
Abstract
Abstract
We examined the influence of recipient oocytes on in vitro development, oxygen consumption, and gene expression in the resulting cloned bovine embryos. Oocytes derived from slaughterhouse ovaries and ovum pickup (OPU)-derived oocytes were used as recipient cytoplasts for the production of cloned embryos. A series of OPU sessions was conducted on Holstein cows without follicular growth treatment (FGT). In the same cows, we then performed dominant follicle ablation and subsequently administered follicle-stimulating hormone and prostaglandin F2α with controlled internal drug release device before a second series of OPU. Cumulus cells collected from single Holstein cows were used as donor cells. After measurement of oxygen consumption at the blastocyst stage with modified scanning electrochemical microscopy, analysis of 10 genes (CDX2, IFN-tau, PLAC8, OCT4, SOX2, NANOG, ATP5A1, GLUT1, AKR1B1, and IGF2R) was performed with real-time RT-PCR. Rates of fusion, cleavage, and blastocyst formation were not different among the treatment groups. Levels of oxygen consumption in cloned blastocysts derived from slaughterhouse ovaries or OPU without FGT were significantly lower than in blastocysts derived from artificial insemination (AI). However, oxygen consumption was increased in cloned blastocysts derived from OPU with FGT, depending on the individual oocyte donor. Furthermore, gene expression of IFN-tau and OCT4 in cloned blastocysts derived from OPU with FGT was similar to that in AI-derived blastocysts, whereas expression of those genes in cloned blastocysts derived from slaughterhouse ovaries or OPU without FGT was significantly different from that in AI-derived blastocysts. Thus, recipient oocytes collected by OPU in combination with manipulation of follicular growth in donor cows are suitable for producing cloned embryos.
Introduction
Two crucial factors of the SCNT process probably affect the reprogramming efficiency: the state of the donor cell and a suitable recipient cytoplast. In mouse studies, the epigenetic status of SCNT embryos, including histone hypermethylation and gene expression, was improved by using a donor with cytoplasmic lysates of germinal vehicle-stage oocytes (Bui et al., 2008) and using a donor cell in which Xist is deleted (Inoue et al., 2010). These manipulations resulted in increased cloning efficiency. In contrast, involvement of the recipient cytoplasm on the reprogramming efficiency has received much less attention, even though the fundamental importance of oocyte cytoplasmic factors for reprogramming transferred nuclei and early embryonic development is well recognized (Bordignon et al., 2001; Fulka and Fulka, 2007). Several reports have indicated that the in vitro developmental competence of cloned embryos can be improved using oocytes with various biological characteristics, including oocytes that are in dibutyryl cyclic AMP-treated early metaphase II (Sugimura et al., 2010b), oocytes derived from prepubertal versus adult animals (Betthauser et al., 2000), and oocytes that were matured in vitro versus in vivo (Akagi et al., 2008). In addition, investigations of the maternal source of oocytes have examined factors such as the effect of certain family lines of oocyte donors on SCNT efficiency (Bruggerhoff et al., 2002) and improvement in the in vitro development of cloned bovine embryos with hybrid recipient oocytes (Yang et al., 2005). Most of these reports focused on in vitro development, whereas factors determining the blastocyst quality such as epigenetic status and metabolic activity, which may determine embryo viability after transfer, were not examined.
For most bovine SCNT techniques, large numbers of oocytes with nondefined genomes are collected from ovaries derived from slaughterhouses. Together with slaughterhouse-derived oocytes, those recovered from ovum pickup (OPU) also are included as an oocyte source. OPU allows repeated retrieval of oocytes from arbitrary females. With in vitro fertilization (IVF) in buffaloes, it was shown that oocytes derived from live animals by OPU resulted in a significantly higher blastocyst yield than those derived from slaughterhouse ovaries (Manjunatha et al., 2008). Several studies have been performed in an attempt to establish the most cost-effective procedure for retrieving the highest number of high-quality oocytes using OPU (Blondin et al., 2002; Chaubal et al., 2007; Goodhand et al., 1999). Follicular growth induced by follicle-stimulating hormone (FSH) has been used routinely to rescue follicles back into a developmental stage that would otherwise proceed to atresia (Merton et al., 2003). Furthermore, the relationship between the oocyte and the follicle cells, including cumulus cells and granulosa cells, is influenced by hormonal signals (Eppig et al., 1997, 1998; Norris et al., 2009), which are crucial for the acquisition of oocyte developmental competence, commonly referred to as cytoplasmic maturation.
