
Abstract
Abstract
It is generally accepted that mammalian females are born with a finite pool of oocytes and that this is the sole source of ovules throughout the reproductive life of the adult. This dogma was shaken in 2003 when researchers showed that the oocyte stock might be renewable in adult mammals. It has been proposed that hematopoietic stem cells might be a source of new oocytes. These discoveries have puzzled many researchers and remain controversial. In our study, we attempted to determine if transplanted bone marrow cells could provide new oocytes in PU.1 mice and in severe combined immunideficiency (SCID) mice after treatment with chemotherapeutic agents. We also examined the possibility that grafted bovine embryonic ovarian cortex might provide an environment favoring such a response. We found no evidence that transplanted bone marrow cells provide new fertilizable oocytes in PU.1 mice, in SCID mice treated with chemotherapeutic agents, or with bovine embryonic ovarian tissue grafts. However, transplanted bone marrow cells have improved the fertility of SCID mice previously treated with chemotherapeutic agents. These data suggest that bone marrow cells cannot provide new oocytes but can positively influence ovarian physiology to improve the fertility of mice previously treated with chemotherapeutic agents.
Introduction
Materials and Methods
Animals
Female SCID (CB17/Icr-Prkdc(scid)/IcrIcoCrl) mice aged 21 days were purchased from Charles River laboratories. PU.1-heterozygous mice were obtained from Dr. McKercher (Burnham Institute for Medical Research, Del E. Webb Center for Neuroscience and Aging, La Jolla, CA). Generation and genotyping of PU.1-null mice has been described previously (McKercher et al., 1996). The purine-rich box-1 is a transcription factor that belongs to the ETS (E-twenty six) family. It is a key regulator gene of the hematopoietic system (Scott et al., 1994) and is essential for myeloid differentiation, development of B and T cells, erythropoiesis, and hematopoietic stem cell maintenance (Kastner and Chan, 2008). Without daily injections of antibiotic, PU.1-null mice die from septicemia within 48 h. With the injections, they can survive 20 days (Anderson et al., 1998). To survive for more than 20 days, they need a bone marrow transplant within 48 h after birth (Mezey et al., 2000). Mice expressing the protein GFP (C57BL/6-Tg(ACTB-EGFP)1Osb/J) were obtained from Jackson Laboratories. GFP are transgenic for GFP and are homozygous for wild-type PU.1 (PU.1+/+). All animal work was conducted using procedures reviewed and approved by the Université Laval animal care committee (authorization number: 2006036-1).
Treatment with chemotherapeutic agents
Chemotherapeutic agents were used to destroy oocytes in mouse ovaries (Johnson et al., 2004; Shiromizu et al., 1984) and in fetal bovine ovarian cortex in order to assess the possibility of subsequent bone marrow transplant rescuing fertility (Fu et al., 2008; Lee et al., 2007; Oktay and Oktem, 2007) or leading to the production of new oocytes (Johnson et al., 2005) in SCID mice or in fetal bovine ovary grafts in SCID mice. The treatment consisted of a single intraperitoneal injection of cyclophosphamide 120 mg/kg (Cytoxan; Brystol-Myers, Plainsboro, NJ) and Busulfan 12 mg/kg (Busulfan; Sigma, St. Louis, MO) in 26-day-old female SCID mice. For the bovine xenograft experiment, 54-day-old female SCID mice were given the treatment.
Recovery of bone marrow cells
GFP-expressing female donor mice between 1 and 3 months old were sacrificed and their femurs removed under sterile conditions. Using a 27-gauge needle, bone marrow was flushed from the femurs with Dulbecco's phosphate-buffered saline (DPBS) modified with 1000 mg/L glucose, 0.036 g/L sodium pyruvate, 0.05 g/L streptomycin sulphate, 0.1 g/L kanamycin monosulphate, 0.133 g/L CaCl2 and 0.1 g/L MgCl2, liquid, sterile-filtered and cell culture-tested (Sigma catalog number D4031) supplemented with 2% fetal bovine serum (FBS). This suspension was then filtered through 40-Am nylon mesh (Becton-Dickinson, Franklin Lakes, NJ, 40-μm nylon cell strainer) and centrifuged (2000×g for 10 min at 4°C). The pellet was resuspended in FBS-free DPBS and the cells therein were counted. Cells were then recentrifuged and a solution of 2×107 cells/mL in sterile saline solution was made for transplants. In the PU.1-null mice protocol, cells from one GFP femur were diluted in 20 μL of sterile saline solution.
