
Abstract
Abstract
Increased possibility of universality of ooplasmic reprogramming factors resulted in a parallel increased interest to use interspecies somatic cell nuclear transfer (iSCNT) to address basic questions of developmental biology and to improve the feasibility of cell therapy. In this study, the interactions between human somatic cells and ovine oocytes were investigated. Nuclear remodeling events were first observed 3 h post-iSCNT as nuclear swelling, chromosome condensation, and spindle formation. A time-dependent decrease in maturation promoting activity of inactivated reconstructs coincided with increased aberrations in chromosome and spindle organization of the newly developed embryos. The sequence and duration of nuclear remodeling events were irrespective of donor cell type used. Although the majority of the reconstituted embryos arrested before embryonic genome activation (8–16-cell) stage, less than 5% of them could progress beyond transcription-requiring developmental stage and formed blastocyst-like structures with distinct inner cell mass and trophectoderm at days 7 and 8 post-SCNT. Importantly, real-time assessment of three developmentally important genes (Oct4, Sox2, and Nanog) indicated their upregulation in iSCNT blastocysts. Blastocyst-derived outgrowths had alkaline phosphatase activity that was lost upon passage. Collectively, this study introduced ovine oocyte as a credible cytoplast for remodeling and reprogramming of human somatic cells back to the embryonic stage and provided a platform for further studies to unravel possible differences exist between reprogramming ability of oocytes of different mammalian species.
Introduction
Importantly, a growing body of evidence indicates that oocyte may have a capacity to reprogram somatic cells from distant taxonomical relations (Tecirlioglu et al., 2006). This implies that the reprogramming molecules of oocytes may be conserved among mammalian species (Dominko et al., 1999; Vallée et al., 2008). Accordingly, there is a growing interest to investigate if nonhuman oocytes can be used to generate patient-specific embryonic stem cells (ESCs) (Chung et al., 2010; Hall et al., 2006).
Human interorder SCNT (iSCNT) has been the subject of a number of studies, although the results are contradictory with those that could produce iSCNT blastocysts (Chang et al., 2003; Chen et al., 2003; Illmensee et al., 2006), and those that did not observe even embryonic genome activation (EGA) (Chung et al., 2010). By reviewing the available studies describing iSCNT blastocyst development between species with distant taxonomical relations, Tecirlioglu et al. (2006) suggested that if the oocyte of a certain species has the required potential to reprogram somatic cell nuclei and reset the embryonic–gene expression pattern, the resulted iSCNT blastocyst could be the source of ESCs line. Importantly, Sha et al. (2009) demonstrated that close relatedness between exotic nucleus and the cytoplast had no obvious effect on preimplantation development of iSCNT embryos, but can improve their capacity for postimplantation development. These observations provide a promising platform for further studies to unravel the molecular mechanism(s) of oocyte-mediated reprogramming of differentiated somatic cell, and to address a universal nonhuman cytoplast for possible therapeutic iSCNT purposes. In this study, the capability of ovine oocytes to induce human somatic cells remodeling and reprogramming was investigated.
Materials and Methods
Chemicals and media were obtained from Sigma (St. Louis, MO, USA) and Gibco (BRL, Grand Island, NY, USA) companies, respectively, unless otherwise specified.
Ethics
The complete experimental design was reviewed and approved by Regional Committee of Ethics at Royan Institute (www.royaninstitute.org, Guideline No. 158).
Oocyte in vitro maturation and SCNT
Abattoir-derived ovine oocytes were used for in vitro maturation (IVM) as described previously (Asgari et al., 2011). Human cumulus cells were retrieved from six (HC1–6), 30–40-year-old patients who enrolled in regular introcytoplasmic sperm injection (ICSI) programs, and human fibroblasts were donated by three (HF1–3) female patients, 25–40 years old who were referred for cell therapy. Somatic cells were serum starved for a period of 4–5 days before nuclear transfer. The process of SCNT was according to Lee et al. (2003) as illustrated in Figure 1A. Reconstituted oocytes were rested for up to 4 h and then activated as described previously (Hosseini et al., 2006). Activated oocytes were cultured in Menezo-B2 medium (INRA, France) over vero-cells monolayer (Moulavi et al., 2006) for 9 days at 38.5°C, 5% O2, 5% CO2, and maximum humidity. Cryopreserved ovine fibroblast and cumulus cells prepared during a recent study (Hosseini et al., 2008) were used for intraspecies SCNT.