The aim of our current study was to examine the influence of the oocyte source and follicular growth treatment (FGT), consisting of dominant follicle ablation and subsequent FSH and prostaglandin F2α (PGF2α) administration with controlled internal drug release (CIDR) device before OPU, on the in vitro development, oxygen consumption, and gene expression of cloned bovine embryos. We used oocytes derived from slaughterhouse ovaries and OPU with or without FGT to produce cloned embryos.
Materials and Methods
Animal care and use
This study was approved by the Ethics Committee for the Care and Use of Experimental Animals at the National Livestock Breeding Center located in Nishigo, Japan. All animals received humane care according to law no. 105 and notifications no. 6 and no. 22 of the Japanese Guidelines for Animal Care and Use.
Oocyte collection from slaughterhouse ovaries
Collection of bovine cumulus–oocyte complexes (COCs) was performed as described (Tagawa et al., 2008). Ovaries from Japanese Black or Holstein heifers were collected at a local slaughterhouse, transferred to the laboratory, washed, and stored in physiological saline supplemented with 50 μg/mL gentamycin (Sigma Chemical, St. Louis, MO, USA) at 20°C for ∼20 h. COCs were aspirated from small follicles (2–6 mm in diameter) using a 5-mL syringe equipped with a 19-gauge needle.
Oocyte collection via OPU
As described by Imai et al. (2006), COCs were collected from eight Holstein cows by OPU using an ultrasound scanner (SSD-900; ALOKA, Tokyo, Japan) and 7.5-MHz convex-array transducer (UST-9109P-7.5; ALOKA) with a 17-gauge stainless steel needle guide. Follicles were categorized as small (2–4 mm in diameter), medium (5–8 mm), and large (>8 mm). Follicles >2 mm in diameter were vacuum-aspirated (120 mmHg and 22 mL/min aspiration rate) through a disposable aspiration needle (COVA Needle; Misawa Medical, Tokyo, Japan).
In vitro maturation (IVM)
The IVM medium was 25 mM HEPES-buffered TCM199 (M199; Gibco, Grand Island, NY, USA) supplemented with 5% calf serum (CS; Gibco). Five to 10 COCs were washed twice with IVM medium and incubated in 50 μL IVM medium covered with paraffin oil (Paraffin Liquid; Nacalai Tesque, Inc., Kyoto, Japan) in 35-mm Petri dishes (Nunclon Multidishes; Nalge Nunc International, Roskilde, Denmark) at 38.5°C in a humidified atmosphere of 5% CO2 in air for 18–20 h.
Follicular growth treatment (FGT)
As shown in Figure 1, the first OPU session (OPU without FGT) was performed in eight Holstein cows at arbitrary days of the estrous cycle. Then, follicles larger than 8 mm in diameter were aspirated, and CIDR device (CIDR® 1900; Pfizer Animal Health, Hamilton, New Zealand) was inserted on day 5 (the day of the first OPU session=day 0). Cows then received 30 mg FSH (ANTORIN®-R•10; Kyoritsu Seiyaku, Tokyo, Japan) twice a day from days 7 to 10 in decreasing doses (6, 6, 4, 4, 3, 3, 2, and 2 mg) by intramuscular injection. PGF2α (0.75 mg; Cloprostenol; Clopromate C; Sumitomo Pharmaceuticals Co., Tokyo, Japan) was administered in the morning of day 9. The second OPU session (OPU with FGT) was performed 48 h after PGF2α administration (day 11), and only follicles larger than 5 mm in diameter (i.e., medium and large follicles) were aspirated. CIDR was removed from the cows just before OPU.