Experimental strategy (Fig. 1)
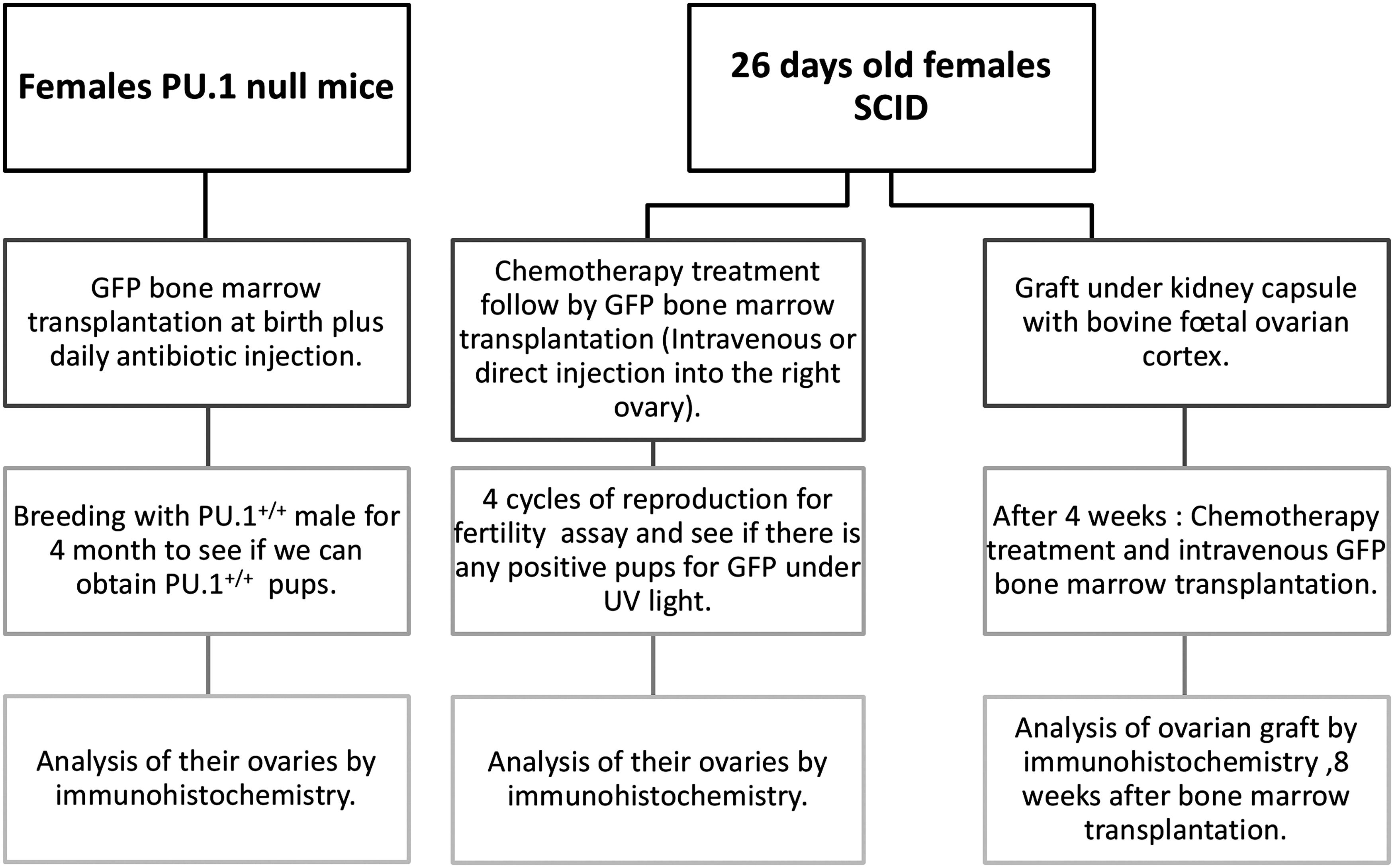
Schematic representation of the experimental protocol.
