
Abstract
Abstract
Distinct signaling pathways are reported to maintain pluripotency in embryo-derived stem cells. Mouse embryonic stem cells (ESCs) respond to leukemia inhibitory factor (LIF) and bone morphogenetic protein (BMP)-mediated activity, whereas human ESCs depend upon Fibroblast growth factor (FGF) and activin signaling. In the majority of mammals investigated, however, the signals that support stem cell pluripotency are not well defined, as is evident by the persistent difficulties in maintaining authentic stable ESC lines. Induction of pluripotency by transcription factor-mediated reprogramming could provide an alternative way to produce ESC-like cells from nonpermissive species, and facilitate identification of core ESC signaling requirements. To evaluate the effectiveness of this approach in pigs, we transduced porcine foetal fibroblasts with retroviruses expressing Oct4, Sox2, Klf4, and c-Myc, and maintained the resulting cultures in medium containing either LIF or FGF2. Alkaline phosphatase positive colonies with compact, mouse ESC-like morphology were preferentially recovered using serum-free medium supplemented with LIF. These cell lines expressed the endogenous stem cell transcription factors, OCT4, NANOG, and SOX2, and the cell surface marker SSEA-4, consistent with acquisition of an undifferentiated state. However, restricted differentiation potential, and persistent expression of retroviral transgenes indicated that reprogramming was incomplete. Interestingly, LIF activated both the transcription factor STAT3 and its target gene SOCS3, and stimulated cell growth, indicating functional coupling of the signaling pathway in these cells. This demonstration of LIF-dependence in reprogrammed pig cells supports the notion that the connection between LIF/STAT3 signaling and the core regulatory network of pluripotent stem cells is a conserved pathway in mammals.
Introduction
The distinct growth factor requirements of mouse and human ESCs led to the proposal that these cells represent different stem cell states (Nichols and Smith, 2009). Pivotal in this conclusion was the discovery that mouse stem cell lines derived from postimplantation embryos also exhibited FGF/activin-dependent growth and possessed a gene expression profile more closely aligned with human ESCs than with mouse ESCs (Brons et al., 2007; Tesar et al., 2007). These mouse epiblast stem cell (mEpiSC) lines represent an advanced, “primed” state, distinct from the more primitive “naive” mESC derived from the preimplantation epiblast. The very limited capacity of mEpiSCs to recolonize the epiblast of host blastocysts observed in initial studies, highlighted functional differences between the two cell types (Brons et al., 2007; Tesar et al., 2007). However, more recent studies have described functional heterogeneity within each cell type, stable capture of intermediate states, and even reversion of epiblast or EpiSCs to an ESC-like cell (Greber et al., 2010; Han et al., 2010; Silva et al., 2008). This apparent destabilization of the boundaries between ESC and EpiSCs has in part been revealed through the use of selective culture conditions. For instance, the chemical manipulation of ESC signaling pathways and stimulation with LIF promoted mEpiSC reprogramming to an ESC state (Silva et al., 2008; Yang et al., 2010). In addition, forced transgenic expression of key stem cell regulators can induce human ESC to adopt an ES cell-like state akin to mESC (Buecker et al., 2010; Hanna et al., 2010).
The recent discovery that expression of four transcription factors Oct4, Sox2, KLF4, and c-Myc can induce the dedifferentiation of somatic cells into a pluripotent (iPSC) ESC-like stem cells (Takahashi and Yamanaka, 2006; Takahashi et al., 2007), not only has important implications for applying stem cells in the study of human disease, but also attracted the attention of researchers trying to generate pluripotent stem cell lines from nonpermissive species (Telugu et al., 2010b). Crucially, for mouse and human cells, the iPSC procedure can reestablish pluripotency by reactivating the endogenous ES cell regulatory circuits, rendering iPSCs independent of the initiating reprogramming factors (Kaji et al., 2009; Soldner et al., 2009; Sommer et al., 2010; Woltjen et al., 2009). Remarkably, mouse iPS clones can pass the most stringent test for ESC developmental potency: they can generate viable pups through tetraploid aggregation that are wholly derived from iPSCs (Boland et al., 2009; Kang et al., 2011; Zhao et al., 2009). Reprogramming of human somatic cells also produces cell lines that exhibit most of the key characteristics of human ESCs (Soldner et al., 2009). In light of these achievements, a number of laboratories have attempted to generate equivalent reprogrammed stem cells from livestock species including the pig (Bao et al., 2011; Esteban et al., 2009; Ezashi et al., 2009; Li et al., 2011; Telugu et al., 2010a; Wu et al., 2009).
The reprogrammed pig cell lines described in the three initial reports, express endogenous stem cell markers, and display multilineage differentiation in teratomas (Esteban et al., 2009; Ezashi et al., 2009; Wu et al., 2009). However, unlike mouse and human iPSCs, the stem cell phenotype of these pig cells is reliant on continuous expression of the reprogramming transgenes, indicating that the endogenous porcine stem cell circuitry has not been appropriately reinstalled or that current culture conditions do not support pig iPSC pluripotency and self-renewal. Indeed, whereas mouse and human iPSCs exhibit the growth factor requirements of their respective ESC counterparts, the culture conditions that support long-term propagation of pig ESCs or iPSCs have not yet been defined. The use of hESC growth conditions to isolate pig iPSCs and their hESC-like colony morphology, however, suggests that the pig iPSCs reported recently are most closely related to EpiSC-like cells (Esteban et al., 2009; Ezashi et al., 2009; Wu et al., 2009). A potential limitation of EpiSC cells is that they tolerate cell dissociation poorly, can display intrinsic biases in differentiation in vitro and exhibit limited developmental capacity in vivo (Brons et al., 2007; Nichols and Smith, 2009; Tesar et al., 2007). This compromises handling and manipulation of these cells in vitro, and limits their utility in vivo, for example, in germline genetic modification. Recognition of the generally more robust and flexible character of naive type ESCs has fuelled efforts to derive human cell lines equivalent to mouse ESCs. Indeed, human cells with these characteristics have been generated recently by artificially maintaining expression of reprogramming genes and applying the cytokine LIF in conjunction with chemical inhibitors of two signaling pathways (2i) that establishes mESC-like “ground-state” growth conditions (Buecker et al., 2010; Hanna et al., 2010). It has also been recently reported that pig IPSC lines phenotypically resembling mESCs can be generated with a further modification of this approach using the chromatin modifying agents VPA and 5-azacytidine (Telugu et al., 2010a). Thus, enforced genetic programming combined with chemical manipulation of extrinsic signaling may support stable propagation of alternative cryptic stem cell states, and may enable derivation of pluripotent stem cell lines from nonpermissive species.