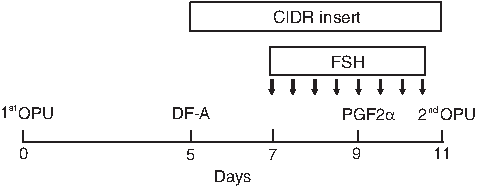
Scheme of OPU with FGT. OPU without FGT (first OPU) was performed at arbitrary days of the estrous cycle. Then, follicles larger than 8 mm in diameter were ablated (DF-A), and CIDRs were inserted on day 5 (the day of first OPU=day 0). The cows then received 30 mg FSH twice a day from days 7 to 10 in decreasing doses (6, 6, 4, 4, 3, 3, 2, and 2 mg) by intramuscular injection. Arrows indicate FSH administration. Cloprostenol (PGF2α; 0.75 mg) was administered in the morning of day 9. OPU with FGT (second OPU) was performed 48 h after PGF2α administration (day 11), and only follicles larger than 5 mm in diameter (medium and large) were aspirated. CIDRs were removed from the cows just before OPU.
Preparation of donor cells
Donor cells were prepared using a slight modification of the methods described in a previous report by Goto et al. (1999). COCs were aspirated from a single Holstein cow by OPU. After in vitro culture of COCs in TCM199 medium supplemented with 5% CS for 20 h, cumulus cells were removed by gentle pipetting. The cumulus cells were incubated in Dulbecco's modified Eagle medium (DMEM; Gibco) supplemented with 10% CS at 38.5°C in a humidified atmosphere of 5% CO2 in air. The culture medium was replaced every 2 days until the cells reached confluence. The cells then were harvested with 0.05% trypsin-EDTA solution (Trypsin-EDTA; Gibco) for 5 min at 38.5°C. Cells then were suspended in modified Dulbecco's phosphate-buffered saline (Modified D-PBS; Gibco), centrifuged at 540×g for 5 min, frozen in DMEM supplemented with 20% fetal calf serum (FCS; Gibco), 1.5 M ethylene glycol (Wako, Osaka, Japan), and 0.1 M sucrose (Research Organics, Cleveland, OH, USA), and stored in liquid nitrogen (Passage 0). Before nuclear transfer, the confluent cells were cultured in DMEM with 0.5% CS for 3 days at 38.5°C in a humidified atmosphere of 5% CO2 in air. The cells were used between passages 3 and 5.
Production of SCNT embryos
After IVM, cumulus cells were removed by pipetting in 0.1% hyaluronidase (Sigma), and oocytes with the first polar body (PB) were enucleated in HEPES-buffered TCM199 supplemented with 5% CS and were used to produce SCNT embryos. A portion of the zona pellucida near the first PB was cut with a cutting needle before enucleation. A small volume of cytoplasm surrounding the PB was squeezed out though a slit made by the cutting needle. The first PB and putative karyoplast were stained with 2 μg/mL Hoechst 33342 (Calbiochem, La Jolla, CA, USA). Enucleated oocytes were identified with fluorescent microscopy (Olympus, Tokyo, Japan). The cumulus cells were transferred into the perivitelline spaces of the enucleated oocytes. Nuclear-transferred couplets were subsequently fused with one DC pulse of 25 V for 25 μsec using a LF101 fusion machine (TR tech, Tokyo, Japan) with a needle-type electrode in Zimmermann's mammalian cell fusion medium. The couplets then were cultured in HEPES-buffered TCM 199 supplemented with 5% CS and 10 μg/mL cycloheximide (Sigma Chemical) for 5 h. After washing with Charles Rosenkrans 1 medium with amino acid (CR1aa) supplemented with 5% CS (Rosenkrans et al., 1993), 10–20 fused embryos were cultured in a droplet of 100 μL CR1aa supplemented with 5% CS in a humidified atmosphere of 5% CO2 in air for 7 days.
