
Abstract
Abstract
Despite extensive efforts, low efficiency is still an issue in bovine somatic cell nuclear transfer (SCNT). The hypothesis of our study was that the use of cytoplasts produced by chemically assisted enucleation (EN) would improve nuclear reprogramming in nuclear transfer (NT)–derived embryos because it results in lower damage and higher cytoplasm content than conventional EN. For that purpose, we investigated the expression of two X-linked genes: X inactive-specific transcript (XIST) and glucose 6-phosphate dehydrogenase (G6PD). In the first experiment, gene expression was assessed in day-7 female blastocysts from embryonic cell NT (ECNT) groups [conventional, ECNT conv; chemically assisted, ECNT deme (demecolcine)]. Whereas in the ECNT conv group, only one embryo (25%; n=4) expressed XIST transcripts, most embryos showed XIST expression (75%; n=4) in the ECNT deme group. However, no significant differences in transcript abundance of XIST and G6PD were found when comparing the embryos from all groups. In a second experiment using somatic cells as nuclear donors, we evaluated gene expression profiles in female SCNT-derived embryos. No significant differences in relative abundance (RA) of XIST transcripts were observed among the groups. Nonetheless, higher (p<0.05) levels of G6PD were observed in SCNT deme and in vitro–derived groups in comparison to SCNT conv. To know whether higher G6PD expression in embryos derived from SCNT chemically assisted EN indicates higher metabolism in embryos considered of superior quality or if the presence of higher reactive oxygen species (ROS) levels generated by the increased oxygen consumption triggers G6PD activation, the expression of genes related to stress response should be investigated in embryos produced by that technique.
Introduction
The success of cloning relies largely on the female gamete. It is known that selection of high-quality recipient oocytes for nuclear transfer increases the cloning efficiency and the number of offspring obtained (Miyoshi et al., 2003). Previous studies (Collas and Robl, 1991; Czolowska, et al., 1984) have demonstrated the effects of host ooplast cell cycle stage on nuclear remodeling and developmental competence of reconstructed embryos. Exchanges of both acidic and basic proteins between donor nuclei and cytoplasm have been observed, and it can be speculated that ooplasmic proteins imported into the nucleus mediate structural rearrangement of the chromatin, which functionally resets it into a totipotent state (for review, see Bordignon et al., 1999, 2001).
The enucleation (EN) process in traditional nuclear transfer (NT) involves ultraviolet (UV) irradiation, which can cause alterations in the membrane and intracellular components of bovine oocytes (Smith, 1993). Moreover, damage caused to the recipient oocyte is aggravated by the concomitant removal of a large volume of cytoplasm surrounding the metaphase plate, which contains mRNA, proteins, and molecular precursors essential to early development until embryonic genome activation (EGA) (Barnes and Eyestone, 1990). An interesting alternative strategy to physical EN is to treat oocytes with agents that modify the processes of karyokinesis and cytokinesis, inducing the formation of a visible protrusion containing condensed chromatin on the oocyte surface, which results in chemically enucleated oocytes at high rates (Kawakami et al., 2003; Li et al., 2004, 2006; Saraiva et al., 2009; Tani et al., 2006; Vajta et al., 2005; Yin et al., 2002).
Nuclear remodeling, which presumably results in its reprogramming, is characterized by a variety of structural changes. It is thought to occur completely and consistently only after nuclear envelope breakdown and chromosome condensation, and it is initiated by a high level of maturation-promoting factor (MPF) (Fulka et al., 1996). It has been reported that MPF activity increased up to 30% in oocytes enucleated by the chemically assisted approach with the use of demecolcine, a microtubule-depolymerizing agent (Li et al., 2009; Tani et al., 2006). Furthermore, the alternative methods for oocyte EN lead to the presence of larger cytoplasmic volume and might allow the conservation of spindle-associated factors in the enucleated cytoplasts (Costa-Borges et al., 2009), which could assist in nuclear reprogramming.
It is known that modifications in the NT protocol have several effects on embryonic gene expression patterns, specifically on stress susceptibility–related genes, trophoblastic function, DNA methylation, and X chromosome inactivation (XCI) (Wrenzycki and Niemann, 2003; Wrenzycki et al., 2001, 2002). Nonetheless, we did not find any reported data that show the effects of alternative methods for oocyte EN on the expression of these essential genes for early embryonic development.
In eutherian mammals, XCI is required to ensure an equal transcriptional level (dosage compensation) for X-linked genes between males and females. Both X chromosomes are active after EGA, and XCI is not fully accomplished during early development, which leads to a double amount of expression of two X-linked genes in female embryos. In bovine species, XCI starts at the early blastocyst stage (De La Fuente et al., 1999), and a recent report (Bermejo-Alvarez et al., 2011) has shown that the process is actually performed between days 7 and 14, i.e., at the early elongation stage. Nonetheless, there is evidence that in vitro production (IVP) systems (Balasubramanian et al., 2007; Lopes et al., 2007; Wrenzycki et al., 2002) and NT procedures (Wrenzycki et al., 2002) can result in changes in the XCI process, altering the dosage compensation of some X-linked genes in embryos obtained by those technologies.
Furthermore, the enzyme glucose-6-phosphate dehydrogenase, encoded by the X-linked gene G6PD, participates in the detoxification of oxygen radicals (Nicol et al., 2000; Peippo and Bredbacka, 1995) and has been considered a cytoprotective enzyme for oxidative stress and DNA damage (Nicol et al., 2000). It has been speculated that higher expression of the G6PD gene in IVP embryos may be an adaptive response to the oxidative stress induced by the in vitro conditions (Lonergan et al., 2003; Lopes et al., 2007). However, the optimal levels of reactive oxygen species (ROS) have not been well defined, and although high ROS levels could cause cell injury, low levels could reduce the expression of genes involved in cell proliferation (Peippo et al., 2001).
Gene expression techniques have recently become a powerful tool for analyzing transcripts related to embryo quality (Warzych et al., 2007). To our knowledge, there are no published data regarding the nuclear reprogramming in NT-derived embryos generated by cytoplasts from alternative EN techniques. The objective of the present study was to verify the relative transcript levels of some essential genes to early developmental stages in reconstructed embryos from chemically assisted EN. For this purpose, initially we investigated the expression of two X-linked genes, XIST and G6PD, by the quantitative real-time PCR (qPCR) procedure, in female bovine embryos produced from cytoplasts originated by conventional and chemically assisted EN, using embryonic blastomeres as nuclear donors. In a second experiment, we evaluated the expression profile of these genes in female embryos reconstructed with somatic cells. Considering the lower damage suffered by cytoplasts derived from chemically assisted EN, the hypothesis of our study was that those structures would contribute to nuclear reprogramming improvement in NT-derived embryos.
Materials and Methods
Chemicals
All chemicals and media were purchased from Sigma-Aldrich Co. (St. Louis, MO, USA), unless otherwise stated.
