
Abstract
Abstract
Cell death by apoptosis is considered to be irreversible. However, reports have indicated that its reversibility is possible if the cells have not yet reached the “point of no return.” In order to add new information about this topic, we used cells at different moments of apoptotic process as nuclear donors in somatic cell nuclear transfer (SCNT) in order to test if programmed cell death can be reversed. Adult bovine fibroblasts were treated with 10 μM of staurosporine (STP) for 3 h and analyzed for phosphatidylserine externalization (Annexin assay) and presence of active caspase-9. Annexin-positive (Anx+) and Caspase-9-positive (Casp-9+) cells were isolated by FACS and immediately transferred into enucleated in vitro matured bovine oocytes. After STP treatment, 89.9% of cells were Anx+ (4.6% in control cells; p<0.01) and 24.9% were Casp-9+ (2.4% in control cells; p<0.01). Fusion and cleavage were not affected by the use apoptotic cells (p>0.05). Also, the use of Anx+ cells did not affect blastocyst production compared to control (26.4% vs. 22.9%, respectively; p>0.05). However, blastocyst formation was affected by the use of Casp-9+ cells (12.3%; p<0.05). These findings contribute to the idea of that apoptosis is reversible only at early stages. Additionally, we hypothesize that the “point of no return” for apoptosis may be located around activation of Caspase-9.
Introduction
A well-studied phenomenon observed during SCNT procedures is the potential for remodeling and reprogramming of donor cell chromatin by an ooplast (enucleated oocyte). The cytoplasm of a matured oocyte seems to be capable of reprogramming and remodeling chromatin even in unconventional situations, as has been shown in xenotransplantation when a donor nuclei and ooplast were combined from different species (Dominko et al., 1999), when nuclei were derived from lyophilized cells (Das et al., 2010), from cells exposed to overheating (Loi et al., 2002) and even from cells derived from a section of tissue that was inadequately frozen without cryoprotectors (Wakayama et al., 2008).
Based on the great potential for an enucleated, matured oocyte to reprogram somatic cells even under the critical conditions mentioned above, we recently proposed a novel approach in order to test oocyte's remodeling potential. We have shown that the use of anx-positive cells (sorted by FACS) as nuclear donors for SCNT negatively affected embryonic development, total blastocyst cell number, and increased the apoptotic rate in developing SCNT embryos. However, despite the reduction in blastocyst formation rate and quality, one live-born calf was obtained from an apoptotic cloned cell after the cloned embryo was transferred to a recipient (Miranda Mdos et al., 2009).
Apoptosis is a type of programmed cell death characterized by specific stages that are associated with several physiological processes in pluricellular organisms (Kerr et al., 1972; Kroemer et al., 2009). In a simplistic view, apoptosis may be divided into four stages (Morita and Tilly, 1999). The first stage is the exposure of a cell to a potentially lethal stimulus (e.g., UV radiation, drug treatment, etc.). The second stage of apoptosis is the recruitment of intracellular signaling molecules, primarily the Bcl-2 family of proteins responsible for translating several kinds of death stimulatory information into a central regulatory checkpoint for interpretation, which is the third stage. It is in the third stage that proteins such as cytochrome c and apoptosis induction factor (AIF) play important roles. The fourth stage is characterized by the activation of effector or “executioner” proteins (e.g., caspases) or AIF itself in the case of caspase-independent apoptosis (Susin et al., 1999).
In our previous study, serum starvation (0.5% for 24 h) was chosen as a donor cell cycle synchronization protocol for SCNT, and the number of apoptotic cells (∼15%) in the donor cell population was detected by Annexin assay, sorted by FACS and used for cloning. However, we conjecture that serum starvation triggered an asynchronous apoptotic stimulus, which made it impossible to evaluate in which apoptotic stage the donor cells were when they were introduced into cytoplasts. Therefore, in this study we used staurosporine (STP) to induce the apoptotic process in bovine fibroblasts, then used the synchronized fibroblasts as nuclear donors for cloning at different phases of the apoptotic process prior to internucleosomal DNA fragmentation. We show that the use of anx-positive cells as nuclear donors has no effect on embryo production; however, the use of caspase 9-positive cells hampers the development of cloned embryos after SCNT. These results contribute to the hypothesis that apoptotic cells contain a “point of no return,” and suggest that this critical juncture is located after caspase 9 activation, but before nuclear fragmentation.
Materials and Methods
Unless otherwise noted, chemicals and growth media were purchased from Sigma (St. Louis, MO). Oocytes, embryos, and fibroblast cultures were maintained at 38.5°C in an atmosphere of 5% CO2 in air at maximum humidity.
Primary cell isolation and culture
Bovine skin fibroblasts were obtained from an adult Nelore female (Bos indicus) by ear-punch biopsy. After hair removal and rinsing in 70% ethanol, the tissue was washed in Dulbecco's Phosphate-Buffered Saline (DPBS), minced thoroughly with small scissors and placed on plastic Petri dishes. The cells were allowed to dry partially to increase attachment to the surface of the plastic, and the culture dish was then carefully filled with Dulbecco's modified Eagle's medium (DMEM; Gibco Laboratories, Gaithersburg, MD) supplemented with 10% fetal bovine serum (FBS; Gibco Laboratories) and 50 mg/mL gentamicin. Cells were regularly passaged when they approached confluence. Cells were utilized between passages 3 and 6 for all experiments in this study.