Collection of blastocysts derived from artificial insemination (AI)
In vivo-derived embryos were collected as described by Hashiyada et al. (2005). In vivo embryos were collected from superovulated Holstein cows. Superovulation was performed by injecting 30 A.U. FSH and 2 mL PGF2α and was followed by AI. On day 8 after AI, embryos were recovered by uterine flushing. Blastocysts and expanded blastocyst-stage embryos were selected and stored in D-PBS (Gibco) containing 20% CS.
Measurement of oxygen consumption
Oxygen consumption by individual bovine blastocysts was measured noninvasively with a scanning electrochemical microscopy system (HV-405; Hokuto Denko Co., Tokyo, Japan). Embryos were transferred individually to a plate filled with 5 mL embryo respiration assay medium-2 (ERAM-2; Research Institute for the Functional Peptides, Yamagata, Japan), and the embryos were dropped individually to the bottom of the microwell. The temperature of the medium was maintained at 38.5°C by placing the plate on a warming plate on the microscope stage. Oxygen consumption was measured as described by Shiku et al. (2001). The XYZ-stage and the potentiostat were controlled by HV-405 ver. 2.04 (Hokuto Denko). Voltammetry of the Pt-microdisc electrode (Hokuto Denko) in ERAM-2 solution showed a steady-state oxygen reduction wave. No response from other electrochemically active species was observed near the embryo surface. The oxygen consumption rate of embryos was calculated using software. The oxygen concentration difference between the bulk solution and the sample surface (ΔC) and the oxygen consumption rate (F) of a single sample were estimated according to spherical diffusion theories (Shiku et al., 2001). We repeatedly scanned the electrode back and forth to estimate the mean of the ΔC for each sample two times.
Quantitative real-time RT-PCR analysis
Eleven genes were analyzed with real-time PCR (Table 1). Individual blastocysts were lysed in 50 μL of extraction buffer (Arcturus, Carlsbad, CA, USA), incubated at 42°C for 30 min, and stored at −80°C. Total RNA was extracted from each sample using a PicoPure RNA Isolation kit (Arcturus) according to the manufacturer's instructions. Residual genomic DNA was removed with recombinant RNase-free DNase I (Roche, Mannheim, Germany). RNA was eluted from the purification column using 11 μL of elution buffer (Arcturus). RNA was reverse transcribed into cDNA using a RevaTra Ace qPCR RT kit (Toyobo Bio, Osaka, Japan) and following the manufacturer's instructions. Each 20-μL reaction product was diluted with nuclease-free water to a final volume of 40 μL. Quantitative real-time RT-PCR was performed using StepOnePlus™ Systems (Applied Biosystems, Foster City, CA, USA) in a 20-μL reaction volume containing 2 μL cDNA, 0.5 μL each of forward and reverse primers to amplify histone H2Az (H2AFZ), octamer-binding transcription factor 4 (OCT4), SRY (sex determining region Y)-box containing gene 2 (SOX2), nanog homeobox (NANOG), caudal type homeobox 2 (CDX2), interferon tau (IFN-tau), placenta-specific 8 (PLAC8), ATP synthase, H+ transporting, mitochondrial F1 complex, alpha subunit 1, cardiac muscle (ATP5A1), glucose transporter 1 (GLUT1), aldo-keto reductase family 1 member B1 (AKR1B1), insulin-like growth factor 2 receptor (IGF2R), 7 μL nuclease-free water, and 10 μL Fast SYBR Green PCR Master Mix (Applied Biosystems). In each PCR reaction, duplicate cDNA samples were run to control for the reproducibility of the real-time RT-PCR results. Universal thermal cycling parameters (initial step of 20 sec at 95°C, followed by 45 cycles of 3 sec at 95°C, 10 sec at 60°C, and 20 sec at 72°C) were used to quantify the expression of each gene. After the end of the last cycle, a melting curve program was run. A standard curve was generated for both the target and the endogenous control gene using serial 10-fold dilutions of amplified cDNA derived from blastocysts in every PCR run. Final quantitative analysis was performed with the relative standard method, and results were reported as relative expression after normalization of the transcript amounts to H2AFZ (the endogenous control gene). The quality of the PCR products was confirmed with melting curve analysis.