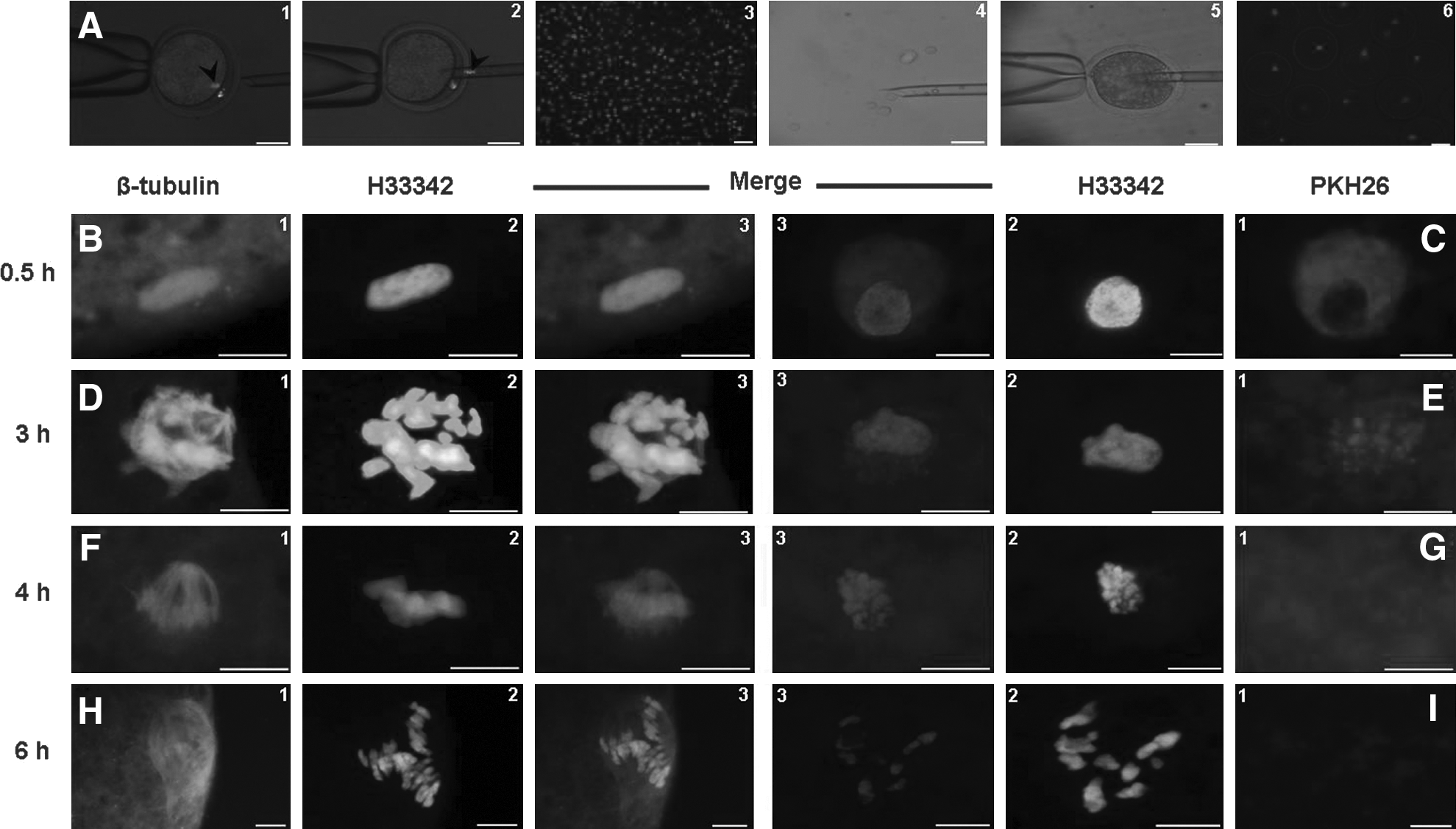
(
Nuclear remodeling and maturation promoting factor (MPF)
Immediately before oocyte reconstitution, the plasma membranes of donor cells were stained with PKH26-red fluorescent cell liner kit (Sigma-Aldrich) according to the manufacturer's protocol (Fig. 1A). At 0.5, 2, 3, 4, and 6 h postreconstitution, oocytes were fixed in PF (15 min), counterstained with 2 μg/mL H33342, and observed using epifluorescent microscope (Olympus BX51, Japan) (excitation: 551 nm, emission: 567 nm). Simultaneously, microtubule and chromosome organizations of the cloned embryos were assessed using immunostaining with anti-β-tubulin monoclonal primary antibody (1:100) in phosphate-buffered saline (PBS) and FITC-labeled antimouse-IgG secondary antibody (1:100) as described previously (Asgari et al., 2011). Moreover, assuming the relationship between remodeling capability of oocyte and MPF activity (Campbell et al., 1993), relative abundances of cyclin-B (the rate-limiting subunit of MPF) transcripts during different intervals postreconstitution were assessed using real-time RT-PCR as described later.
Clone verification
To detect the presence of human DNA in cloned embryos, iSCNT embryos and positive (human cells) and negative (ovine cells/embryos) controls were fixed in PF (4%, 15 min), permeabilized in Triton X-100 (0.2%, 1 h), blocked with bovine serum albumin (BSA) (2%, 1 h) and then incubated with mouse antihuman Ribonucleoprotein (ah-RNP, 1:500, AbD-Serotec, UK) overnight at 4°C. Samples were then incubated with FITC-conjugated secondary antibody (1:1000, 1 h) and counterstained with propidium iodide (1 μg/mL, 15 min) before being observed using epiflouresence microscope (excitation: 494 nm, emission: 518 nm).
DNA was extracted from iSCNT embryos and their corresponding donor cells using QIAamp DNA Micro Kit (Qiagen, Germany) and used for PCR amplification with special annealing temperature for specific designed primers (Table 1) as described previously (Nasr-Esfahani et al., 2011).