Cell cycle synchronization and analysis
The cell cycle of donor cells was synchronized by serum starvation for 24 h according to our previously reported protocol (Miranda Mdos et al., 2009), which synchronized ∼80% of the cells to G1/G0 of the cell cycle. Briefly, cells were seeded on 75-cm2 plastic flasks (TPP, Trasadingen, Switzerland) at a concentration of 4×104 cells/mL in DMEM supplemented with 10% FBS, which lead to greater than 60% confluency. After 24 h, the medium was changed to DMEM supplemented with 0.5% FBS (serum starvation) and maintained for an additional 24 h. After this period, cells were subjected to experimental treatments.
Induction and time course of apoptosis
After cell cycle synchronization by serum starvation, cells were treated with 10 μM staurosporine for 0 (untreated control), 3, 12, and 24 h. After each time point, cells were trypsinized and divided into two samples for incubation with either Annexin V (Anx) conjugated to FITC (Apoptest-FITC; Dako, Hoeven, The Netherlands) at a dilution of 1:100 in the calcium containing binding buffer provided with the kit or with a caspase 9 detection kit (FLICA Apoptosis Detection Kit Caspase Assay; Immunochemistry Technologies, LLC; Bloomington, MN) according to the manufacturer's recommendations.
After incubation with either Anx or caspase 9, propidium iodide (PI) was added at a final concentration of 5 mg/mL to detect necrotic cells. The data was collected using a FACSAria flow cytometer (BD, Franklin Lakes, NJ) excited at 488 nm and analyzed using the FACSDiva software application (Becton Dickinson, Fullerton, CA). The cell populations were observed in a “dot plot” graph that discriminated: live cells (population of Anx or Casp-9-negative/PI-negative cells); apoptotic cells (Anx or Casp-9-positive/PI-negative cells); necrotic cells (Anx or Casp-9-negative/PI-positive cells); late apoptotic/necrotic cells (Anx or Casp-9-positive/PI-positive cells). Cell samples from each time point were stained with 5 μg/mL Hoechst 33342 for 10 min in order to evaluate signs of chromatin fragmentation, which indicates completion of the apoptotic process.
Assessment of apoptosis in donor cells for cloning by JC-1 and TUNEL assay
Loss of mitochondrial membrane potential (ΔΨm) in bovine fibroblasts undergoing apoptosis (to be utilized as nuclear donors for SCNT) was detected by using MitoProbe™ JC-1 Assay Kit (Invitrogen, Eugene, OR). JC-1 (5,5′,6,6′-tetrachloro-1′,3,3′,tetraethylbenzimidazolocarbocyanine iodide) has been widely used as a probe for analysis of mitochondrial membrane potential. In apoptotic cells, JC-1 enters the cytoplasm as a monomer, emitting green fluorescence, but in healthy cells it is taken up by the mitochondria and forms JC-1 aggregates that emit red fluorescence (Cossarizza et al., 1993). After STP treatment, cells were stained with 2.5 μg/mL of JC-1 in DMEM with 0.5% FBS for 10 min at room temperature. Cells were then washed in DMEM with 0.5% FBS and analyzed under a fluorescent microscope and by a BD FACSAria flow cytometer. During flow cytometric analysis, the ratio of the fluorescence at 590 nm (red fluorescence), which is proportional to the ΔΨm, was compared to the fluorescence at 538 nm (green fluorescence), which is proportional to the mitochondrial mass. This analysis was performed in triplicate.
Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) was utilized for detecting DNA fragmentation (In Situ Cell Apoptosis Detection Kit; Roche Diagnostics, Mannheim, Germany). Briefly, adult bovine fibroblasts cells were seeded onto coverslips and treated as previously described with 10-μM STP for 3 h. Control cells without STP treatment were included in all analyses. For TUNEL staining, STP-treated cells were washed in PBS containing 1 mg/mL polyvinyl alcohol (PBS–PVA), and fixed for 1 h at room temperature with 3.7% (w/v) paraformaldehyde diluted in PBS. After fixation, the cells were washed three times in PBS–PVA, permeabilized in PBS containing 0.5% (v/v) Triton X-100 and 0.1% (w/v) sodium citrate for 1 h at 4°C, washed three times in PBS–PVA again, and incubated in TUNEL reaction solution for 1 h in a humidified chamber at 37°C. Positive controls were incubated in 1000 IU deoxyribonuclease I/mL (DNase I; Sigma) for 1 h at 37°C and washed twice in PBS–PVA before TUNEL staining. Negative controls were incubated in fluorescein–dUTP in the absence of TUNEL assay enzyme (TdT).