Here we describe reprogrammed cell lines derived from porcine fetal fibroblasts by retroviral transduction of four mouse stem cell factors (Oct4, Sox2, Klf4, and c-Myc) following culture in mESC growth conditions. These novel pig cell lines expressed endogenous markers of pluripotency but had restricted differentiation potential, a possible consequence of incomplete silencing of the retroviral transgenes. Interestingly, although this might suggest that these cells were not fully reprogrammed, their growth was strongly stimulated by LIF, pointing to activation of latent ESC regulatory circuitry within these pig cell lines.
Materials and Methods
Cell culture and retroviral transduction
Porcine embryonic fibroblasts (PEFs) were isolated from fetuses (approximately embryonic day 40) arising from a mating between a Large White boar carrying a CMV-GFP transgene and a wild-type gilt. Pig PEFs were cultured in DMEM culture medium (Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum (FBS), 2 mM L-Glutamine, 0.1 mM nonessential amino acids, and 1 mM sodium pyruvate. PEFs were plated at a density of 1.3×105 cells per well (9.6 cm2) in six-well dishes. Cells were transduced 16 h after plating. Virus was produced by transfecting HEK293T with pMXs-based retroviral vectors containing the mouse cDNAs for c-Myc, Klf4, Sox2, and Oct4 (Takahashi and Yamanaka, 2006) and pCL-10A1 (Imgenex Cat# 10047P). For transfection of each pMXs retrovirus vector, 8 μg or pCL-10A1 and 8 μg of one of the pMXs plasmid were mixed with 68 μL Fugene HD in Optimem and used to transfect 3.6×106 HEK293T/17 cells. Supernatants were removed 24 h after transfection, and replaced with fresh HEK293T/17 culture medium. The collected supernatants were filtered through a 0.45 μm filter and mixed. Polybrene was added at 8 μg/mL and 1 mL of supernatant applied to each well of PEFs. This transduction procedure was repeated 24 h later so each well had two applications of retrovirus. The next day the transduced PEF cells were passaged using accutase and plated onto SNL feeder cells (SNL76/7, ECACC) (Takahashi et al., 2007). SNL cells are STO fibroblasts stably transfected with a G418 resistance cassette and LIF expression vector. Each well was passaged into four wells, three wells for continued culture and the remainder stored as a frozen stock. Three culture media were tested; FL composed of GMEM with 10% FBS, 1 mM glutamine, 0.1 mM nonessential amino acids, 1 mM sodium pyruvate, 0.1 mM β-meracaptoethanol and 103 U/mL hLIF (Millipore, Bedford, MA). SRL was composed of KO-DMEM with 20% serum replacement containing 2 mM L-Glutamine, 0.1 mM nonessential amino acids, 0.1 mM β-mercaptoethanol (SR) supplemented with 103 U/mL hLIF. SF contained SR medium supplemented with 4 ng/mL hFGF2 (Peprotech, Rocky Hill, NJ). Reprogrammed pig lines were maintained on feeders and passaged using accutase.
Alkaline phosphatase (AP) staining and immunocytochemistry
AP staining was performed using the Leukocyte Alkaline Phosphatase Kit (Sigma, St. Louis, MO). For immunocytochemistry, cells were fixed in 4% paraformaldehyde in PBS or 10 min at room temperature washed three times with phosphate-buffered saline (PBS) and then processed as described previously (Meek et al., 2010). Antibodies used were raised against Oct4 (C-10, Santa Cruz, Santa Cruz, CA, 1:200), Nanog (Ab21603, Abcam, Cambridge, MA. 1:200), SSEA-1 and −4 (MC-480, MC-813-70, DSHB, 1:200), Class III β-tubulin (Tuj1, Covance, Richmond, CA, 1:200). Primary antibodies were detected using Alexa Fluor 488 or 595 conjugated secondary antibodies (Invitrogen). Nuclei were stained with DAPI (10 μg/mL).
Microarray profiling
RNA was prepared from two reprogrammed pig lines (A1 and D3), the PEF culture used for the reprogramming, and a differentiated D3 culture. To minimize the contribution of SNL feeders in the A1 and D3 samples, the iPS cell suspensions were preplated on uncoated tissue culture plastic for 15 min, to allow attachment of feeders and facilitate their selective removal. RNA was extracted from the cell pellets using the Qiashredder and RNeasy kit (Qiagen, Chatsworth, CA). Gene expression in the pig cells was examined using a modified Agilent Porcine 44 K array. The standard Porcine Agilent microarray design 020109 was augmented by the addition of oligonucleotides for 48 genes implicated as ES cell differentiation markers (Supplementary Table 1; Supplementary Data are available online at http://www.liebertonline.com/cell). The oligonucleotides synthesized on the microarrays were designed using Agilent eArray software (https://earray.chem.agilent.com/earray/; AMADID design number 023540). Duplicate aliquots of each sample RNA were labeled using Ambion's MessageAmp kit, which included the addition of an aminoallyl modified nucleotide during the in vitro transcription. The resulting amplified RNA was coupled to Cy3 dye and then hybridized with the arrays using the Agilent Gene Expression hybridization kit according to the manufacturer's protocols. The slides were scanned using an Axon 4200 AL scanner set at 5 micron scanning. Image files were analyzed and normalized using the Agilent Feature extraction software version 9. Initial data analysis was carried out using Partek Genomics Suite (Partek Inc., St. Louis, MO) using the standard Gene Expression Workflow. Genes that were differentially expressed between the four cell populations were also identified using the Bioconductor packages limma, affyPLM, genefilter and statmod. Preprocessing of the data was carried out using the genefilter package to remove control probes and features with a mean expression intensity <40 across all arrays. The data for the remaining 31,861 was log2 transformed and quantile normalized using the Bioconductor package affyPLM. In order to identify transcripts differentially expressed between the different populations the Empirical Bayes function was used within the limma package followed by correction for multiple testing using the Hochberg & Benjamini procedure (Hochberg and Benjamini, 1990). Transcripts exhibiting a more than twofold change in expression with an adjusted p-value <0.05, were considered to be significantly differentially expressed. The data from all 4223 transcripts matching these criteria were then saved as an “.expression” file. Following import into BioLayout Express3D (Freeman et al., 2007) (www.biolayout.org) a pairwise Pearson correlation matrix was calculated thereby performing an all versus all comparison of the expression profile of each probe on the array. All Pearson correlations where r≥0.9, were saved to a “.pearson” file. Based on a user defined threshold of r=0.99, an undirected network graph of the data was generated. In this context nodes represent individual probes (genes/transcripts) and the edges between them are Pearson correlation coefficients above the selected threshold. The resulting network graph was then clustered into groups of genes sharing similar profiles using the MCL algorithm with an MCL inflation value, to control granularity of clustering, set at 1.5.