In vivo embryo production
In vivo–derived (IVD) bovine embryos were produced using the standard superovulation protocol and artificial insemination of Bos taurus×Bos indicus crossbred cows. Briefly, follicular waves were synchronized by dominant follicle aspiration and insertion of a progesterone (P4)-releasing device (Crestar, Intervet Internation GmbH, Unterschleissheim, Germany). In all, 220 mg of follicle-stimulating hormone (FSH; Folltropin™, Bioniche Animal Health, Belleville, Canada) were administered intramuscularly (i.m.) in eight decreasing doses, twice a day, for 4 consecutive days, beginning 3 days after follicular aspiration. To induce luteolysis, 150 μg of prostaglandin F2α analog were administered with the fifth or sixth FSH injection (
In vitro embryo production
Oocyte recovery and in vitro maturation
Bovine oocytes were obtained by follicular aspiration from ovaries obtained at a local slaughterhouse. The ovaries were transported to the laboratory in 0.9% saline solution at 30–35°C. Follicles with diameters between 3 and 8 mm were aspirated using an 18-gauge needle attached to a 20-mL syringe and cumulus–oocyte complexes (COCs) with at least three layers of cumulus cells and homogeneous cytoplasm were selected. The COCs were washed in HEPES-buffered tissue culture medium-199 (TCM-199; Gibco BRL, Grand Island, NY, USA) supplemented with 10% fetal calf serum (FCS) heat-inactivated at 55°C for 30 min, as well as 0.20 mM of sodium pyruvate and 83.4 μg/mL of amikacin (Instituto Biochimico, Rio de Janeiro, Brazil). Groups of 20–25 COCs were matured in droplets (100-μL) of TCM-199 supplemented with 10% FCS, 1.0 μg/mL FSH, 50 μg/mL hCG (Profasi™, Serono, São Paulo, Brazil), 1.0 μg/mL estradiol, 0.20 mM of sodium pyruvate, and 83.4 μg/mL amikacin under mineral oil (Dow Corning Co., Midland, MI, USA).
In vitro fertilization
To produce embryos by IVF, at 24 h postmaturation, groups of 15 expanded COCs were transferred to 30-μL drops of TALP-IVF medium supplemented with 6 mg/mL bovine serum albumin (BSA), 30 μg/mL heparin, 18 μM penicillamine, 10 μM hypotaurine, 1.8 μM epinephrine, and 0.2 mM sodium pyruvate, covered with sterile mineral oil. IVF was performed with X-sorted sperm from the same bull used to obtain IVD embryos. Frozen straws containing approximately 2 million spermatozoa were thawed at 35.5°C and motile spermatozoa were separated by density gradient in 45% and 90% (vol/vol) Percoll (Amersham Pharmacia Biotech AB, Uppsala, Sweden). The final suspension was divided among five drops containing the oocytes, in a final concentration of approximately 104 spermatozoa for each oocyte. Oocytes and sperm cells were co-incubated for 22 h at 38.5°C under 5% CO2 in air and maximum humidity.
Establishment of embryonic nuclei donor cells
Blastomeres derived from in vitro–produced female day-5 morulae were used for NT. For blastomere synchronization at the metaphase stage, the embryos were cultured in synthetic oviductal fluid (SOF) medium in the presence of 0.4 μg/mL demecolcine for 12 h. Approximately 3 h before the beginning of micromanipulation, the embryos were washed several times and cultured in demecolcine-free SOF, so that at the time of use the embryonic cells were in the G1 phase (Oback and Wells, 2002), a stage compatible with metaphase II enucleated cytoplasm. Before micromanipulation, the embryos were exposed to a 0.5% pronase solution for 30 sec to remove the zona pellucida, followed by disaggregation of blastomeres using a fine-bore pipette.
Establishment of somatic nuclei donor cells
Female bovine fibroblasts, obtained from a Nellore (Bos indicus) cow, were used as nucleus donors. Fragments from a skin biopsy were initially cultivated in Dulbecco's modified Eagle medium (DMEM; Gibco BRL) supplemented with 50% FCS and 83.4 μg/mL amikacin at 38.5°C under 5% CO2 in air. After establishment of the primary cell line, the cells were maintained in DMEM supplemented with 10% FCS and amikacin and passaged when they reached confluence. The medium was replaced every 48 h and cells underwent three to five passages (0.05% trypsin with 1% poultry serum) before freezing and storage in liquid nitrogen. Before NT, the cells were cultured in DMEM supplemented with 0.5% FCS for 3–5 days to induce cell quiescence and cell cycle synchronization in stage G0–G1 (Campbell et al., 1996), compatible with metaphase II enucleated cytoplasm.
Oocyte EN, nuclear transfer, and electrofusion
Chemically assisted EN was performed after 19 h of maturation according to the protocol reported by Saraiva et al. (2009). COCs were denuded with 0.2% (wt/vol) hyaluronidase for 5 min, and then incubated in IVM medium supplemented with 0.05 μg/mL demecolcine for 2 h. Oocytes presenting the first polar body (1PB) and protrusion in the membrane were incubated in SOF supplemented with HEPES (HSOF) with 10% FCS and 7.5 μg/mL cytochalasin B for removal of the 1PB and protrusion, with minimal cytoplast removal. The procedure was performed under an inverted optical microscope (Olympus IX-70), and the 1PB and the protrusion was removed with a 25-μm (external diameter) glass pipette. Traditional EN was performed without exposure of the oocytes to demecolcine. The same conditions were employed, except that the oocytes were incubated in SOF with 10 μg/mL Hoechst 33342, and UV light was used to visualize the nucleus and to confirm EN.
For groups reconstructed with embryonic or somatic donor cells, a single blastomere or an individual fibroblast, respectively, was transferred to the perivitelline space of the recipient oocyte, and electrofusion was done in mannitol solution (0.28 M mannitol, 0.05 mM CaCl2, 0.1 mM MgSO4, 0.05 mM HEPES acid, and 3 mg/mL BSA). One pulse of 1.5 kV/cm was used during 70 μsec (Bordignon et al., 1999) for embryonic cells, and two 20-μsec pulses of 2.0 KV/cm for somatic cells, produced by a BTX Electrocell Manipulator 2001 (BTX, San Diego, CA). Fusion rates were evaluated 30–60 min after electrofusion.
Chemical activation
Successfully reconstructed oocytes were artificially activated by incubation in TCM-199 with HEPES and 10% FCS supplemented with 5 μM ionomycin for 5 min, followed by incubation in SOF supplemented with 2 mM 6-dimethylaminopurine (6-DMAP) for 4 h at 38.5°C under a humidified atmosphere of 5% CO2 in air.
Culture and evaluation of embryo development
Embryo culture was performed in 100-μL microdrops of SOF supplemented with 2.5% FCS and 5 mg/mL BSA at 38.5°C under a humidified atmosphere of 5% CO2. Groups of 10–20 one-cell embryos were cultured for 7 days until the blastocyst stage. The medium was renewed on days 3 and 5 of embryo culture.
Blastocysts from all groups were stored individually in PBS supplemented with 0.1% polyvinylpyrrolidone and 1 U/μL RNase inhibitor (Invitrogen, Carlsbad, CA, USA). They were immediately immersed in liquid nitrogen and stored at −80°C for later analysis of gene expression.