Karyotyping and PCR amplification of mtDNA
Human iSCNT embryos were used for karyotyping as described by Change et al. (2003). Human and ovine cloned embryos were also used for PCR amplification of mtDNA using specific primers (Table 1) described by Kristinsson and Lewis (2009) and Li et al. (2005). Accordingly, human mtDNA was amplified using the primers: 5′-CACCATTAGCACCCAAAGCT-3′ and 5′-CTGTTAAAAGTGCATACCGCCA-3′. Ovine mtDNA was amplified using the primers: 5′-AACTGCTTGACCGTACATAGTA-3′ and 5′-AGAAGGGTATAAAGCACCGCC-3′. Appropriate positive and negative controls were included for each experiment.
Real-time analysis of gene expression
The relative abundance of Oct4, Sox2, and Nanog were analyzed using quantitative real-time RT-PCR as described previously (Jafarai et al., 2011; Jafarpour et al., 2011). For each cDNA sample, three replicates of PCR were carried out. The expression levels of the interested genes were compared with human fibroblast. For cyclin-B analysis, intact (MII), enucleated, and reconstituted oocytes at 0.5, 2, 3, 4, and 6 h postnuclear transfer were used (n=15). Primer sequences, annealing temperatures, and the size of amplified products are shown in Table 1.
ESC culture
The process of ESC culture was as described previously (Baharvand and Matthaei, 2004). In brief, iSCNT blastocysts were cultured over mouse embryonic fibroblast feeder cells in knockout DMEM supplemented with 15% knockout fetal calf serum, 2 mM L-glutamine, 0.1 mM nonessential amino acids, 1000 IU/mL leukemia inhibitory factor (Chemicon, Germany), and 10 ng/mL basic fibroblast growth factor at 37°C, 5% CO2, and maximum humidity. Blastocysts were allowed to attach to the feeder layer; then, trophectoderm was dissected out and dome-like formed ICM passaged on fresh feeder. The developed colonies were checked for alkaline phosphatase activity according to manufactures' protocol (Sigma-Aldrich, 85L3R).