The first strategy was to assess whether transplanted bone marrow cells could produce new oocytes in the PU.1-null mouse model. PU.1-null mice require bone marrow transplantation at birth in order to survive. We thus sought to generate GFP-positive oocytes. PU.1+/− mice were bred to produce PU.1−/− (null) mice. At birth and daily thereafter, litters were injected with antibiotic, 2.5 mg/kg (Baytril, Bayer HealthCare, Animal Health Division, Bayer Inc. Toronto). Tail-tip blood smears were stained with Wright Giemsa for rapid identification of null individuals by absence of neutrophils (McKercher et al., 1996). Null mice then received bone marrow cells (from female GFP, PU.1+/+ wild-type) by intraperitoneal injection. Bone marrow-transplanted PU.1−/− females that survived until adulthood were mated with PU.1+/+ males to test for the presence of PU.1+ oocytes, appearing as PU.1+/+ homozygous pups. The genotyping of PU.1 pups from this mating was performed as described previously (McKercher et al., 1996).
The second strategy was to determine whether GFP bone marrow could replace the loss of oocytes due to chemotherapeutic agents in female SCID mice. Female SCID mice 26 days old were treated as described above and given an intravenous injection of 1×107 female GFP bone marrow cells the next day.
The third strategy was to inject GFP bone marrow directly into the right ovary of female SCID mice the day after treatment with chemotherapeutic agents. The right ovary was exposed surgically and about 1 μL of GFP bone marrow solution (2×104 cells) was injected. One week after the surgery, these SCID females were mated with SCID males (see SCID fertility assay below).
The fourth strategy was to assess whether transplanted bone marrow might be able to provide new oocytes in grafted tissue from a different species such as cattle. Fetal bovine ovarian cortex was thus grafted under the kidney capsule of 26-day-old female SCID mice. Ovaries were removed from bovine fetuses 30 to 70 cm in length obtained from a local slaughterhouse. About 1 mm3 of cortex was grafted. Four weeks later, the mice were treated with chemotherapeutic agents (as described above) to destroy the oocytes present in the graft and given an intravenous GFP bone marrow transplant (1×107) the following day. Eight weeks after the bone marrow transplant, the bovine ovarian xenografts were recovered for immunohistological analysis.
Fertility assay
The day after receiving the bone marrow transplant, female SCID mice were mated with SCID males. The number of pups in each litter was noted and the day-old pups were placed under UV light to detect GFP and then sacrificed. The SCID females were mated repeatedly with SCID males and sacrificed after a minimum of four completed pregnancies. Ovaries were recovered for immunohistological analysis. PU.1-null mice that survived until adulthood were mated with wild-type PU.1 males to determine if they could produce homozygous wild-type progeny. The fertility of transplanted PU.1-null mice was likely affected by other factors such as a major decrease in growth rate, either due to or exacerbated by malformation and protrusion of the teeth in odd directions. These mice needed soft food, were very docile and displayed weak maternal behavior.
Immunohistological analysis
Each ovarian sample was fixed overnight at 4°C in neutral-buffered 10% formalin solution and embedded in paraffin. Tissue sections 8 μm thick were sliced from samples and mounted on slides, dewaxed in toluene, rehydrated, and boiled for 20 min in Tris/EDTA pH 9.0. Sections were blocked for 20 min using 10% goat serum in Tris-buffered saline, followed by incubation overnight at 4°C with rabbit antimouse-MSY2 (kindly provided by Richard M. Schultz, Department of Biology, University of Pennsylvania, Philadelphia, PA) and chicken anti-GFP (Abcam; ab13970) diluted respectively 1:4000 and 1:500 in Tris-buffered saline. The sections were blocked again the following day with the Tris-buffered goat serum solution for 20 min and incubated with goat antichicken-Ig conjugated with Alexa Fluor® 488 (Invitrogen Alexa Fluor®; A-11039) and goat antirabbit-Ig conjugated with Alexa Fluor® 568 (Invitrogen Alexa Fluor®; A-11036), both diluted 1:200 in Tris-buffered saline. Sections were then mounted with ProLong® Gold anti-fade reagent with DAPI (Invitrogen; P36935) and slides were analyzed with a Nikon Eclipse E600 fluorescence microscope.