RNA extraction, reverse transcription, and qPCR amplification
Day-7 blastocysts (4–10 from each group) were individually submitted to RNA extraction using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and linear acrylamide (Ambion, Foster City, CA, USA) following the manufacturer's instructions. cDNA was synthesized using the ImProm-II Reverse Transcription System (Promega, Madison, WI, USA) according to the manufacturer's instructions, and the cDNA from each embryo was preamplified using TaqMan PreAmp Master Mix (Applied Biosystems, Foster City, CA, USA) with 45 nM of the primers shown in Table 1. The program of preamplification consisted of a denaturing cycle at 95°C for 10 min and 14 cycles of PCR (95°C for 15 sec, 57°C for 45 sec, and 60–C for 4 min).
Due to the high variability among embryos, in this study we assessed individual gene expression instead of embryo pool analysis. The preamplification of cDNA targets was necessary due to the small amount of mRNA obtained from each embryo. Nonetheless, in a separate experiment we evaluated the preamplification efficiency in female in vitro–produced embryos, according to the system validated by Mengual et al. (2008), to compare the correlation of cDNA quantity (Nanodrop, Thermo Scientific) and gene expression data (ΔCT) between non-preamplified (NPA; n=5) and preamplified (PA; n=5) samples.
Amplifications were performed in a real-time PCR thermocycler (Applied Biosystems 7500 Real Time PCR System, Applied Biosystems, Foster City, CA, USA) using SYBR Green (Platinum® SYBR® Green qPCR SuperMix-UDG, Invitrogen, Carlsbad, CA, USA), 2-μL of PA or NPA (for preamplification efficiency evaluation) cDNA, 0.20 μM of each primer pair for GAPDH and G6PD, and 0.1 μM for XIST. The reactions were carried out in triplicate, in a final volume of 20-μL and were initiated with incubation at 50°C for 2 min, followed by denaturation at 95°C for 10 min, and 45 amplification cycles at 95°C for 15 sec and at 60°C for 1 min. The melting curves observed after PCR amplification confirmed the specificity of the amplified products. Real-time PCR data were analyzed using the 2−ΔΔCT method employing GAPDH as reference gene (Goossens et al., 2005).
Experimental design
Experiment I
Real-time PCR quantification of XIST and G6PD transcripts was performed on female day-7 blastocysts from NT groups reconstructed with embryonic donor nuclei cells [conventional EN, ECNT conv; chemically assisted EN, ECNT deme (demecolcine)], which were compared to IVD and in vitro–fertilized (IVF) embryos.
Experiment II
Real-time PCR quantification of XIST and G6PD transcripts was performed on female embryos of the same groups as experiment I, but the NT groups were reconstructed with somatic donor nuclei cells.
Statistical analysis
In relation to preamplification efficiency evaluation, linear regression analysis was performed to compare cDNA quantity and gene expression data (ΔCT) from NPA targets (ΔCTNPA=CTNPA target−CTNPA GAPDH) vs. PA targets (ΔCTPA=CTPA target−CTNPA GAPDH). The correlation data were processed using the GraphPad Prism 4.0 software (GraphPad Prism, San Diego, CA, USA).
In experiments I and II, PCR efficiency was estimated using linear regression of the log of fluorescence at each cycle, using LinReg software (Ramakers et al., 2003) with default parameters (number of points between 4 and 6 and best correlation coefficient). For each pair of primers, the fluorescence threshold line was fixed at the average of the lower and higher fluorescence values used by the software to estimate PCR efficiency. To measure the differences, we used the pairwise fixed reallocation randomization test in the Relative Expression Software Tool (REST; Pfaffl et al., 2002). The null hypothesis (H0) was no different between groups, and the differences were considered to be statistically significant when the probability of the alternative hypothesis [P(H1)] was less than 0.05.
Results
Preamplification efficiency of specific cDNA prior to gene expression analysis
The preamplification process allowed us to load a greater amount of cDNA from each sample (∼1.100 ng for NPA vs. ∼1.500 ng for PA samples). We found an overall mean CT decrement of ∼8.0 in PA samples. High correlation (r=0.9109) of cDNA quantity was observed when the samples were compared. Furthermore, linear regression analysis showed a high correlation (p<0.05) between gene expression measurements (XIST, r=0.9277; G6PD, r=0.9107) of NPA and PA samples for the validated genes. Thus, the overall relative gene expression levels in PA samples remained proportional to the original gene expression levels in NPA samples.
Expression of X-linked (XIST and G6PD) genes in embryonic NT-derived embryos
Regarding XIST expression, we did not detect in any embryo in the IVD group. However, in the IVF group, four female embryos (80%; n=5) displayed detectable XIST expression (Table 2).
In embryos produced by ECNT, XIST expression varied according to the EN technique employed. In the conventional EN (ECNT conv) group, only one embryo (25%; n=4) expressed XIST transcripts. However, in the chemically assisted EN (ECNT deme) group, most embryos showed XIST expression (75%; n=4) (Table 2).
The relative mRNA abundance (RA) of XIST in embryos that expressed the gene from all groups was also evaluated, but no significant differences were found (Fig. 1). The IVD group was not evaluated because no embryo expressed the gene. However, it is important to highlight that the RA of day-7 blastocysts had high individual variability (Fig. 2).