Human oocyte and embryos
For comparative gene expression analysis, seven matured oocytes from two patients whom couples had no sperm after ICSI-TESE were used with their permission. These oocytes were inseminated with donor sperm and four blastocysts were obtained. Moreover, OPU-derived oocytes without polar body were gathered from those ICSI-patients who had adequate number of 2PN zygotes for transfer and vitrication. These donated oocytes were cultured for in vitro maturation and oocytes that formed polar body were used.
Statistical analysis
During this study, additional iSCNT trials were performed to provide adequate number of cloned blastocysts for (1) real-time assessment of gene expression, (2) mtDNA analysis, and (3) karyotyping. Each experiment was replicated as least three times, except for real-time analysis of gene expression, which was repeated two times, due to limited number of ICSI-derived human blastocysts. Percentages data were modeled to the binomial model of parameters by ArcSin transformation and the transformed data were analyzed by one-way analysis of variuance (ANOVA) model of SPSS-17. Differences were compared by Tukey multiple comparison post hoc test. All data were presented as means±SEM, and differences were considered as significant at p<0.05.
Results
Nuclear remodeling and MPF activity
As shown in Figure 1B–I and 2A–C, until 2 h postreconstitution, almost all of the cells introduced into the oocytes had compact nuclei (appeared in dark blue) with heavily red-stained plasma membrane and no sign of spindle formation. Three hours postreconstitution the majority of the cells were enlarged about twofold, the plasma membrane weakly emitted red signals, the nuclei were swollen with condensing chromosome, and the newly developed spindle was elongating. At 4 h postinjection, the red signal was non/barely distinguishable; the nucleus was completely swollen with distinguished chromosomes aligning on well-structured bipolar spindle. Further incubation until 6 h did not changed PCC and cell swelling, but increased the percentages of reconstituted oocytes containing chromosomes haphazardly dispersed on multipolar spindles. These events were regardless of the type (fibroblast vs. cumulus) and species (human vs. ovine) of donor cells used. Enucleation and reconstitution did not dramatically decreased MPF activity. There was a time-ependent decline in MPF activity that was more prominent beyond 4-h postreconstitution (Fig. 2D).
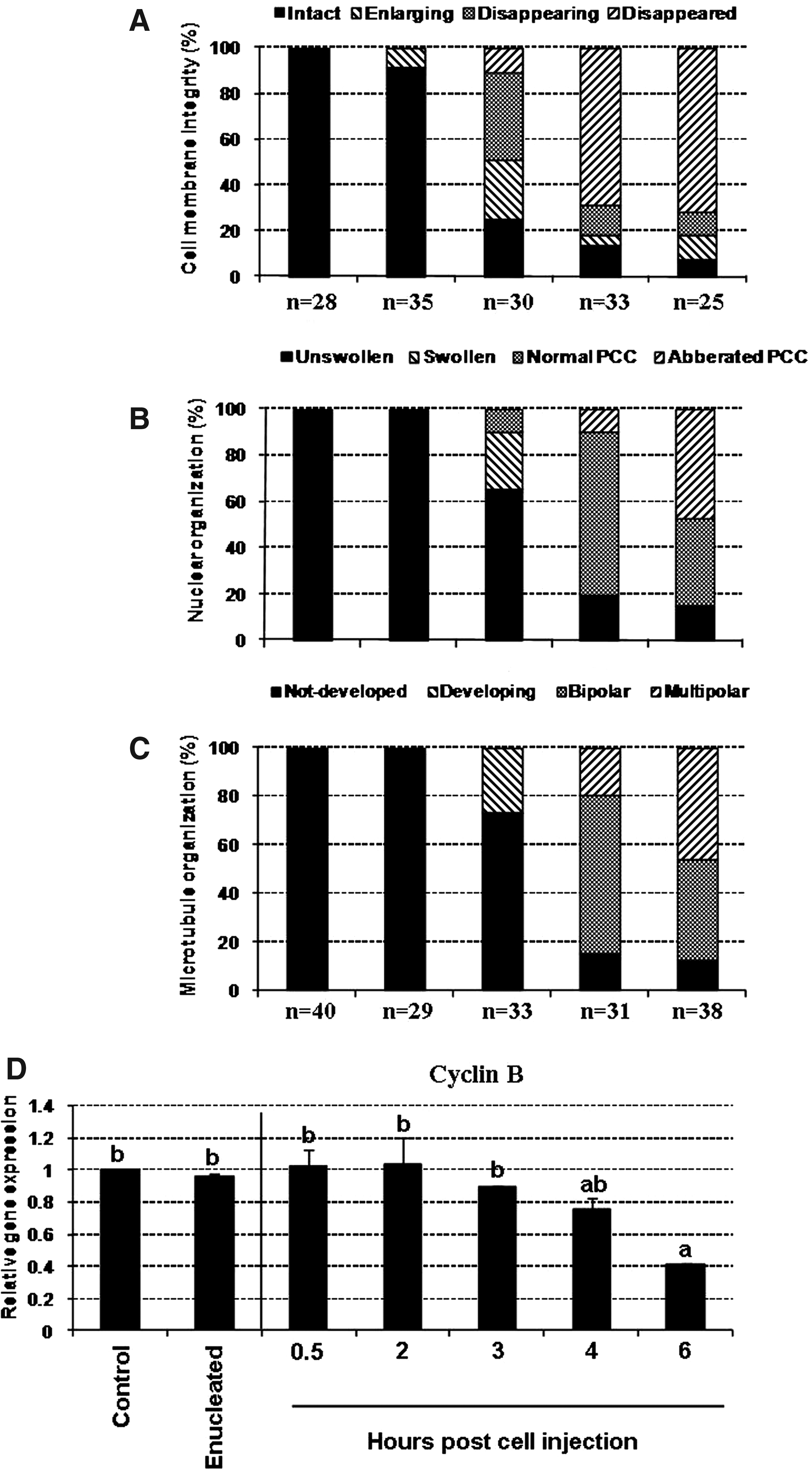
Summary of time-dependent changes occurred in nuclear remodeling and cyclin-B activity of the reconstituted oocytes during early hours post-iSCNT: cell membrane integrity (
Embryo development
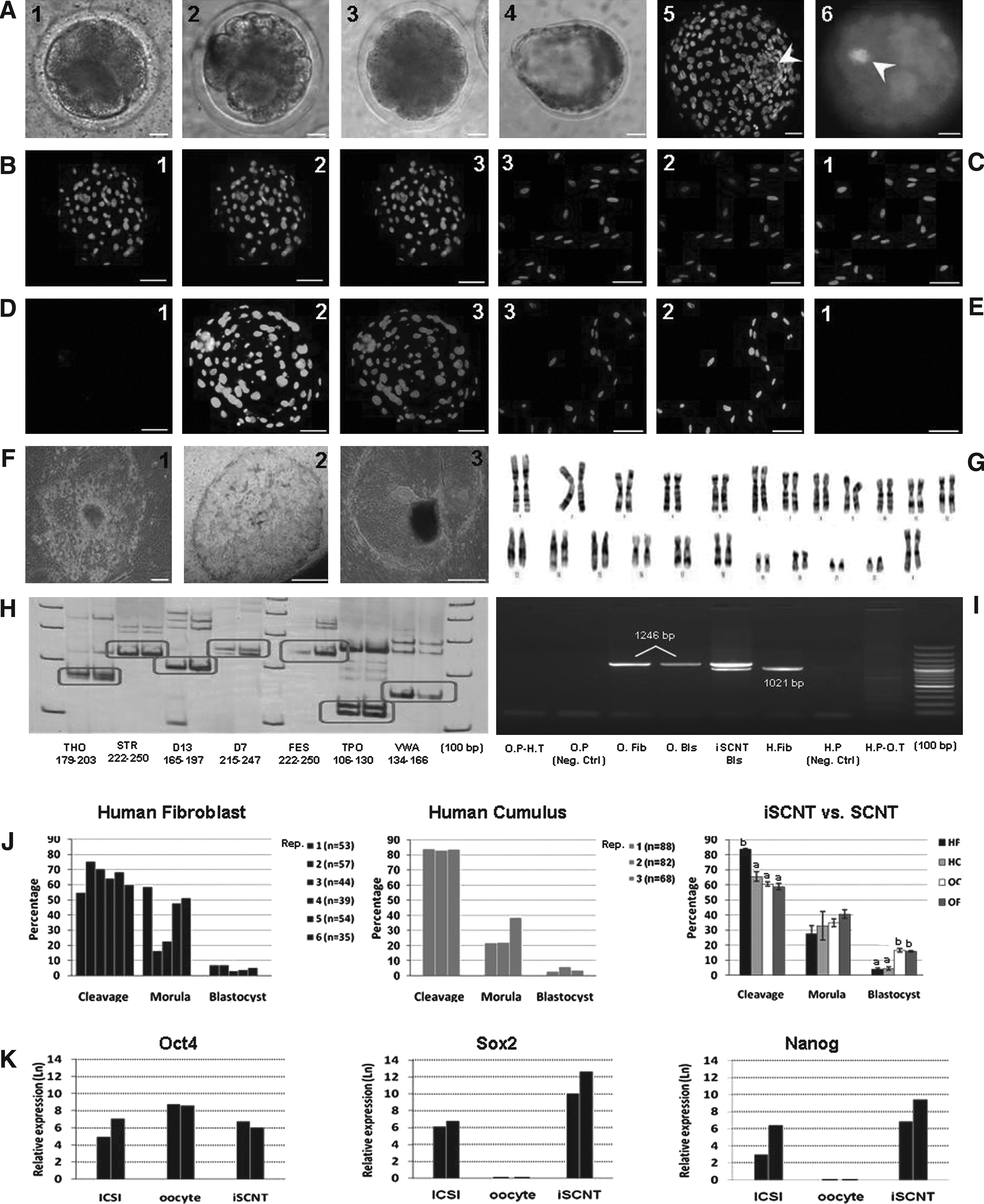
A patient-specific difference in cleavage and in vitro development of the reconstituted iSCNT embryos was observed (Fig. 3J). With pulled data, the cleavage rate of HC reconstructs was significantly lower than HF (65.5±3.01 vs. 