Statistical Analysis
Statistical analyses were conducted using Prism 5.00 GraphPad for Windows (GraphPad Software Inc., San Diego, CA; www.graphpad.com). Statistical significance was assessed by ANOVA followed by Bonferoni's multiple comparison tests to identify individual differences between treatments. Probabilities of p<0.05 were considered statistically significant. All values are presented with their corresponding SEM. Different letters represent statistical differences between treatments.
Results
GFP bone marrow transplants in PU.1-null (PU.1−/−) mice
By transplanting hours-old PU.1-null females with female GFP bone marrow, we expected to determine whether bone marrow cells could migrate to the ovary and produce new fertilizable oocytes and thus GFP-positive progeny. Because a cross between PU.1−/− and PU.1+/+ should produce only heterozygous (PU.1+/−) pups, the presence of homozygous PU.1+/+ pups in the progeny would indicate that transplanted bone marrow cells became oocytes. Despite nearly 20 attempts, we obtained only 2 litters of two pups each from two different females. None of the pups were homozygous for wild-type PU.1 (Fig. 2), indicating that they came from endogenous oocytes and not transplanted GFP bone marrow cells that have the wild-type version of PU.1 (PU.1+/+). Immunohistological analyses of the six ovaries from three PU.1 transplanted mice were done in order to check for the presence of the specific markers Y-box protein MSY2 (expressed exclusively in male and female germ cells) and GFP in oocytes, which would indicate neo-oogenesis from bone marrow transplants. No GFP-positive oocytes were found in any of the 900 tissue sections (150 per ovary) thus examined (Fig. 3A). These results therefore do not provide evidence of new fertilizable oocytes in the PU.1-null mouse model.

Genotyping of four pups from female PU.1-null mice transplanted with GFP bone marrow crossed with wild-type PU.1 male. Lanes 2 and 3—heterozygous pups from the first PU.1-null female; lanes 4 and 5—heterozygous pups from the other PU.1-null female; lanes 1 and 7—DNA mass ladder; lane 6—blank. Wild-type PU.1 corresponds to the upper band (1170 bp) and PU.1-null corresponds to the lower band (980 bp).
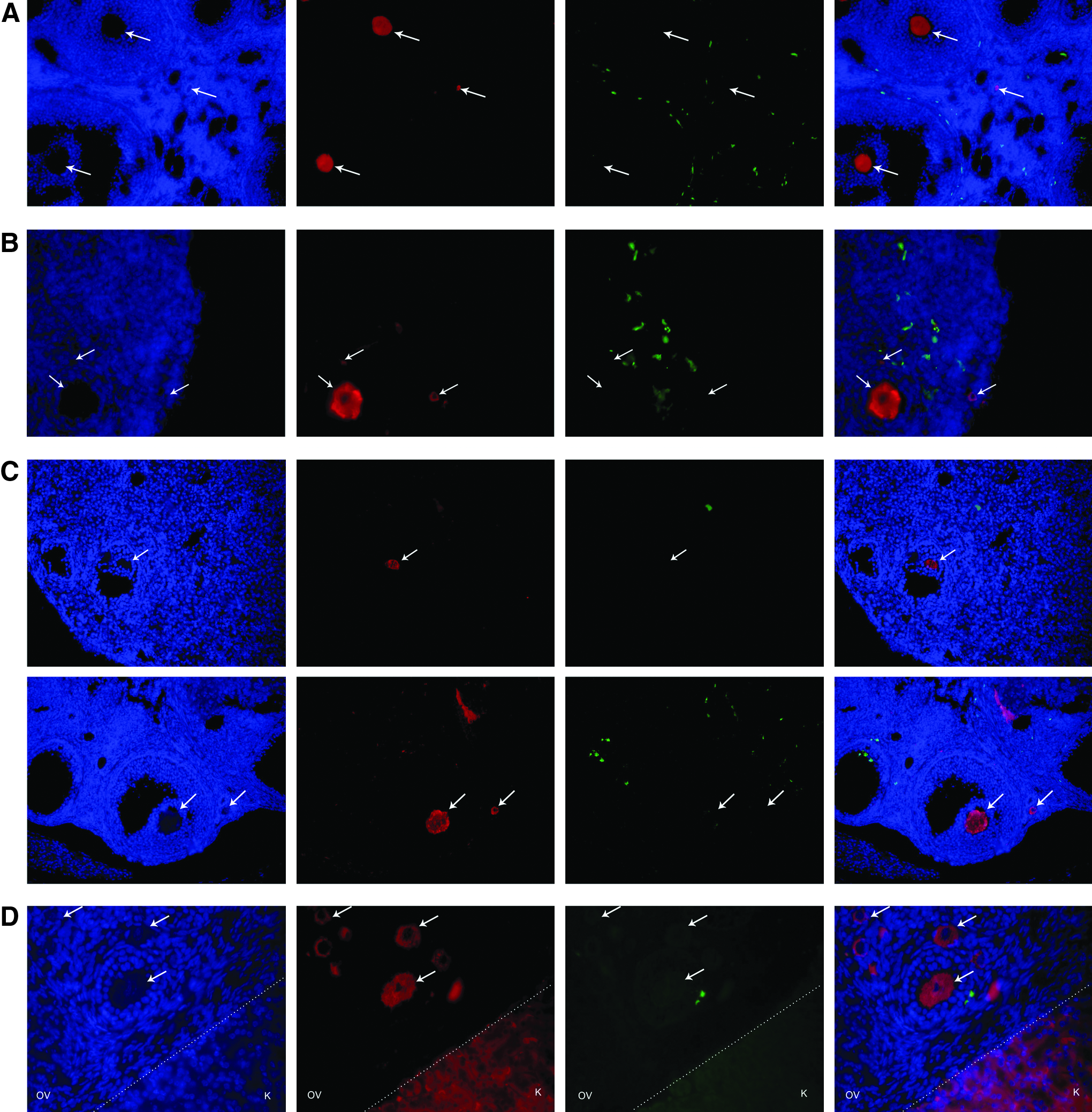
Representative dual immunofluorescence images of ovarian sections (
Bone marrow transplant in female transgenic SCID mice