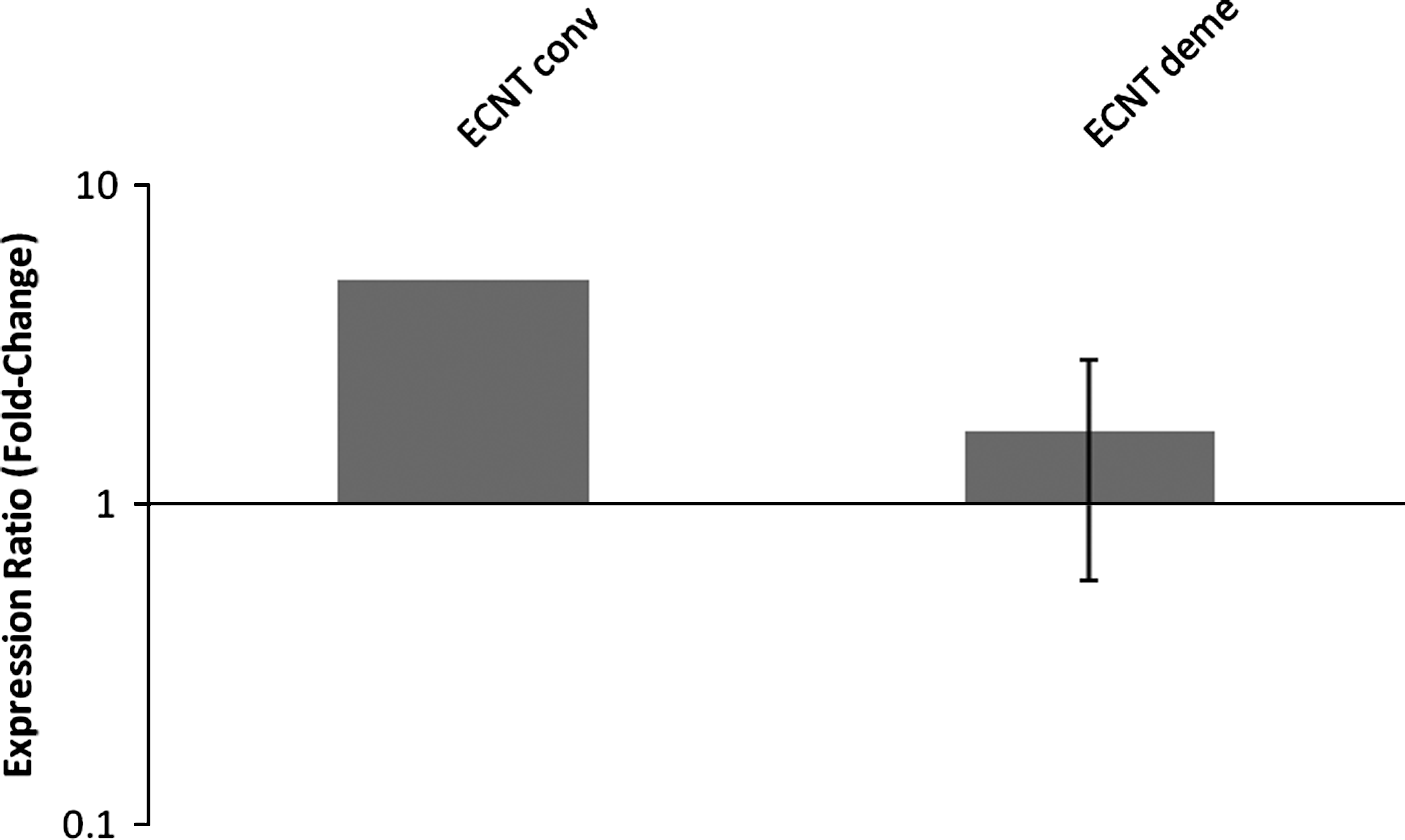
XIST expression in female bovine blastocysts produced by embryonic cell nuclear transfer after conventional (ECNT conv) or chemically assisted (ECNT deme) enucleation. Relative quantification (RQ) of XIST transcripts, normalized by GAPDH, was plotted as logarithmic expression and calibrated to the IVF group. Data are shown as mean±standard error. RQ in reference group (IVF) is considered 1.
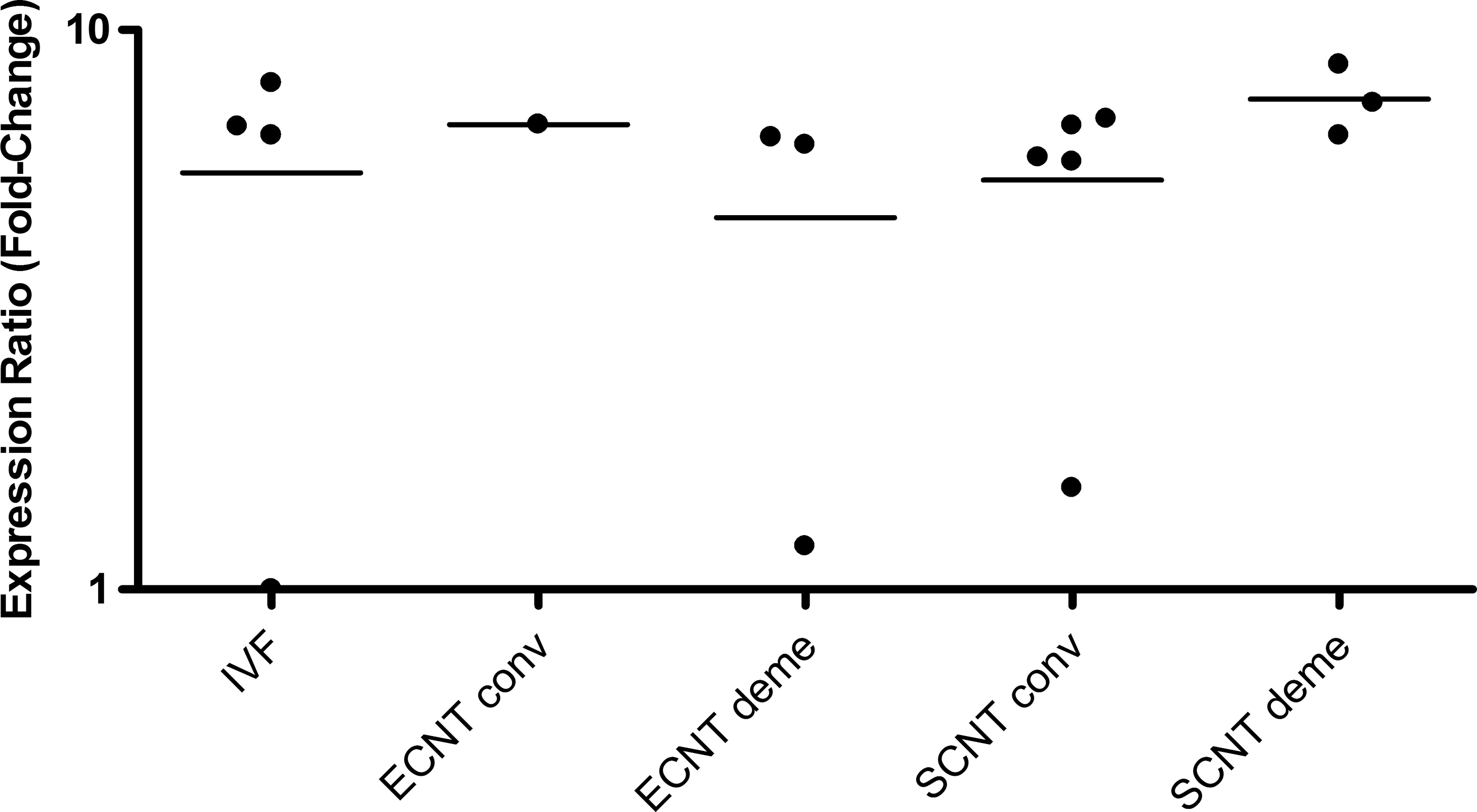
XIST relative expression level for each female bovine blastocyst produced in vitro (IVF), by embryonic cell nuclear transfer (ECNT), or by somatic cell nuclear transfer (SCNT) after conventional (conv) or chemically assisted (deme) enucleation. Relative quantification (RQ) of XIST transcripts, normalized by GAPDH, was plotted as logarithmic expression and calibrated to one blastocyst from IVF group with the lower RQ. Relative quantification in reference group (IVF) is considered 1.
G6PD transcripts were detected in all female IVD embryos (n=4), but just in 60% of IVF embryos (n=5; Table 2), probably indicating deviations from the normal expression pattern. In NT-derived embryos, G6PD expression was observed in 100% and 75% of the embryos in ECNT conv (n=4) and ECNT deme (n=4) groups, respectively (Table 2).
No significant differences in RA of G6PD were observed (Fig. 3) in embryos produced by ECNT. Nonetheless, a trend toward higher G6PD expression was observed in IVD compared to IVF female embryos (Fig. 3). Once again, the large variability between embryos may have contributed to the lack of statistical differences among groups (Fig. 4).