83.6±0.31%). The majority of the cleaved embryos arrested at the 8–16-cell stage. H33342-staining indicated that some of the arrested embryos were fragmented, that is, contained blastomere-like structures without nucleus (Fig. 3A). Blastocoel cavity formation was infrequently observed at days 7 and 8, without significant difference between them (HC: 4.4±1.1 vs. HF: 4.0±1.0 %). Morphologically, iSCNT embryos were similar to SCNT embryos. The majority of the developed blastocysts contained distinct ICM and trophectoderm cells (Fig. 3A). The blastocyst rates of ovine cloned embryos (cumulus: 16.6±1.3 vs. fibroblast: 15.9±0.5%) were significantly higher than iSCNT embryos (Fig. 3J).
(
Clone verification
All analyzed iSCNT blastocysts (n=4), had nuclei that positively stained with mouse antihuman nuclear antigen. Positive-stained nuclei were also observed only in human cells, but not ovine cells/embryos (Fig. 3B–E). From four iSCNT embryos used for microsatellite analysis, one embryo failed to amplify and gave no reliable data. The microsatellites of the remained iSCNT embryos were identical to the corresponding donor cells (Fig. 3H).
Genetic and cytogenetic analyses
Figure 3G shows successful karyotyping of iSCNT embryos containing 46 pairs of autosome and sex chromosome of XX. Moreover, Figure 3I shows PCR analysis of human and ovine mtDNA in iSCNT and SCNT blastocysts and their negative and positive controls. As shown, iSCNT blastocysts contained both human and ovine mitochondria.