Reverse transcription-polymerase chain reaction (RT-PCR)
Total RNA was prepared using Qiashredder and RNeasy kit (Qiagen) following the manufacturer's protocol. Total RNA was reverse-transcribed using Superscript First-Strand Synthesis kit (Invitrogen). PCR amplification was performed using Taq polymerase (Invitrogen). Primers used are listed in Supplementary Table S2. PCR was performed by denaturing the cDNA at 95°C for 2 min followed by 30 cycles of denaturing at 94°C for 60 sec, annealing at temperatures listed in Supplementary Table S1 for 30 sec and extension at 72°C for 30 sec with a final 10 min extension at 72°C. PCR products were analyzed on 2% agarose gels. For analysis of SOCS3 induction by LIF, cells were plated on PBS rinsed, laminin-coated wells (10 μg/mL laminin in PBS for 3–24 h at 37°C). To facilitate selective removal of fibroblast feeders, cultures were preplated on uncoated tissue culture plastic for 15 min. Recovered cells were seeded on the laminin coated wells in SRL medium. The next day SRL medium was replaced with either SRL or LIF free medium for 24 h. Cells were then treated with LIF for 1 h and the cells were harvested for RNA.
Real-time qPCR
Real-time PCR was performed using Platinum SYBR Green (Invitrogen, 11733). Reactions were amplified on a Stratagene Mx3000P qPCR thermocycler under the following conditions; one cycle of 50°C for 2 min, 95°C for 2 min, followed by 40 cycles of 95°C for 15 sec, 60°C for 30 sec, and a final dissociation step of 95°C for 1 min, 60°C for 30 sec, and 95°C 15 sec. Primer sequences are detailed in Supplementary Table S2. Using the ΔCt method, mRNA expression was normalized relative to β-actin.
In vitro differentiation
Cells were detached using accutase and transferred to low attachment six-well plates in differentiation medium (DMEM, 10% FBS, 1 mM glutamine, 0.1 mM nonesential amino acids, 0.1 mM sodium pyruvate, 0.1 mM β-mercaptoethanol). Aggregated cells were maintained in suspension for 5 days. Embryoid bodies (EBs) were then plated onto gelatin-coated tissue culture dishes and incubated for another 12 days.
LIF Induction and Western blotting
Preplated pig lines were seeded on laminin-coated plates (see above) 6×105 cells/six-well cluster in SRL medium. The next day SRL medium was replaced with LIF free medium for 3 h. The cultures were then treated with mouse LIF at 1×103 Units/mL or human LIF at 1×103 Units/mL for 10 min. Cells were then lysed and analyzed by Western blotting for levels of phosphorylated and total STAT3 as described previously (Burdon et al., 1999).
Cell proliferation assay
Reprogrammed pig lines were plated on laminin-coated plates (see above). Cells were plated in 96-well plates at a density of 1×104 cells/well in 100 μL/well with six replicates per time point. Cell proliferation was assessed using the Cyquant direct cell proliferation assay kit (Invitrogen). At each time point wells were incubated with an equal volume of detection reagent at 37°C for 1 h, after which fluorescence was measured using a 488/520-nm filter in a multilabel plate reader (Wallac, Waltham, MA).
Karyotyping
Cells were treated with 0.1 μg/mL colcemid (Invitrogen) for 2 h at 37°C before being harvested for metaphase preparation. Cells were then trypsinized, pelleted by centrifugation at 250×g for 8 min, agitated on a vortex mixer, and resuspended in 10 mL of 75 mM KCl, and incubated for 8 min at room temperature (RT). Cells were fixed by adding 1 mL of ice-cold fixative solution composed of one part of acetic acid and three parts of methanol, and mixed gently by inversion. Cells were pelleted at 400×g for 5 min and resuspended in 3 mL fixative solution by gentle inversion. After repeating this washing step two more times, the recovered cell pellet was finally resuspended in 100–200 μL of fixative solution and 10–20 μL dropped onto a glass slide. Slides were incubated at 37°C overnight and then stained with Giemsa for 15 min, rinsed three times with water, and mounted with coverslips.
Teratoma formation and analysis
Approximately 1–5×106 cells (per injection) from three reprogrammed pig lines were injected under the kidney capsule or intramuscularly into the hind leg of a nonobsese diabetic/severe combined immune deficient mice (NOD/SCID; Harlan UK Ltd.). All mice injected with lines isolated and grown in SRL gave rise to tumors (Supplementary Table S3). Three of five mice injected intramuscularly with the SRF line A2 also developed tumors. Tumors were harvested ≥3 months after injection, dissected, and fixed in formalin. The samples were then embedded in paraffin wax, cut into 10-μm sections and stained with Masson's trichrome. Teratoma-derived DNA was prepared from fixed tissue using the DNeasy Blood and Tissue kit (Qiagen) and PCR amplified using mouse and pig actin specific primers (Supplementary Table 2) and FastStart Taq DNA polymerase (Roche, Indianapolis, IN).
Animal welfare and ethics
All animal work conformed to guidelines for animal husbandry according to the UK Home Office, and ethics were approved by the Roslin Institute Animal Ethics Committee. Animals were naturally mated and sacrificed under schedule 1 procedures that do not require specific Home Office approval.