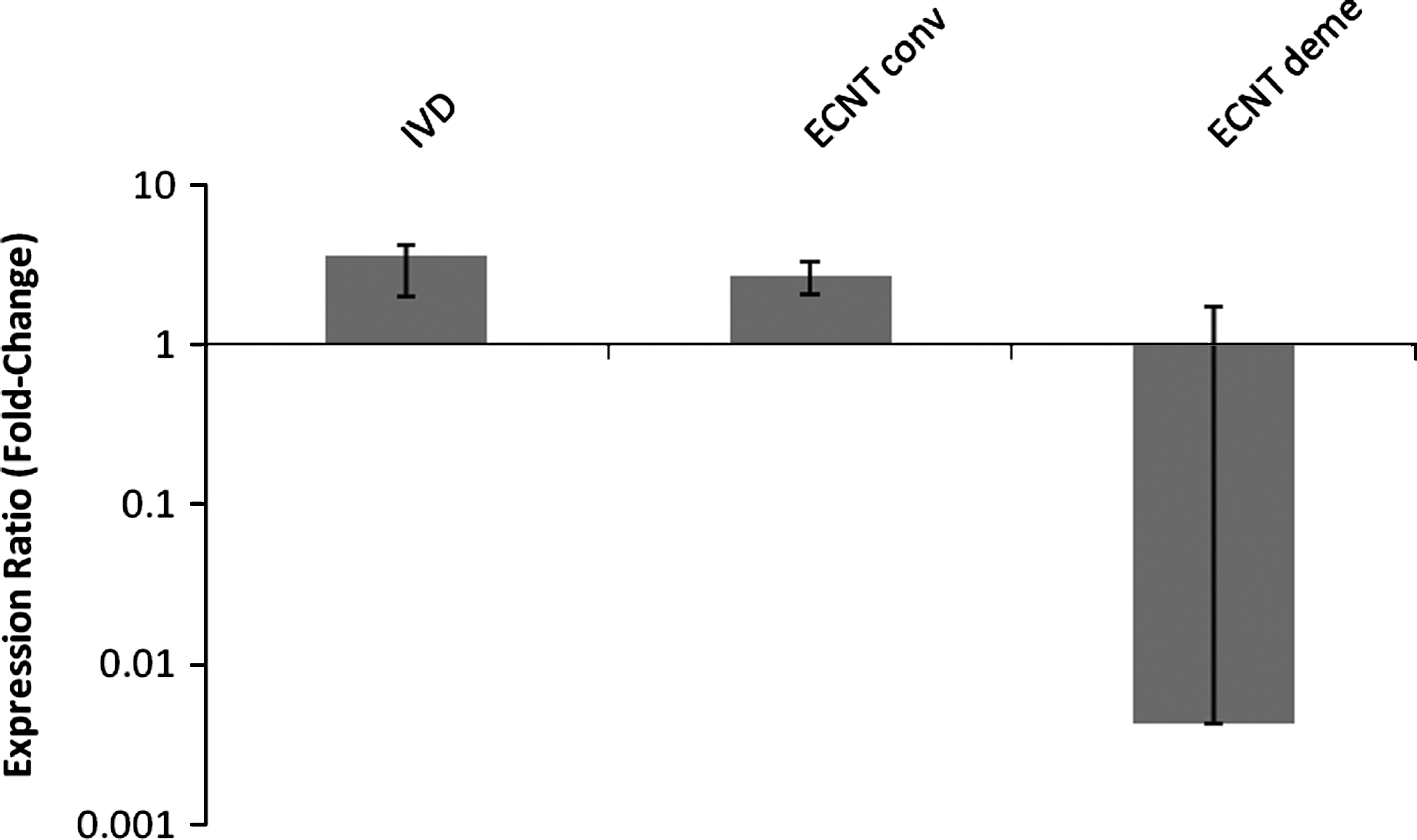
G6PD expression in female bovine blastocysts produced in vivo (IVD) or by embryonic cell nuclear transfer after conventional (ECNT conv) or chemically assisted (ECNT deme) enucleation. Relative quantification (RQ) of G6PD transcripts, normalized by GAPDH, was plotted as logarithmic expression and calibrated to the in vitro-fertilized (IVF) group. Data are shown as mean±SE. RQ in reference group (IVF) is considered 1.
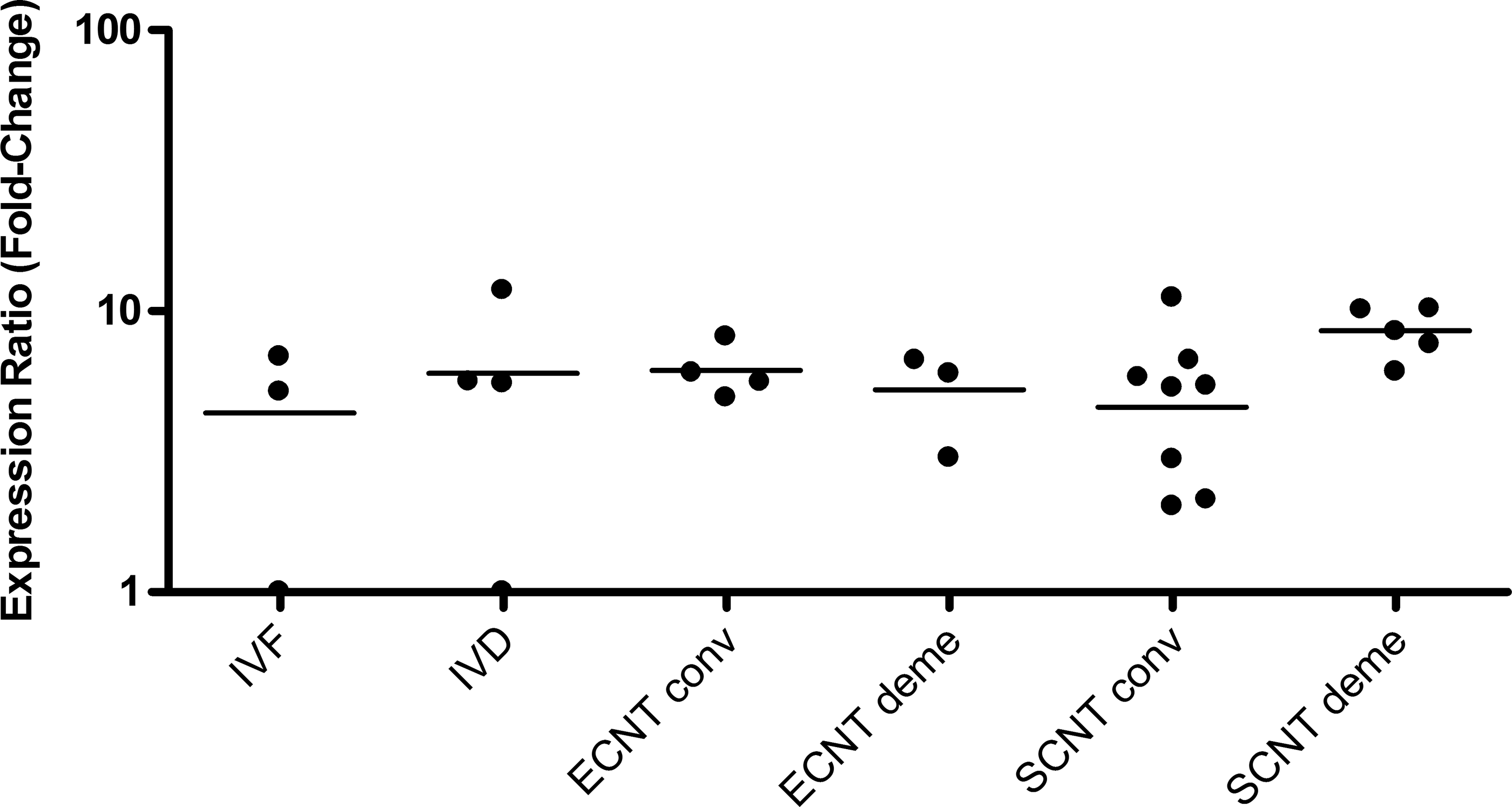
G6PD relative expression level for each female bovine blastocyst produced in vitro (IVF), by embryonic cell nuclear transfer (ECNT), or by somatic cell nuclear transfer (SCNT) after conventional (conv) or chemically assisted (deme) enucleation. Relative quantification (RQ) of G6PD transcripts, normalized by GAPDH, was plotted as logarithmic expression and calibrated to one blastocyst from IVF group with the lower RQ. Relative quantification in reference group (IVF) is considered 1.