Statistic analysis
Data were analyzed using analysis of variance (ANOVA) followed by a Tukey-Kramer test or least-squares regression. All percentage data were arcsine transformed. For all data, p<0.05 was considered significant. All analyses were conducted using StatView (SAS Institute Inc., Cary, NC, USA).
Results
Effect of oocyte source and FGT on in vitro development
As expected, our developed FGT protocol affected follicle growth in Holstein cows by significantly increasing the incidence of large follicles (>8 mm) and significantly decreasing the incidence of small follicles (2–4 mm) compared to the nonstimulated condition (Table 2). Although the rate of blastocyst formation in cloned embryos using oocytes derived from OPU with FGT was increased by 1.5 times compared to the rate obtained using oocytes derived from slaughterhouse ovaries, the difference was not significant (p>0.05). The rates of fusion and cleavage also did not differ significantly among the groups (Table 3).
FGT, follicular growth treatment.
Different letters indicate significant differences (p<0.05).
OPU, ovum pickup; FGT, follicular growth treatment.
FGT was performed before OPU.
Effect of oocyte source and follicular treatment on oxygen consumption
Oxygen consumption in cloned blastocysts using oocytes derived from slaughterhouse ovaries (mean±SD: 0.50±0.19×10–14 mol sec–1) or OPU without FGT (0.47±0.25×10–14 mol sec–1) was significantly lower than that in AI-derived blastocysts (0.81±0.25×10–14 mol sec–1). However, oxygen consumption was increased in cloned blastocysts using oocytes that were recovered from FGT animals using OPU (0.67±0.29×10–14 mol sec–1) and was not significantly different from that in AI-derived blastocysts (Fig. 2). Regarding three donors from which more than two blastocysts were produced from both OPU without and with FGT, blastocysts from two donors, H184 and H475, appeared to show improved oxygen consumption after OPU with FGT as observed some of blastocysts with physiological level of oxygen consumption that was similar to that in AI-derived blastocysts. The significant difference between OPU without FGT (0.43±0.20×10–14 mol sec–1) and OPU with FGT (0.91±0.27×10–14 mol sec–1) was observed in H475 (Fig. 3).
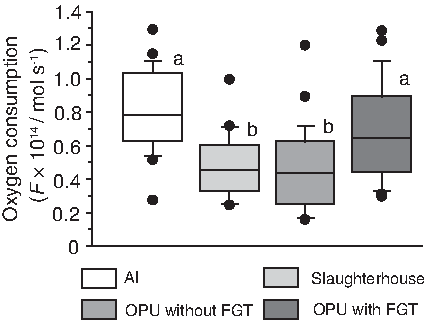
Effect of oocyte source and FGT on oxygen consumption in cloned bovine blastocysts. Oxygen consumption in blastocysts derived from AI (n=24) and cloned blastocysts using oocytes derived from slaughterhouse ovaries (n=20), OPU without FGT (n=25), and OPU with FGT (n=19) was measured individually with scanning electrochemical microscopy. Boxes reflect two quartiles, the 25th and 75th percentiles, and the interior horizontal line indicates the median. Whiskers indicate maximum and minimum values within the acceptable range defined by the two quartiles. Filled circles denote outliers. a,bValues with different superscripts are significantly different.