Results
PEFs were transduced with four retroviruses expressing cDNAs for Oct4 (O), Sox2 (S), KLF4 (K),and c-Myc (M), three retroviruses O, S, and K (without Myc), or c-Myc alone. The transduced cultures were replated 48 h later, into wells containing mitotically inactivated mouse SNL76/7 feeder cells and one of three different ESC support media: mESC medium containing fetal calf serum and hLIF (FL), serum-free mESC medium containing serum replacement and hLIF (SRL), and hESC medium containing serum replacement and hFGF2 (SRF). In all culture media, transduction with the four factors (OSKM) or c-Myc alone markedly stimulated cell growth. In contrast, OSK or mock transduced cells showed no response. To facilitate clone selection the transduced cells were replated at low cell density and 10 days after the initial retroviral transduction, colonies expressing the stem cell marker alkaline phosphatase were identifiable throughout all wells containing the OSKM transduced cells (Fig. 1A). Closer microscopic inspection of these OSKM cultures revealed that many cells in SRL medium formed compact, coherent three-dimensional colonies, whereas those in SRF grew flatter with less regular morphology (Fig. 1B). Nevertheless, cells of both types stained strongly for alkaline phosphatase (Fig. 1C). In contrast, colonies cultured in FL medium exhibited a more differentiated morphology. Cultures transduced with c-myc alone generated far fewer alkaline phosphatase positive colonies. To establish stem cell lines we picked and expanded colonies from the OSKM, and c-myc transduced cultures in all three growth conditions.
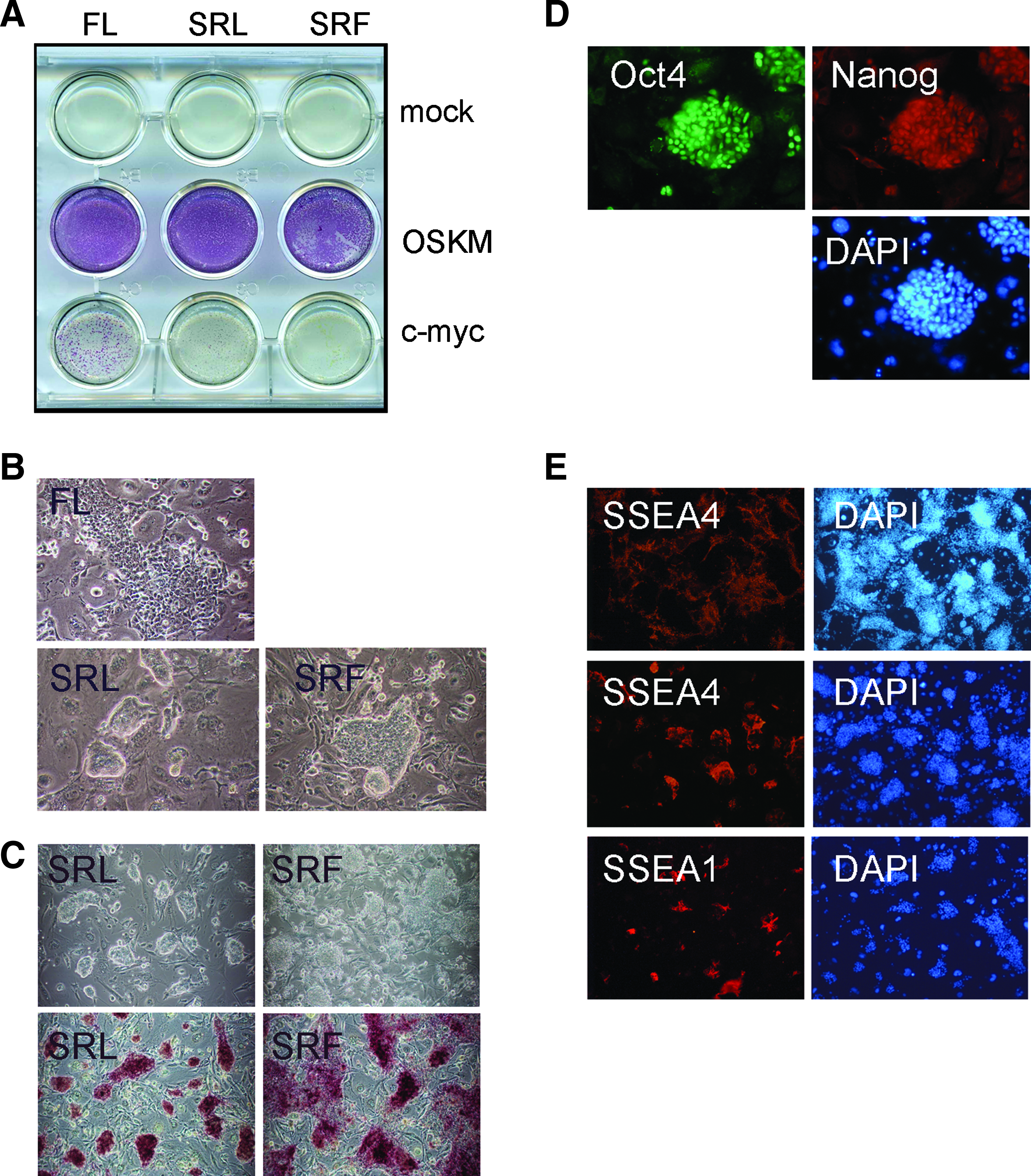
Generation of reprogrammed pig cells. (
During expansion in culture, many OSKM clones derived in the SRL medium retained a compact undifferentiated morphology, reminiscent of mouse ESC/iPSCs. In contrast, SRF lines typically grew flatter and did not form such distinct coherent colonies. The more typical ESC morphology of OSKM SRL cultures indicated that they might represent a more primitive naive stem cell state analogous to mouse ESC. To assess expression of stem cell markers in these putative pig iPS lines, cells were immunostained for the stem cell markers OCT4, NANOG, SSEA-1, and SSEA-4 (Fig. 1D and 1E). Because OCT4 protein could arise from either the mouse Oct4 cDNA or the endogenous pig gene, detection of OCT4 immunoreactivity was not diagnostic of reprogramming, but did nonetheless clearly identify the undifferentiated stem cell-like colonies. Furthermore, many OCT4 positive cells also expressed the nuclear factor NANOG, and cell surface marker SSEA-4 indicating that reprogramming had induced expression of endogenous pig stem cell markers. SSEA-4 was detected in all of the SRL lines screened but expression was variegated, and the extent differed between SRL lines. The cell surface marker SSEA-1, expression of which is associated with naive ESC, was not readily observed on the majority of SRL lines. However, low-level expression was identifiable in some cells of a line (A1) that formed particularly tight, compact colonies, reminiscent of mouse ESC (Fig. 1C and E).