Expression of X-linked (XIST and G6PD) genes in somatic cell NT-derived embryos
Both NT groups showed high percentages of embryos expressing the XIST gene (55% in SCNT conv, n=9; 50% in SCNT deme, n=6; Table 2). No significant differences in RA of XIST were observed among groups (Fig. 5).
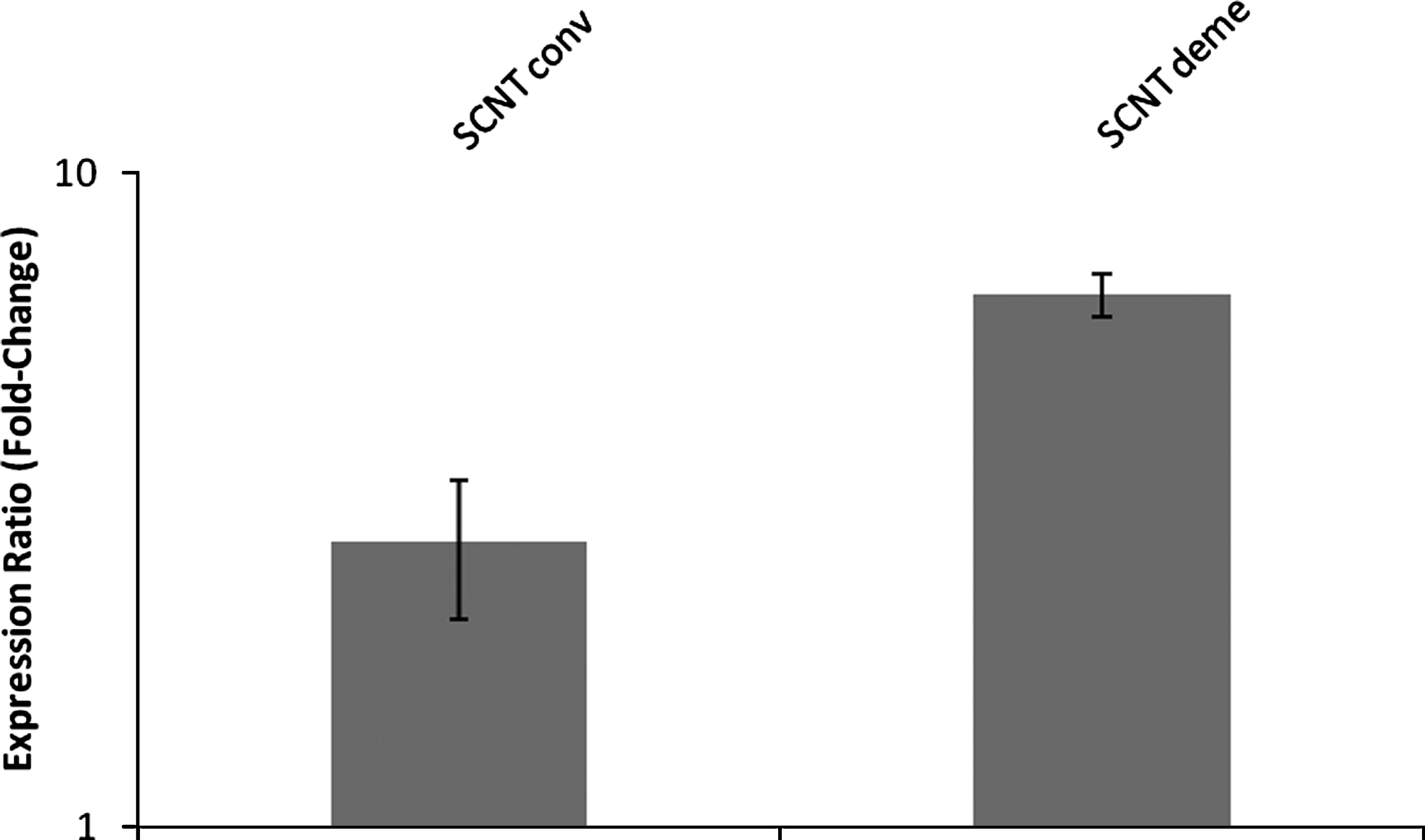
XIST expression in female bovine blastocysts produced by somatic cell nuclear transfer after conventional (SCNT conv) or chemically assisted (SCNT deme) enucleation. Relative quantification (RQ) of XIST transcripts, normalized by GAPDH, was plotted as logarithmic expression and calibrated to the in vitro-fertilized (IVF) group. Data are shown as mean±SE. RQ in reference group (IVF) is considered 1.
G6PD was expressed in almost all NT embryos (100% and 83.3% for SCNT conv and deme, respectively; Table 2). Relatively higher (p<0.05) levels of G6PD transcripts were observed in the SCNT deme and IVD groups in comparison to the SCNT conv group (Fig. 6). Nonetheless, no significant differences were found between the IVD and IVF groups.