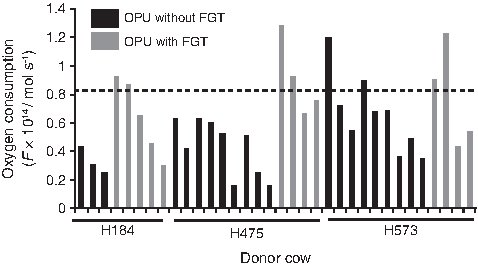
Oxygen consumption in individual cloned bovine blastocysts using oocytes derived from a donor cow. The oxygen consumption in individual cloned blastocysts with oocytes derived from three donor cows (H184, H475, and H573), which were produced from oocytes derived from both OPU without FGT and OPU with FGT, are shown as each bar. The dotted line indicates the mean value of oxygen consumption (0.81×10–14 mol sec–1) in AI-derived blastocysts.
Effect of oocyte source and follicular stimulation on gene expression
Analysis of 10 genes (OCT4, SOX2, NANOG, CDX2, IFN-tau, PLAC8, ATP5A1, GLUT1, AKR1B1, and IGF2R) was performed by real-time RT-PCR. The mean relative expression level of PLAC8 in cloned blastocysts derived from slaughterhouse ovaries was significantly lower than that in AI-derived blastocysts, but the expression level of this gene in cloned blastocysts derived from OPU with or without FGT was similar to that in AI-derived blastocysts (Fig. 4). Expression levels of OCT4 and IFN-tau in blastocysts derived from OPU with FGT were similar to those in AI-derived blastocysts. The relationship between the expression of IFN-tau and that of OCT4 in cloned blastocysts was characterized as a negative correlation, with r=0.33 (Fig. 5). Expression levels of SOX2, NANOG, and IGF2R were not significantly different among the groups (Fig. 4). On the other hand, expression levels of CDX2, ATP5A1, GLUT1, and AKR1B1 were significantly different between AI and cloned blastocysts, regardless of the oocyte source and whether FGT was used before OPU, suggesting that aberrant expression of these genes depends on factors other than oocyte source and FGT in recipient oocytes (Fig. 4).
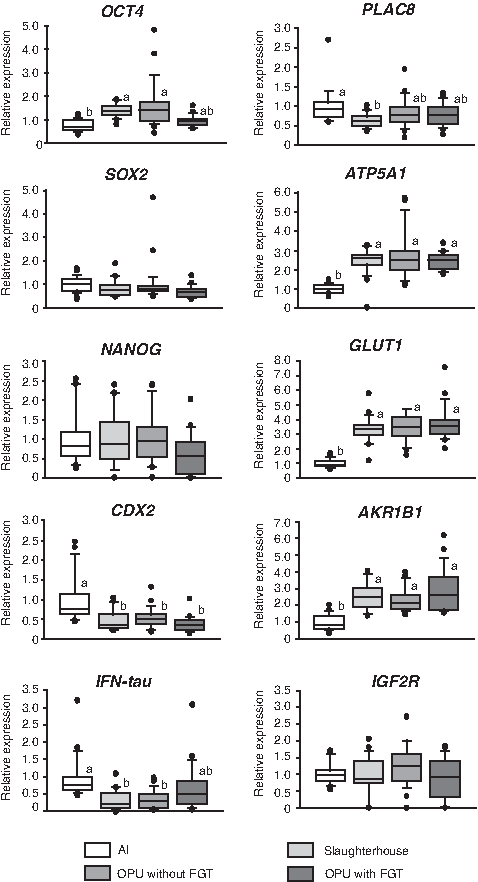
Effect of oocyte source and FGT on gene expression in bovine cloned blastocysts. Expression of 10 genes (OCT4, SOX2, NANOG, CDX2, IFN-tau, PLAC8, ATP5A1, GLUT1, AKR1B1, and IGF2R) in blastocysts derived from AI (n=24) and cloned blastocysts using oocytes derived from slaughterhouse ovaries (n=20), OPU without FGT (n=25), and OPU with FGT (n=19) was measured individually with real-time RT-PCR. Relative expression is presented as the fold change from the mean value of AI-derived blastocysts, which was set at 1. Boxes reflect two quartiles, the 25th and 75th percentiles, and the interior horizontal line indicates the median. Whiskers indicate maximum and minimum values within the acceptable range defined by the two quartiles. Filled circles denote outliers. a,bValues with different superscripts are significantly different.