In view of their ESC-like morphology and marker expression we further investigated the extent of molecular reprogramming in SRL cultures by comparing the transcriptional profile of two SRL cell lines with the parental PEF cells by microarray analysis. RNA prepared from PEFs and SRL clones A1 and D3 was converted to cDNA, hybridized to customized Agilent pig arrays, and analyzed by statistical and network-based methods. This analysis showed that fibroblast and mesenchymal-associated genes were downregulated in both reprogrammed clones compared with PEF cells, suggesting significant extinction of the original fibroblast genetic programme, coincident with reprogramming (Fig. 2A). Indeed, downregulation of Snail and transforming growth factor (TGF)-β expression, combined with upregulation of E-cadherin was indicative that reprogramming had involved a mesenchymal to epithelial transition. Importantly, we found that many ESC-associated transcripts were upregulated in both of the reprogrammed pig cell lines, relative to PEFs, pointing to acquisition of a primitive stem cell phenotype. RT-PCR analysis confirmed the upregulation of porcine stem cell markers OCT4, NANOG, SOX2, KLF4, and KLF5 (Fig. 2B). Although, basal expression of some of these genes could be detected in PEF cultures, as previously reported (Ezashi et al., 2009), measurement of NANOG mRNA by quantitative RT-PCR demonstrated that there was significant upregulation of this endogenous stem cell marker, in line with the microarray data. Interestingly, two genes that are specifically enriched in mESC, the orphan nuclear hormone receptor NROB1 (Kim et al., 2008; Nichols and Smith, 2009) and the metabolic enzyme threonine dehydrogenase TDH (Wang et al., 2009), were among the most highly induced genes in the reprogrammed pig cell lines. Other strongly upregulated pig transcripts, for example, EGF, STAT4, NEUROD, and TRKB, are not exclusively ESC/EpiSC markers, but includes genes that have previously have been shown to be upregulated during reprogramming of mouse fibroblasts (Sridharan et al., 2009). Based on their transcription profiles, our reprogrammed pig cells exhibit many features characteristic of ESCs.
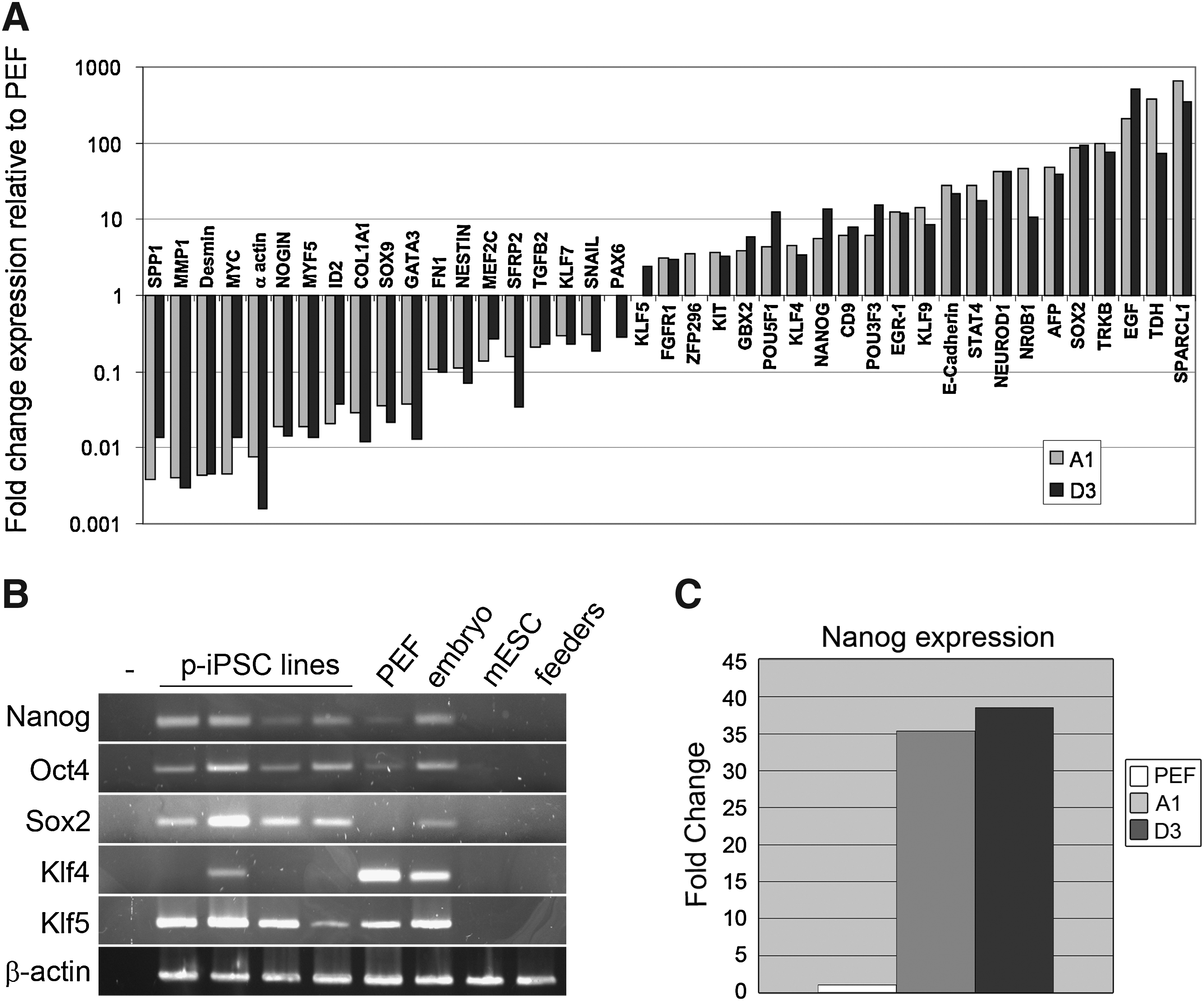
Stem cell profile of the pig cell lines. (
To assess the functional impact of reprogramming on the developmental potency of SRL cells we examined their differentiation potential. SR cell lines cells were injected into SCID mice and tumors were obtained (Supplementary Table 2). DNA-PCR confirmed that the tumors were derived from the pig cells (Supplementary Fig. 1). However, histological analysis of tumor sections revealed a predominantly glandular phenotype with evidence of necrosis and rudimentary differentiation, in a pattern more typically obtained with embryonal carcinoma cells (Supplementary Fig. 1). Thus, although the iPSC-like lines we isolated upregulate many endogenous stem cell markers, they show limited differentiation in vivo. Chromosome counts performed on two cell lines showed that the majority of cells in these cultures contained 38 chromosomes (A1:64%, D3:54%, 50 plates counted), consistent with maintenance of a porcine diploid karyotype (Supplementary Fig. 1). This suggests that restricted differentiation is probably not due to gross karyotypic abnormality or instability. An alternative explanation might be that the pig cells are trapped in a proliferative/self-renewal growth mode in vivo, due to continued expression of the retroviral transgenes, particularly c-myc (Takahashi and Yamanaka, 2006). Indeed, RT-PCR analysis revealed sustained expression of the OSKM transgenes, indicating that they had not been efficiently silenced (Fig. 3A). This criterion, associated with incomplete reprogramming in mouse and human iPSCs, indicates that the pig cell lines are not fully reprogrammed, and may account for their limited differentiation capacity.