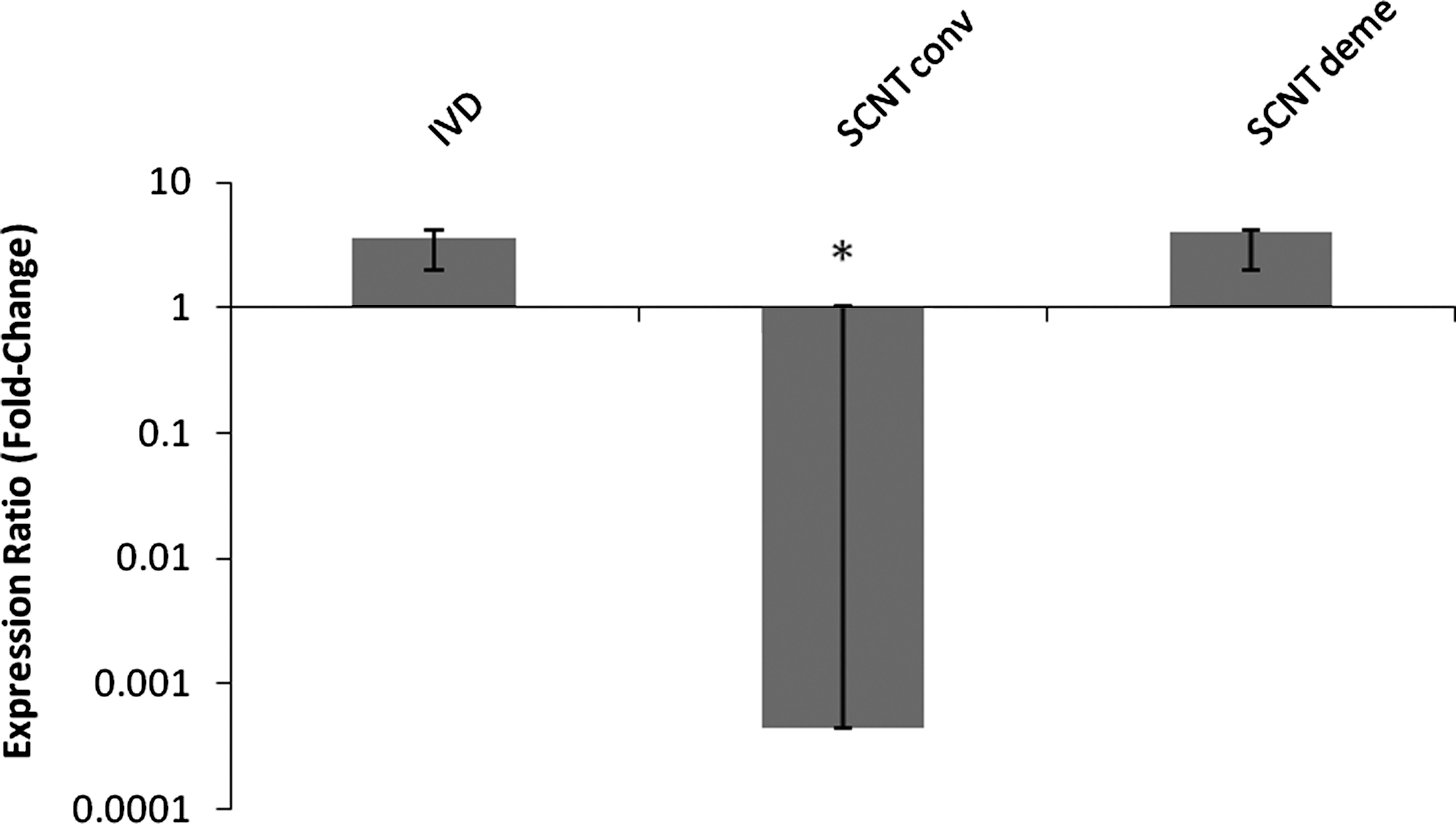
G6PD expression in female bovine blastocysts produced in vivo (IVD) or by somatic cell nuclear transfer after conventional (SCNT conv) or chemically assisted (SCNT deme) enucleation. Relative quantification (RQ) of G6PD transcripts, normalized by GAPDH, was plotted as logarithmic expression and calibrated to the in vitro-fertilized (IVF) group. Data are shown as mean±SE. RQ in reference group (IVF) is considered 1.
Discussion
We assessed XIST and G6PD gene expression in embryos IVD, IVF, and reconstructed with embryonic blastomeres or with somatic cell nuclei, comparing two EN techniques (conventional and chemically assisted). The main findings of the present study are that: (1) XIST mRNA was not detected in day-7 IVD blastocysts, in contrast to IVF embryos; (2) a different XIST and G6PD expression pattern was observed in NT-derived embryos, according to the source of donor nucleus cells (embryonic versus somatic cells) and to the EN technique (conventional versus chemically assisted); and (3) higher RA of G6PD was verified in SCNT embryos produced with chemically assisted enucleated cytoplasts.
Because accurate gene expression quantification can be limited by the low RNA quantity obtained from some clinical samples (Mengual et al., 2008), such as mammalian embryos, in this study it was necessary to preamplify the cDNA of the samples. A novel cDNA preamplification system enables multiplex preamplification of cDNA targets and, therefore, could provide a sufficient amount of specific amplicons for their further analysis. Recently, this method was validated by Mengual et al. (2008) by performing a multiplex preamplification of 47 genes in 22 samples, comparing the relative gene expression levels between NPA and PA samples. They found a high correlation (r) between the gene expression measurements of NPA and PA samples (r=0.970). Nevertheless, they reported that checking the preamplification uniformity in each target gene against control material before its evaluation in testing samples is mandatory. Our results regarding preamplification were similar to those previously reported, with high correlations of cDNA quantity and relative expression between NPA and PA samples, proving that the levels in PA samples remained proportional to the original levels, thus allowing appropriate interpretations of the results obtained.
Besides that, like most studies, our work used IVD embryos as a control group, obtained after superovulation of female donors, to obtain a sufficient number of structures for assessment. However, there are reports in humans and mice that show interference of superovulation on the methylation patterns of certain genes. Market-Velker et al. (2010) recently observed in human blastocysts that this procedure perturbed genomic imprinting of maternally and paternally expressed genes in a dose-dependent manner. Because of these reports in humans and mice, one must be careful in extrapolating these data to bovine species, as preimplantation embryos of different mammalian species differ in many aspects at the cellular level and regarding genetic control (Gardner and Lane, 2005), and also some genes imprinted in mice and humans are not necessarily imprinted in the domestic animals.
It is known that one possible reason for the limited success of bovine cloning is the incorrect reprogramming of the transferred nucleus in reconstructed embryos. It has been speculated that ooplasmic proteins imported into the nucleus mediate structural rearrangement of the chromatin (Bordignon et al., 2001), showing the importance of cytoplasts in the nuclear reprogramming process. It is believed that one beneficial consequence of chemically assisted EN is the presence of larger cytoplasmic volume, and hence more reprogramming factors.
Regarding X-linked gene expression, we did not find XIST expression in IVD embryos, indicating a possible physiological condition of day-7 blastocysts. Thus, a disruption of normal XIST expression likely occurs in IVF embryos, evidenced by the high percentage of embryos expressing the XIST gene, as shown in previous studies (Merighe et al., 2009; Wrenzycki et al., 2002).
In the experiment with ECNT, we observed a similar XIST expression pattern between NT-derived embryos from the conventional EN technique and IVD group, with only a few embryos presenting expression of this gene. Therefore, when we used embryonic blastomeres in the NT, the conventional technique was able to maintain the XIST gene expression patterns observed in the in vivo group. This was probably due to the source of the donor cell nuclei.