To assess the likelihood of bone marrow cells colonizing mice ovaries and producing fertilizable oocytes, we treated female SCID mice with the chemotherapeutic agents to destroy most of the native pre- and postmeiotic germ cell pools in their ovaries (Johnson et al., 2004; Plowchalk and Mattison, 1992; Shiromizu et al., 1984). The following day, the mice received an intravenous injection (i.v.) or a direct injection into the right ovary (ROV) of female GFP bone marrow. They were later mated with SCID males to determine if they could give rise to GFP progeny. None of the pups were positive for GFP and therefore did not come from the transplanted cells. The number of pups per litter was noted. Control mice not given chemotherapeutic agents had about six pups per litter, whereas chemotherapeutic-treated mice without GFP bone marrow transplant had four pups per litter. The ROV mice had on average five pups per litter. However, this increase was not significantly different from chemotherapeutic-treated mice not given bone marrow. The chemotherapeutic-treated IV group had on average six pups per litter, the same as the control group (Fig. 4). These results do not support the hypothesis that bone marrow transplants can lead to the production of new fertilizable oocytes, but do suggest that they may improve the fertility of SCID mice treated with chemotherapeutic agents. We also examined SCID ovaries for the presence of GFP-positive oocytes using antibody raised against the oocyte-specific Y-box protein MSY2 as well as antibody raised against the GFP protein. Colocalization of these two proteins within an oocyte would indicate that it was derived from transplanted bone marrow cells and not the original SCID ovarian stock. Both ovaries from SCID mice treated with the chemotherapeutic agents and given GFP-expressing bone marrow cells intravenously or by direct transplant into the right ovary were analyzed using this immunohistological procedure. No cell positive for both MSY2 and GFP was found, and hence, no support for the claim that bone marrow cells can become oocytes (Fig. 3B). Furthermore, GFP-positive cells were found in both the right and left ovaries of the mice that received the surgical transplant and no significant difference was observed between the ovaries. However, there were usually fewer GFP-positive cells in these ovaries than in those of the mice that received the intravenous transplant (Fig. 3C). These results thus do not provide any support for the hypothesis that transplanted bone marrow cells can provide new oocytes in SCID mice treated with chemotherapeutic agents.
