et al., 2005). Also, Wnt signaling regulates expression of OCT4, NANOG, and REX1 (Sato et al., 2004). The role of these signaling pathways and their relationship with FGF signaling in buffalo ES cell-like cells needs to be further examined.
In conclusion, the present study describes a culture system that could support sustained self-renewal of buffalo ES cell-like cells in an undifferentiated state over a long period of time. This system used a combination of bFGF2 and LIF, both of which offered beneficial effects when used alone, but were more effective when used together. These results highlight the importance of FGF and LIF/JAK/STAT3 signaling in the self-renewal of buffalo ES cell-like cells.
Footnotes
Acknowledgments
The present work was funded by National Agriculture Innovative Project (NAIP) grant to M.S.C. (C-2067 and 075) and S.K.S. (C 2-1-(5)/2007). A.G. and R.S. are supported by a CSIR senior research fellowship.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.