Successful nuclear reprogramming requires erasure of the donor nuclei's epigenetic pattern and the re-establishment of embryonic epigenetic characteristics and gene expression in the cloned embryo (Tonge and Nagy, 2010). NT-derived embryos typically show abnormal patterns of DNA methylation and histone modifications (Blelloch et al., 2006), and the development of these embryos appears to be influenced by the differentiation state of the donor genome (Hochedlinger and Jaenisch 2002; Rideout et al., 2001). Differentiation imposes structural constraints on the donor chromatin that limit access of the transcriptional machinery to regulatory sequences in the genome (Oback and Wells, 2002). Blastomeres show little or no differentiation and may retain factors that are necessary for early embryonic development and do not have to be reprogrammed. Furthermore, the DNA methylation status of blastomeres is more compatible with early development. Nonetheless, the ECNT deme group displayed a high percentage of embryos expressing the XIST gene, even with the use of embryonic cells in the reconstitution, signaling for a possible abnormal X-linked gene expression pattern caused by the chemically assisted technique.
When somatic cells were used in the NT procedure, both techniques resulted in a higher percentage of embryos expressing XIST transcript compared to the in vivo group, i.e., micromanipulation techniques probably did not permit the correct nuclear reprogramming of somatic cells and caused such changes in embryo expression patterns. Recently, Inoue et al. (2010) reported that SCNT fails to regulate XIST expression from the X chromosome. The authors observed that SCNT embryos are not able to suitably repress XIST activity, and ectopic XIST expression was present in both male and female preimplantation embryos. By using XIST-deficient donor nuclei, the authors showed that this ectopic expression in cloned blastocysts contributed to the downregulation of half of the affected X-linked genes, and the modification of gene expression levels had a dramatic effect on the efficiency of generating viable cloned animals, which reached more than 10%, corresponding to eight- to nine-fold higher success rate when compared to wild-type controls.
Regarding the G6PD gene, we found that 100% of the IVD, ECNT conv, and SCNT conv groups' embryos showed transcripts on day 7. In the IVF, ECNT deme, and SCNT deme groups, the proportions were 60%, 75%, and 83.3%, respectively. Apparently, embryos produced by the conventional technique follow patterns more similar to those displayed by in vivo–produced embryos in comparison with embryos obtained by the chemically assisted technique. Nonetheless, in the RA analyses, higher levels of G6PD transcript were observed in the SCNT deme group in comparison to embryos produced by the conventional technique. These levels were similar to those presented by the IVD group, signaling the normality of the results found in the chemically assisted group.
Controversial reports have been published on the dosage compensation of the G6PD gene. Previous reports have shown higher levels of G6PD expression in female IVF blastocysts than in their male counterparts, whereas the levels found for both sexes in IVD embryos were similar (Gutierrez-Adan et al., 2000; Lucas-Hahn et al., 2001; Peippo et al., 2002; Wrenzycki et al., 2002). This phenomenon has been associated with alterations in dosage compensation under in vitro conditions caused by problems or delay in XCI. However, there are reports that show lower G6PD transcript levels in female than in male IVF embryos, suggesting double silencing or other mechanisms of control for this gene. Although we did not find significant differences in G6PD expression between IVD and IVF embryos, probably due to the large variability observed for individual embryos, we observed a trend to higher G6PD expression in female IVD in comparison to IVF embryos (almost two-fold). This finding can be explained by a recent report (Bermejo-Alvarez et al., 2011) showing that in bovine species substantial XCI probably occurs between blastocyst hatching and initiation of elongation. Therefore, higher G6PD levels indicate that XCI has not yet occurred in female IVD blastocysts, unlike in female IVF embryos.
Many of the previous studies measured the expression of genes by a semiquantitative real-time PCR technique (Daniels et al., 2000; Wrenzycki et al., 2002). Detailed quantitative analysis of individual blastocysts has revealed a large degree of embryo variation and subtle changes in expression levels. This may have contributed to the different results observed in our study.
The G6PD gene encodes the rate-limiting enzyme of the pentose-phosphate pathway (PPP) (Rieger, 1992). It was also found that GPD6 acts as a sentinel responsible for maintaining the cellular redox state due to oxidative stress (Gutierrez-Adan et al., 2004). Through the nicotinamide adenine dinucleotide phosphate (NADPH) generation in the PPP, G6PD also participates in the detoxification of oxygen radicals (Nicol et al., 2000; Peippo and Bredbacka, 1995) and therefore has been considered a cytoprotective enzyme for oxidative stress and DNA damage.
Lopes et al. (2007) suggested the possibility that the increased G6PD expression is indicative of embryo quality. Oxygen consumption of individual embryos increases significantly around compactation and blastulation, and it is higher among embryos of superior quality (Agung et al., 2005; Lopes et al., 2006). Embryos from more advanced stages (Donnay and Leese, 1999; Sturmey and Leese, 2003; Thompson et al., 1995) and good-quality female bovine embryos (Agung et al., 2005) have also been related to higher oxygen consumption. In addition, Lopes et al. (2007) demonstrated that the respiration rates of IVP embryos correlate with the mRNA levels of the G6PD gene.
Alternatively, the increased G6PD expression associated with higher oxygen uptake by the embryos could have been caused by the presence of ROS. Higher oxygen consumption causes enhanced adenosine triphosphate (ATP) production by mitochondrial oxidative phosphorylation, but also produces ROS (Harvey et al., 2002), and thus high respiration rates could be a manifestation of metabolic stress instead of superior quality (Lopes et al., 2007).
Recent data regarding embryonic and fetal development after the use of the chemically assisted technique has been encouraging, with rates similar (Li et al., 2006; Tani et al., 2006) or superior (Lan et al., 2008; Li et al., 2009; Meng et al., 2011) to those obtained with the traditional technique. In bovine species, higher blastocyst rates and average cell numbers have been reported [40% and 103, respectively, by Li et al. (2009), and 62.2% and 130 cells by Meng et al. (2011)] for the chemically assisted technique than for the controls. Also, Li et al. (2009) had a higher pregnancy rate (12.5% vs. 3.3%) when demecolcine was used in the process. Therefore, despite the apparent disruption in the expression of X-linked genes in embryos produced by the chemically assisted technique, these structures can develop normally, with similar or superior rates than those obtained by the conventional technique. It is possible the higher RA levels in those embryos that are able to properly express the G6PD gene are related to this fact.
In conclusion, the results obtained with the use of the chemically assisted technique show three possibilities: (1) alteration of dosage compensation due to disturbance of XCI; (2) indication of higher metabolism in superior-quality embryos; and/or (3) G6PD activation resulting from the presence of higher ROS levels generated by increased oxygen consumption. The expression of stress response–related genes should be investigated in embryos produced by chemically assisted EN to reveal whether the differences observed between the techniques are indicative of embryo quality.
Footnotes
Acknowledgments
This research was supported by the Foundation for the Support of Research of the State of São Paulo (FAPESP), Brazil. The authors thank Roberta Vantini for laboratory assistance, Luciana Correia de Almeida Regitano (Embrapa Southeast Livestock) for allowing the use of qPCR apparatus, and Adriana Mercia Guaratini Ibelli for contributions during the real-time PCR analysis.
Author Disclosure Statement
No competing financial interests exist.