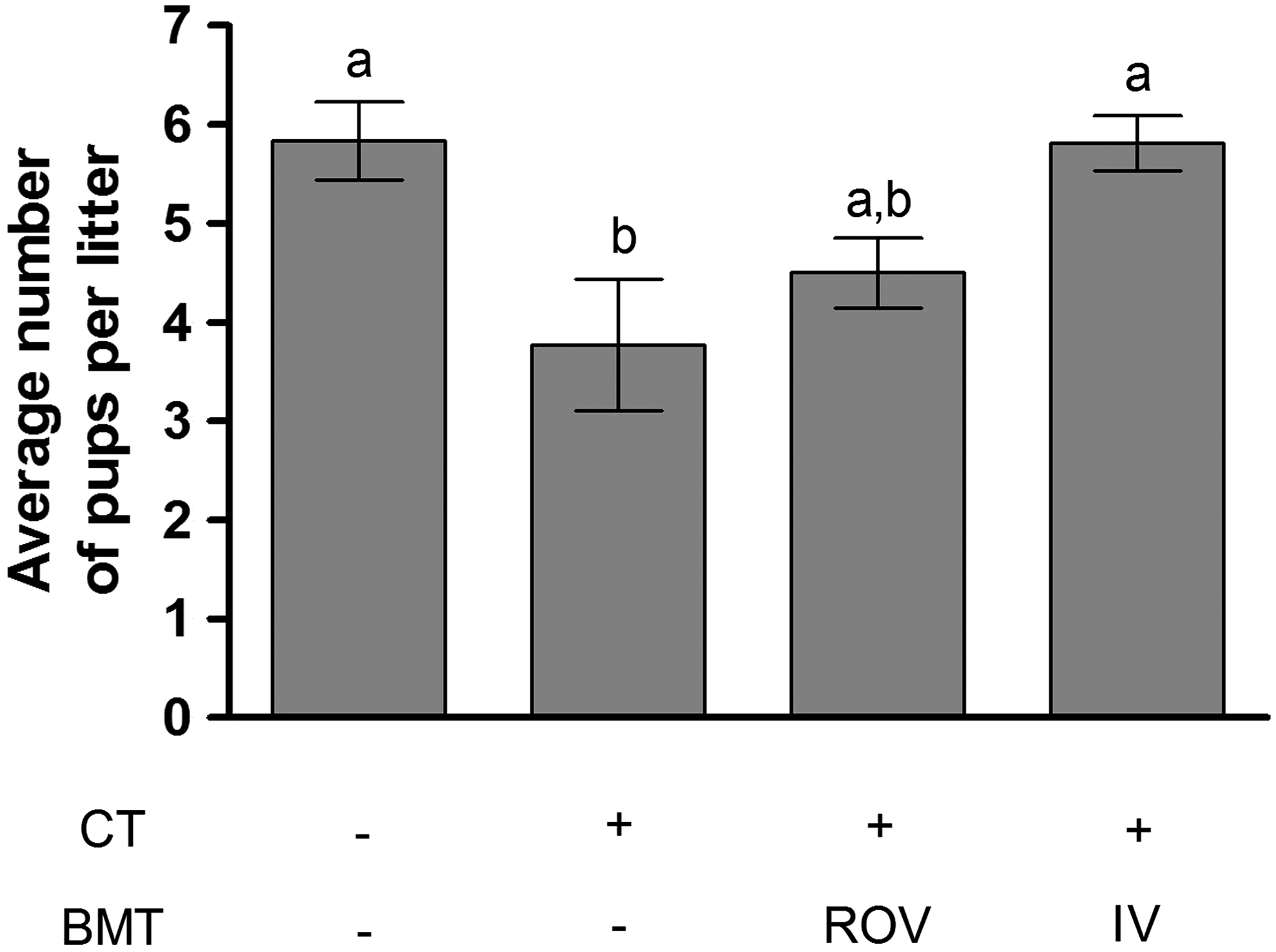
Effect of bone marrow transplants on the fertility of female SCID mice (four mice per treatment with a minimum of four completed pregnancies per mice). CT—single injection of cyclophosphamide (120 mg/kg) and Busulfan (12 mg/kg); BMT—transplant of bone marrow cells [from female mice expressing GFP) in the right ovary (ROV) or by intravenous injection (i.v.)]. Data represent the mean±SEM. Statistical analysis was performed using ANOVA (p<0.05). Differing superscripts indicate significant differences based on Bonferroni post tests.