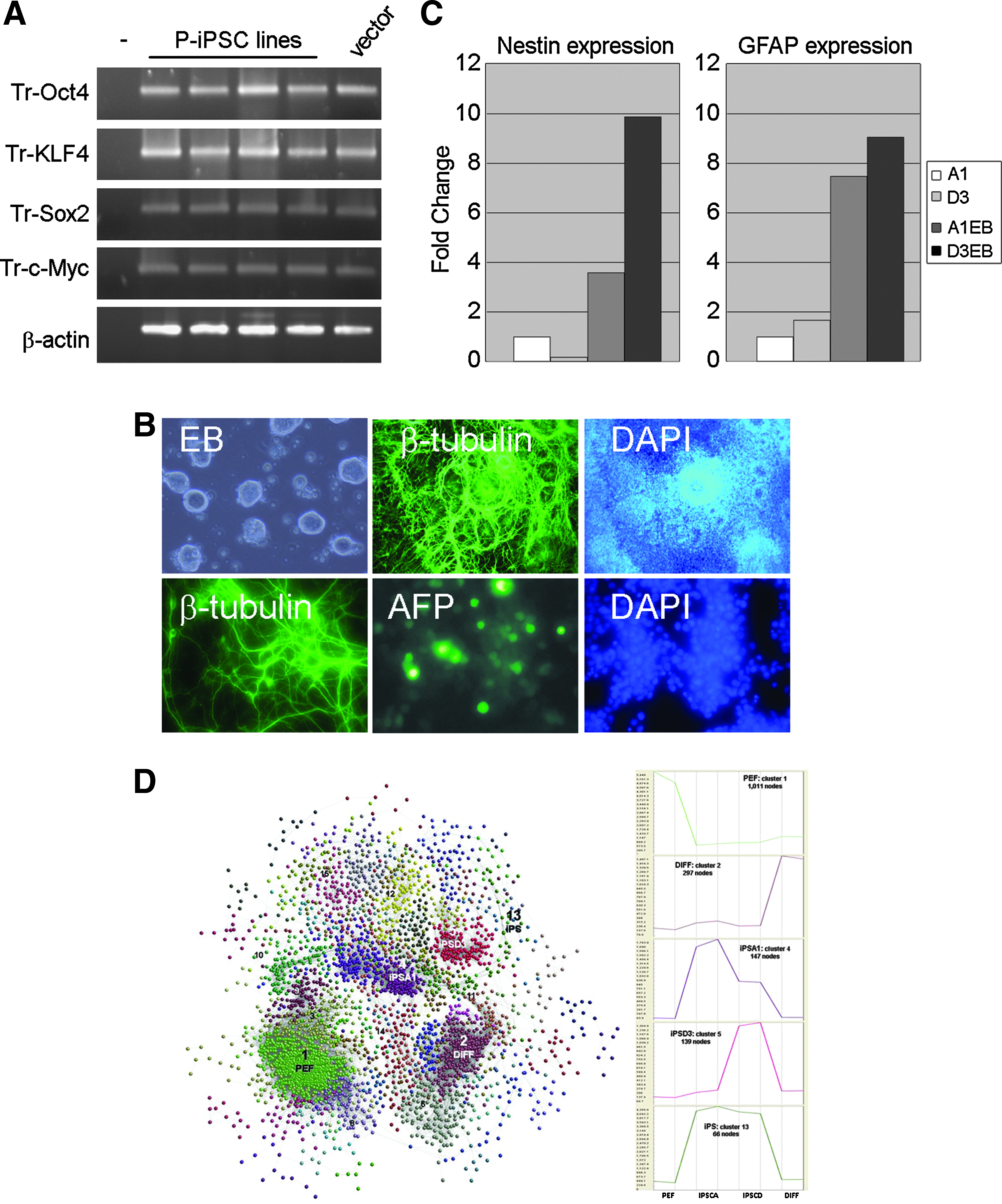
Differentiation of pig cell lines. (
To examine whether the pig cell lines exhibit greater differentiation potential in vitro we cultured several SRL cell lines on nonadherent adherent plates in the presence of FCS to induce EB differentiation. All cell lines readily formed aggregates in suspension (12/12) and upon attachment to gelatinized plates 7 out of 12 generated outgrowths containing cells with diverse differentiated morphologies including obvious neurons. Extensive neural differentiation was confirmed by immunostaining for Class III β-tubulin, and the detection of Nestin and GFAP mRNA expression by quantitative RT-PCR (Fig. 3B and C), indicating the presence of ectoderm derivatives. Endoderm differentiation in some of the differentiation cultures was supported by upregulation of alpha fetoprotein (Fig. 3B and Supplementary Fig. 2). To assess the overall extent of differentiation we compared the mRNA expression profile of differentiated D3 cells (the cell line that produced the greatest diversity of cell morphologies and markers), with undifferentiated D3 and A1 cells, and the parental PEFs. We identified 4223 transcripts that were differentially expressed between the four cell types when the microarray expression profile of each was compared to the other three cell populations. The data from these differentially regulated transcripts were then used to construct a network graph (Fig. 3C) based on the correlation between individual expression profiles using BioLayout Express3D program (Freeman et al., 2007). This cluster analysis confirmed that the largest group of coordinately regulated transcripts were PEF-associated genes downregulated during reprogramming. The second largest group was restricted to differentiated D3 cells but generally not expressed in PEFs. To examine these data further in an unbiased manner, we mapped appropriately annotated PEF and DIFF cluster genes onto an expression atlas of human cell lines (Freeman et al., unpublished). Whereas the PEF enriched transcripts mapped largely to mesenchymal/fibroblast cell types, a number of genes within the differentiation-specific cluster were associated with neuronal cell types. Interestingly, a similar neuronal-associated pattern was also exhibited by two of the reprogramming genes, SOX2 and c-MYC.