Real-time analysis of gene expression
For real-time analysis of gene expression, a new set of iSCNT experiments performed to provide adequate number of iSCNT blastocysts. However, because a minimum two blastocysts and 10 oocytes were required to provide an adequate amount of RNA for assessment of the three genes, only two replicates of real-time RT-PCR were carried out using human oocytes (n=20) and ICSI-derived blastocysts (n=4). As shown in Figure 3K, Human oocytes expressed Oct4 transcripts, but failed to express Sox2 and Nanog transcripts. Although very limited expressions of Oct4, Sox2, and Nanog genes were detected in human somatic cells, their expressions were upregulated in iSCNT embryos, at rates comparable to ICSI-derived blastocysts. Ovine oocytes failed to react with human primers.
ESC culture
From six iSCNT blastocysts cultured over the feeder, five attached. Isolated ICMs on the feeder layer were flat with some lipid droplets in the cytoplasm. These colonies sustained their morphology until the third passages when the undifferentiated state did not continue. Importantly, alkaline phosphatase activity was positive only for the first passage (Fig. 3F).
Discussion
Ovine oocyte supports human somatic cell remodeling
During the first hours following SCNT, the transferred nucleus undergoes dramatic structural changes called remodeling (Latham, 2005). Given the regulatory role of subnuclear compartmentalization on gene expression, nuclear remodeling significantly impacts transcriptional status of the cloned embryos (Prunske et al., 2006). Accordingly, the results of this study demonstrated that ovine oocyte was capable of remodeling human somatic cells. The sequence and duration of remodeling events were regardless of somatic cell type used, suggesting their similar reprogrammability with ovine remodeling factors. Although the duration of remodeling events in this study was similar to what was described in porcine (Lee et al., 2003), Byrne et al. (2003) demonstrated that injection of streptolysin-O-permeabilized mouse cells into the Xenopus MII-oocyte resulted in 100-fold increase in nuclear volume within 1 h. Moreover, Kwon et al. (2010) showed that microtubule organization of bovine nuclear transferred embryos occurred within 15 min postfusion. This difference in the duration of remodeling events may be a reflection of different SCNT methods used in these studies. For instance, by simultaneous comparison between two SCNT methods (electrofusion and intracytoplasmic injection), Nagashima et al. (2003) demonstrated clear differences in the pattern of nuclear remodeling and development of the reconstructed embryos.
Importantly, this study showed that prolonged exposure of chromatin (beyond 4 h) to MII-ooplasm adversely affected chromosomes and spindle organization of the reconstituted oocytes. These alterations were concomitant with alterations occurred in MPF activity. The regulatory role of MPF on nuclear organization was also illustrated with a bovine SCNT study in which the percentages of abnormal microtubule organization in cloned embryos treated with roscovitine, MPF activity inhibitor, was significantly higher than those treated with caffeine, the of the MPF activity enhancer (Kwon et al., 2010). Therefore, in this study, reconstituted oocytes were rested for up to 4 h before chemical activation.