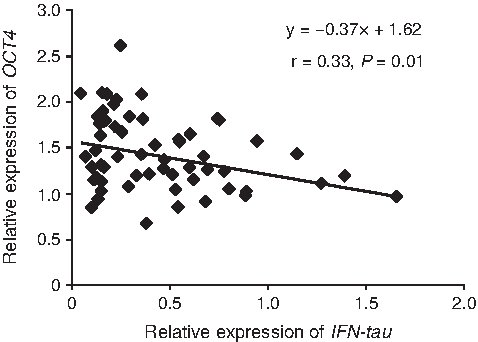
Relationship between expression of IFN-tau and OCT4 in cloned bovine blastocysts. The graph was depicted from overall cloned blastocysts. The coefficient of determination, r, was statistically significant.
Relationship between oxygen consumption and expression of OCT4
To clarify the relationship between oxygen consumption and expression level of OCT4, a correlation coefficient was calculated. The level of oxygen consumption in cloned blastocysts was negatively correlated with the expression of OCT4 (r=0.40, p<0.01), suggesting that the oxygen consumption level reflected expression level of OCT4 (Fig. 6).
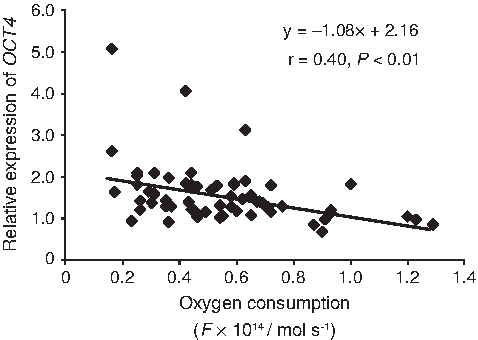
Relationship between oxygen consumption and expression of OCT4 in cloned bovine blastocysts. The graph was depicted from overall cloned blastocysts. The coefficient of determination, r, was statistically significant.
Discussion
We examined the influence of the oocyte source and FGT on in vitro development, oxygen consumption, and gene expression in cloned bovine embryos. Our results indicated that oocytes derived from OPU, regardless of FGT, did not show a significantly improved rate of blastocyst formation in cloned embryos (Table 3). The intraovarian environment to which oocytes were exposed appears to play a major role in determining developmental competence in vitro. Follicular stimulation with FSH involves signals between the oocyte, cumulus cells, and granulosa cells, including growth and differentiation factor 9, bone morphogenetic proteins (BMP) 15 and 6, and oocyte-secreted protein, all of which are involved in promoting the signaling essential for the acquisition of competence to undergo normal embryogenesis (Eppig et al., 2002). In OPU-IVF, FSH stimulation before OPU improves in vitro developmental competence compared to OPU without FSH stimulation, suggesting that cytoplasmic maturation is acquired progressively during the course of follicle development (Berlinguer et al., 2004; Chaubal et al., 2007; Presicce et al., 2011; Schramm and Bavister 1994). Although this difference has not been explained, oocyte factors required for in vitro development may differ between IVF and SCNT (Wakai et al., 2008).