In the standard in vitro embryoid body differentiation assay it appears that the SRL lines are capable of differentiating into diverse cell types belonging to germ lineages that are distinct from the original PEFs. This contrasted with SRF lines derived in FGF2, which produced relatively uniform monolayers of cells displaying a singular primitive undifferentiated morphology in the EB differentiation protocol (Supplementary Fig. 2). In light of this difference, we were interested to determine if LIF, a key signaling factor in culturing rodent ES cells, might contribute to supporting the stem cell characteristics of the pig SRL cells.
To determine whether SRL cell lines respond to LIF we first examined activation of its key downstream effector molecule STAT3 and one of its likely target genes SOCS3. To eliminate any cellular or paracrine contribution from feeder cells, pig cells were plated on laminin, without feeder support. The cultures were starved of growth factor stimulation for 24 h and then stimulated with murine and human LIF before being lysed for protein analysis. Western blotting for Stat3 tyrosine phosphorylation revealed a strong activation of STAT3 in response to human LIF and a lesser response to murine LIF (Fig. 4A). To ascertain if this increase in STAT3 phosphorylation correlated with increased STAT3 target activity we examined expression of SOCS3 gene by RT-PCR in pig cells that were either starved of LIF, stimulated for 2 h with LIF, or grown continuously in LIF. Expression of porcine SOCS3 was upregulated in pig cells following both acute and sustained exposure to LIF (Fig. 4B). These data suggest that the LIF signalling pathway is active in the reprogrammed pig cell lines.

Reprogrammed pig lines are responsive to LIF. (
Because LIF/STAT3 signaling plays a key role in regulating self-renewal of mouse ES cells, we wanted to determine if the pig cell lines also exhibited a biological response to LIF. To assess their functional response to LIF, OSKM SRL cell lines were plated at low density on laminin-coated wells and cultured for a week in the presence or absence of LIF. Staining these cultures for the stem cell marker alkaline phosphatase (AP), revealed that the majority of lines tested (7/10) showed increased AP staining in response to LIF (Fig. 4C). The other three lines grew poorly on laminin regardless of the presence of LIF. Titration of LIF concentration on two representative LIF-dependent cell lines demonstrated that this response was dose dependent. (Fig. 4D). Microscopic inspection of the cultures indicated that reduced levels of AP staining in the absence of LIF was largely due to poor cell growth in these conditions rather than downregulation of AP activity resulting from differentiation. To test this directly we assayed growth of pig iPSC lines by measuring the cumulative increase in DNA content of cultures. The parental pig embryonic fibroblasts, or cells transduced with c-myc retrovirus alone, showed little or no response to LIF. In contrast, OSKM SRL cell lines exhibited a dramatic increase in growth when stimulated with LIF (Fig. 4E). Based on colony morphology, gene expression profile and response to LIF, it appears that OSKM reprogramming in these pig cell lines has induced a stem cell-like state that share markers and growth characteristics with mouse ESCs.
Discussion
Pluripotent pig stem cells have important applications in modelling embryonic development and disease processes, both in agricultural and biomedical research (Hall, 2008; Telugu et al., 2010b) and so we have used direct reprogramming to generate iPSC-like pig stem cells (Takahashi and Yamanaka, 2006). Although it appeared that reprogramming was incomplete, factor-induced pig cells formed ESC-like colonies, expressed endogenous stem cell markers and acquired novel developmental potential. Interestingly, the cells displayed a functional growth response to LIF, a cytokine that promotes self-renewal of naïve rodent ESCs, indicating that reprogramming may have uncovered a basic stem cell signaling network that is latent in mammalian ESCs (Nichols and Smith, 2009).
Most iPSCs reported to date can be assigned to one of two identifiable stem cell states, either an early “naive” pluripotent stem cell state analogous to rodent ESCs or a more mature “primed” ESC state similar to EpiSC and hESCs. We found that reprogramming pig fibroblasts in the presence of LIF, but without serum (SRL), generated clones that displayed growth characteristics reminiscent of mouse ESCs, and distinct from previously reported pig iPS cell lines (Esteban et al., 2009; Ezashi et al., 2009; Wu et al., 2009). The compact, refractile, colony morphology of many SRL clones, and upregulated expression of the orphan receptor NROB1, were compatible with conversion to a naive-type stem cell (Brons et al., 2007; Nichols and Smith, 2009; Tesar et al., 2007). However, the lack of expression of the early epiblast gene Rex1, indicated that the reprogrammed pig cells do not fully align with either one of the standard ESC states, as defined in other species. Although this indeterminate state may arise through incomplete reprogramming, heterogeneity within the pig iPS cultures may also be a contributory factor. The latter would be consistent with variation in SSEA-1 and SSEA-4 expression we observed in our cultures, and may explain variability in expression of these markers between previously reported pig iPS cell lines (Alberio et al., 2010; Esteban et al., 2009; Ezashi et al., 2009; Montserrat et al., 2011; Telugu et al., 2010a; Wu et al., 2009). Regardless of their exact status, gene expression profiling of our pig iPS cells revealed widespread extinction of the original primary fibroblast program and substantial upregulation of many endogenous ESC-associated genes, pointing to acquisition of developmental potency as a result of reprogramming.