Ovine oocyte supports human somatic cell reprogramming
EGA is a nuclear reprogramming event that transforms the genome from transcriptional quiescence at fertilization/SCNT to robust transcriptional activity shortly thereafter (Bultman et al., 2006). Therefore, SCNT embryo development beyond EGA is considered as a widely accepted hallmark of successful reprogramming (Bultman et al., 2006; Chung et al., 2010). EGA occurs around the two-third cell cycle in human embryos (Bultman et al., 2006). Accordingly, the results of this study indicated that ovine enucleated MII-oocyte could successfully reprogram differentiated human cells and compel them back to the embryonic stage. The overall ratios of cleavage and embryo development up to the blastocyst stage, and gene expression analysis further confirmed this issue. However, the percentage of human iSCNT embryos developed to the blastocyst was significantly lower than ovine SCNT. This rate of human iSCNT blastocyst production is comparable with the related rate reported in human–bovine iSCNT (Illmensee et al., 2006), but is far below than human–rabbit iSCNT (Chen et al., 2003) embryos. In contrast, by injecting human somatic cells into bovine, rabbit, mouse, and even human oocytes, Chung et al. (2010) could not find embryo development beyond the EGA stage. These contrasting results were also observed in primate iSCNT studies even using the oocytes of a certain animal species. For example, although chimpanzee/bovine iSCNT embryos did not progress beyond 8–6-cell stage in the study of Wang et al. (2009), Dominko et al. (1999) demonstrated that bovine oocyte supported monkey, porcine, sheep, and rat iSCNT embryos to progress beyond the transcription-requiring, species-specific developmental stages, and formed blastocysts with distinct ICM and trophectoderm. Although the exact explanation for this controversy was not understood, Mitalipov et al. (2007) demonstrated that there is a link between SCNT methodology and nuclear remodeling that is instrumental to successful reprogramming. They suggested that species-specific challenges in SCNT outcome can be overcome by protocol alterations. Interestingly, Zhou et al. (2006) showed that development of a one-step manipulation (OSM) technique in which donor cell was first introduced before oocyte enucleation, for the first time, resulted in routine production of SCNT blastocysts, most of which appear normal by immunochemical, cytochemical, and in vitro developmental criteria. Also, Wrenzycki et al. (2001) showed that the protocol used for SCNT in terms of fusion-activation timing affects pattern of gene expression in cloned cattle embryos. In another study, it was demonstrated that two electrical pulses used to fuse the donor cells with MII-cytoplasts were sufficient to decrease the remaining MPF and prevent NEBD and PCC from occurring (Dominko et al., 1999), which is in agreement with the study of Wakayama et al. (1998).
Ovine oocyte resets human somatic cell pattern of gene expression
Transformation of a terminally differentiated somatic cell to embryonic stage requires a global resetting of gene expression pattern of donor nucleus occurring correctly within a very short period of time (Chung et al., 2010). Among these, Oct4, Sox2, and Nanog are the most important hallmarks of reprogramming and pluripotency, and therefore, their differential expression in cloned embryos is considered as proof of principle for successful reprogramming (Chung et al., 2010; Wang et al., 2009). Accordingly, this study showed that trace transcripts of Oct4, Nanong, and Sox2 were present in somatic cells. Similarly, Page et al. (2009) observed that primary adult human fibroblasts have basal expression of mRNA for Oct4, Nanong, and Sox2, although they suggested that “translation of these messages into detectable proteins and their subcellular localization depends on cell culture conditions.” Moreover, human oocytes expressed Oct4 transcripts, but failed to express Sox2 and Nanog transcripts, which is in agreement with the report of Hansis et al. (2001). Importantly, active transcriptions of the three genes was detected in human iSCNT blastocysts,which were comparable to ICSI-derived blastocysts. Therefore, this result can be considered as proof of principle for the capacity of ovine oocytes in induction of pluripotency in human somatic cells. In contrast, Chung et al. (2010) found that although human oocytes significantly upregulated Oct-4, Sox-2, and Nanog, bovine and rabbit oocytes either showed no difference or downregulation of these genes in human–somatic cell-cloned embryos. Although the exact reason of this controversy was not understood, a comparison between reprogramming ability of ovine and bovine oocytes in a study and assessment of a global pattern of gene expression are needed to provide a final conclusion about the mechanism of pluripotency induction in interspecies SCNT studies. Even though early evidences of morphology and immunostaining indicated the undifferentiated status of isolated and cultured ICM cells, subsequent cultures failed to produce ESC. Because there is no available report about the establishment of ovine ESC, it seems that the procedure of ovine ESC culture has yet to be achieved. It implies that there is a place for further studies to increase the chance of ESC derivation from human cloned embryos.
Footnotes
Acknowledgments
This study was funded by the grant of Royan Institute. The author gratefully thanks Mrs. Mansouri and Mr. Shdanloo for statistical analysis of data, and Mr. Heidari and Khajo for ovary preparation.
Author Contributions
H.S.M. and N.M.H. conceived and designed the experiments. H.S.M., H.M., F.M., M.F., A.P., O.S., K.F., K.K.h., B.H., A.H., A.V., and J.F. performed the experiments. H.S.M. and N.M.H. analyzed the data. H.S.M. and N.M.H. wrote the article.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.