GFP-expressing bone marrow cells in bovine fetal ovarian cortex grafts in SCID mice
To assess the likelihood of mouse bone marrow cells evolving into new oocytes in ovarian tissues of other mammals such as cattle, about 1 mm3 of bovine fetal ovarian cortex was grafted under the kidney capsule female SCID mice. Three weeks after the surgery, the mice were treated with chemotherapeutic agents to destroy the oocytes within their ovaries as well as most bovine oocytes that have remained in the graft. The following day, the mice received GFP-expressing bone marrow cells by intravenous injection. Two months later, the presence of GFP-positive oocytes within three bovine grafts was checked using the immunohistological technique already described, targeting the oocyte-specific marker Y-box protein MSY2 and GFP (Fig. 3D). No colocalization of GFP and MSY2 was observed, and hence, no support for the hypothesis that an ovarian tissue xenograft might enable transplanted bone marrow cells to provide new oocytes in SCID mice.
Discussion
Neo-oogenesis in the postnatal mammalian ovary has been the subject of heated debate in the fields of reproductive and developmental biology. Two studies have suggested that bone marrow cells are potential contributors to this phenomenon (Johnson et al., 2005; Lee et al., 2007), whereas others have been unable to validate this hypothesis (Begum et al., 2008; Eggan et al., 2006), as was the case in the present study. In none of the models that we investigated (PU.1-null mice, SCID mice treated with chemotherapeutic agents or bovine fetal ovarian cortex grafted in SCID mice thus treated) could oocytes originating from transplanted bone marrow cells be detected. It has been reported that transplanted bone marrow cells may be involved in ovarian recovery and improvement of female fertility after chemotherapy (Fu et al., 2008; Lee et al., 2007; Liu et al., 2007) and our results appear to corroborate this finding. Indeed, bone marrow transplantation has not leaded to the production of new fertilizable oocytes, but may have improved the fertility of SCID mice treated with chemotherapeutic agents. Unfortunately, the fertility of transplanted PU.1-null mice was likely affected by other factors such as a major decrease in growth rate, either due to or exacerbated by malformation and protrusion of the teeth in odd directions. These mice needed soft food, were very docile and displayed weak maternal behavior. Two interpretations have been proposed in the literature to explain improvement of female fertility after chemotherapy. Lee et al. (2007) proposed that the rescue of mouse fertility by transplanted bone marrow cells after treatment with chemotherapeutic agents confirms the existence of germ-line stem cells in bone marrow, whereas a recent review article proposes the involvement of the immune system, via autoimmune response in the ovary following chemotherapeutic treatment (Notarianni, 2011). According to the latter hypothesis, chemotherapeutic agents induce ovarian failure, cellular apoptosis and depletion of ovarian antigen-specific regulatory T cells, thereby promoting activation of effector T cells and inducing autoimmune response to ovarian antigens, leading to clearance of oocytes that survived exposure to chemotherapeutic agents. It is known that bone marrow transplants attenuate autoimmune responses induced by chemotherapy and they might do this by providing regulatory T cells in the ovary to promote resumption of self-tolerance. Follicles that survive chemotherapy could thus grow and reach ovulation rather than being cleared, as they apparently are in mice receiving chemotherapeutic agents but not bone marrow cells. This might explain the increased number of pups observed in litters from mice treated with chemotherapeutic agents and bone marrow cells. In conclusion, this study suggests that the beneficial effects of transplanted bone marrow cells on the fertility of female mice could be due to restoration of self tolerance of ovarian antigens, thus sparing follicles not destroyed by exposure to chemotherapeutic agents from subsequent autoimmune clearance. However, this explanation remains speculative due to the absence of immunological data. Because female fertility is often seriously compromised after chemotherapy, any findings that suggest the potential of a treatment to restore or protect fertility warrant further investigation. Deeper understanding of this phenomenon may shed light on the true nature of apparent cases of neo-oogenesis.
Footnotes
Acknowledgments
This study was funded by the CRSNG. We gratefully acknowledge Dr. Shultz for providing us the antibody against MSY2 and Dr. McKercher for providing PU.1 transgenic mice.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.