Reestablishment of pluripotency is an essential feature of iPS cells, and although our pig iPS-like cells exhibited limited differentiation in vivo, evidence from in vitro experiments indicated that the SRL cell lines could give rise to derivatives of all three embryonic germ layers. The most clearly identifiable of these, based on morphology and marker expression, were neuronal cells, confirming that the reprogrammed pig fibroblasts had achieved a multipotent stem cell state. However, continuous expression of the retrovirus-driven reprogramming factors in the pig cells may have directly contributed to restricted differentiation, as well as highlighting the failure to silence the retroviral promoters, a characteristic associated with incomplete reprogramming (Hotta and Ellis, 2008). Although partially reprogrammed pre-iPS cell lines have previously been reported for mouse, expression of stem cell markers such as NANOG in the undifferentiated pig cells, and their demonstrated capacity for differentiation in vitro, suggests that the pig stem cells have progressed beyond the equivalent transitional stage (Mikkelsen et al., 2008; Silva et al., 2008; Sridharan et al., 2009). Moreover, whereas suppression of pro-differentiative signals using chemical inhibition of the MAPK and GSK3β pathways (2i medium), facilitate the completion of reprogramming in mouse pre-iPS cells (Silva et al., 2008), our pig stem cell cultures degenerated after repeated passaging in similar medium, producing cultures largely consisting of differentiating neural cells (Thomson et al., unpublished). Taken together, it seems likely that the SRL cells represent a multipotent stem cell state, that depending on growth conditions, can participate in aspects of embryonic differentiation programs.
Continuous expression of the reprogramming factors in the pig cell lines described here, and in previous reports, may imply that maintenance of the undifferentiated pluripotent state in pigs, in contrast to mouse and human, relies on input from the exogenous transcription factors (Esteban et al., 2009; Ezashi et al., 2009; Montserrat et al., 2011; Telugu et al., 2010a, 2011; Wu et al., 2009). Derivation of authentic stable pig ESC lines from blastocysts has not yet been reported, and in a recent study, forced expression of KLF4 and Oct4 in embryos was required to facilitate the derivation of stem cell lines (Telugu et al., 2011). On this basis, it appears that the endogenous regulatory networks that control pluripotency in pig cells are unstable in current culture regimes, and enforced activity of stem cell transcription factors is required to prevent differentiation (Wu et al., 2009). Although this antidifferentiative effect has been overcome by applying standard in vitro and in vivo differentiation protocols (Esteban et al., 2009; Ezashi et al., 2009; Montserrat et al., 2011; Telugu et al., 2010a, 2011), continuing expression of exogenous factors might be expected to interfere with differentiation. The recent suggestion that Oct4 and Sox2 can contribute to early stages of germ lineage selection (Thomson et al., 2011), implies that imbalances in reprogramming factors might affect iPS differentiation. Indeed, overexpression of Sox2 has been shown to direct differentiation toward neurectoderm (Zhao et al., 2004), and this factor was among the most highly upregulated pluripotency genes detected in our microarray analysis.
Forced expression of reprogramming factors in ESCs can change their growth characteristics and stem cell status (Guo et al., 2009; Hanna et al., 2010 ; Silva et al., 2009). We found that reprogrammed pig SRL cells not only exhibited characteristics of naive-type ESC, but also acquired a functional response to the cytokine LIF. In mouse ESC, the response to LIF is thought to derive from its role in vivo, maintaining pluripotency in the embryo during delayed implantation (Nichols et al., 2001). Because this response is not normally required during uninterrupted embryo development, the LIF signaling pathway primarily plays a cryptic, regulative role in this context, and evidence for a LIF-dependent response in ESCs from species other than rodents is currently limited. Indeed, human ESC/iPSCs do not normally exhibit a functional response to LIF. However, constitutive expression of reprogramming factors in combination with 2i inhibitors can provoke human stem cell lines to adopt a mESC-like morphology and growth characteristics, including self-renewal in response to LIF (Buecker et al., 2010; Hanna et al., 2010). It is possible that the LIF response we observe in SRL cells could arise from continuing activity of the retroviral transgenes. In addition, activation of STAT3 by LIF during the reprogramming process may aid progression to an ESC-like state, as has been recently demonstrated in mouse cells (Yang et al., 2010). Indeed, LIF efficiently activated STAT3 signaling in PEF cultures and has been reported to support the growth of pig epiblast cells in culture (Wianny et al., 1997).
The observed acquisition of a LIF response through reprogramming in human and pig cells, supports the existence of a latent regulatory pathway in all mammals which, if accessed appropriately, could be used to establish pluripotent cell lines analogous to “naive” mouse ES cells from any mammal (Buecker et al., 2010; Hanna et al., 2010; Telugu et al., 2010a). In contrast to these recent reports, our reprogrammed pig cells did not require additional chemical manipulation of either signaling pathways or chromatin modifiers to acquire a functional LIF response and some naive ESC characteristics. Indeed, our preliminary experiments indicate that simple translation of the chemical inhibitor strategies successfully utilized to derive mouse and human naive ESCs do not necessarily stabilize the stem cell status of our reprogrammed pig cells (Thomson et al., unpublished). An important challenge for the future will be to investigate how manipulation of LIF/STAT3 signaling in conjunction with other regulatory pathways can be used to regulate stem cell states and ultimately generate authentic pig ESC cells.
Footnotes
Acknowledgments
This work was supported by a Biological and Biotechnological Research Council Institute Strategic Programme Grant, and the Roslin Foundation. T.B. and A.T. thank Tim King and John Bracken at the Roslin Large Animal Unit for their help in providing pig tissues, and Denis Headon and Michael Clinton for comments on the manuscript. The SSEA-1 and -4 antibodies developed by Solter and Knowles were obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by The University of Iowa, Department of Biology, Iowa City, IA 52242.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.