On the other hand, the low oxygen consumption in cloned blastocysts was offset in oocytes derived from FGT cows, depending on the individual oocyte donor (Figs. 2 and 3). To our knowledge, this is the first report that oxygen consumption in cloned embryos is affected by the recipient oocytes, not just the donor cells. In general, oxygen consumption is a well-recognized indicator of the overall metabolic activity of individual embryos because ATP is mostly generated by oxidative phosphorylation (OXPHOS) in mitochondria, a process in which oxygen plays a key role. We previously demonstrated in a porcine study that SCNT embryos at the blastocyst stage have low oxygen consumption, which could be a sign of limited cell proliferation in the trophectoderm and increased apoptosis (Sugimura et al., 2010a). In that study, we hypothesized that low oxygen consumption in cloned embryos may be associated with incomplete reprogramming of donor cells. Incomplete reprogramming appears to cause disorders in mitochondrial function because OXPHOS is strictly regulated by communication between the nucleus and the cytoplasm (Dumollard et al., 2009). Our finding suggests that FGT donor cows may provide better oocytes, in terms of nucleocytoplasmic crosstalk. Improved oxygen consumption appears to be positively related to posttransfer viability such as implantation and fetal growth (Lopes et al., 2007; Wakefield et al., 2011). This question will be evaluated in further studies with embryo transfer.
Inheritance of epigenetic memories in donor nuclei and improper reactivation of embryonic genes that result from incomplete reprogramming of nuclei are reflected by gene expression in cloned embryos (Ng and Gurdon, 2005). We observed that the expression of OCT4 and IFN-tau was improved in oocytes derived from FGT cows. Our findings suggest that cytoplasmic changes during follicular growth have different effects on transcripts in cloned embryos and that reprogramming efficiency, as shown by expression of transcription factors such as RYBP, YY1, and HDASC2 in matured oocytes, is altered by FSH stimulation (Zheng et al., 2005). In our current study, although we cannot conclude definitively whether recipient cytoplasm from follicle-stimulated animals is optimal, in terms of reprogramming efficiency, our results suggest the use of IFN-tau as a possible indicator of the developmental potential of cloned bovine embryos (Kato et al., 2007). IFN-tau acts on the uterine epithelium and attenuates secretion of a luteolysin, PGF2α, resulting in maintenance of corpus luteum function (Vallet et al., 1988). IFN-tau has a pivotal role in the establishment of pregnancy, not only as the major signal for maternal recognition of pregnancy in ruminants, but also as an inducer of several endometrial proteins that may be critical for the survival of developing embryos. Ezashi et al. (2001) reported that ETS2-induced transactivation of the bovine IFN-tau promoter is repressed by POU5F1 through direct interaction of ETS2 with POU5F1, indicating that INF-tau expression is tightly regulated by OCT4. In fact, we also observed a negative correlation between expression of IFN-tau and OCT4 (Fig. 5). The OCT4-dependent transcriptional network may be involved in epigenetics, chromatin remodeling, apoptosis, and metabolic pathways such as OXPHOS, and it may point to underlying molecular mechanisms that regulate pluripotency and trophoblast differentiation in humans (Babaie et al., 2007). Although further studies will be needed, follicle-stimulated cytoplasmic characteristics such as gene expression, epigenetic modification, metabolism, and homeostasis may contribute to gene expression of transcription factors such as OCT4 in cloned embryos. This will result in regulation of not only key genes essential for the establishment and recognition of pregnancy (Zheng et al., 2005), but also embryonic metabolic functions such as oxygen consumption (Fig. 6).
In conclusion, we show that recipient oocytes obtained from follicular growth-stimulated cows have characteristic that improve oxygen consumption and gene expression in cloned embryos, suggesting that the recipient oocyte may be a key factor for determining the reprogramming efficiency of donor nuclei. Further studies using a combination of FGT and OPU for understanding physiological and molecular biological features of oocytes from animals with and without follicular stimulation will lead to valuable insights into the role of the recipient cytoplasm in developmental competence and reprogramming factors.
Footnotes
Acknowledgments
The authors thank Kazunori Fujita, Yoshiaki Horie, Masaru Azuma, Masatoshi Mori, Michiyo Omata, Nubuhiro Komatsu, and Ohya Tomoyuki for animal management, and Michiko Oguro and Saiko Oishi for technical assistance, and Noboru Hitomi for transportation of ovaries.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.