FACS and nuclear transfer of apoptotic cells
Once the time course for the two apoptotic markers was evaluated and characterized, and occurrence of apoptosis was confirmed, cells treated with 10 μM of staurosporine for 3 h were stained with either Annexin or Caspase-9 (separately), sorted and utilized for nuclear transfer. Anx or Casp-9-positive cells (1000–3000 cells) were sorted by FACS into room temperature DMEM supplemented with 10% FBS containing a physiological concentration of calcium. Cells were then centrifuged and immediately used for nuclear transfer, with less than 1 h elapsing between cell sorting and the fusion step of SCNT. In all experiments, a group of the untreated unstained cells, which were cultured and synchronized as described above, were used to calibrate the fluorescent background and sorted for SCNT (control group). Gates were designed within quadrants to exclude fluorescent overlap and increase the purity of sorted cells.
The nuclear transfer procedures were performed as previously described (Miranda Mdos et al., 2009). Briefly, donor cell preparation consisted of serum starvation (DMEM with 0.5% FBS) for 24 h prior STP treatment and each round of micromanipulation. Bovine oocytes aspirated from 3 to 6 mm follicles of slaughterhouse-derived ovaries were selected by morphology (de Loos et al., 1989) and matured in vitro. After 18 h, oocytes were denuded and selected for the extrusion of the first polar body. Oocytes were enucleated by micromanipulation, and the donor cell was placed into the perivitelline space of each enucleated oocyte. SCNT couplets were then electrically fused at 24 h after maturation, chemically activated with 5 μM ionomycin for 5 min and 2 mM of N-6 dimethylaminopurine (6-DMAP) for 3 h and then cultured in SOF medium supplemented with 10% FCS in an atmosphere of 5% CO2 in air for 7 days. Fusion rates were evaluated during the SCNT procedure; cleavage rates were evaluated at the third day of culture, and blastocyst rates were evaluated at the seventh day. Parthenogenetic embryos were activated and in vitro cultured under the same conditions and were used as a control for oocyte quality, activation procedure, and in vitro culture.
Results
Induction and time course of apoptosis
STP concentration (10 μM) was optimized in a dose–response experiment where the dosage showing the highest late apoptotic rate together with lowest necrotic rate after 24 h of treatment was considered as the apoptotic specific concentration (data not shown). To investigate the time course of the apoptotic process, bovine fibroblasts were treated with 10 μM staurosporine for 3, 12, or 18 h and then screened for Anx and Casp-9 apoptotic markers. This experiment was repeated three times and the results are summarized in Figure 1.
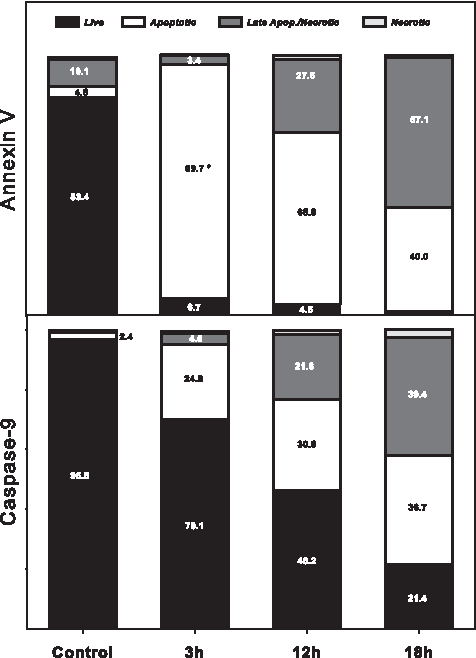
Time course for Annexin and Caspase-9 staining in bovine fibroblasts. Cells were treated with 10 μM of STP for 3, 12 and 18 h, then stained either with Anx/PI or Caspase-9/PI and analyzed by FACS as described and showed in Figure 4. The percentage of apoptotic cells (Anx-positive) increased dramatically after 3 h of STP treatment (*indicates statistical difference in comparison to control; p<0.05). For Caspase-9, a gradual decrease in the percentage of live cells according to treatment length was observed. Three hours after STP treatment, a significant increase in Caspase-9-positive cells was observed.
After 3 h of staurosporine treatment, the number of early apoptotic cells (Anx-positive/PI-negative) was increased when compared to untreated control cells (89.8% vs. 4.6%), whereas only 24.9% of cells were in the effector phase of apoptosis (caspase-9-positive/PI-negative cells). Necrotic and late apoptotic cells showed no increase at this time point for either marker, ranging from 0 to 4.3% (data not shown).
After 12 h of staurosporine treatment, a reduction in Anx-positive apoptotic cells was observed (65.9%), concomitant with an increase in the Casp-9-positive apoptotic cell population (30.6%). PI staining was detected in the late apoptotic stage 12 h after treatment for both Anx (27.6%) and caspase-9 (21.6%) stained cells. At 18 h after staurosporine treatment, the percentage of late apoptotic cells (57.7%) surpassed the percent of early-apoptotic cells (39.0%). The percentage of Casp-9-positive cells reached a similar value to that of late apoptotic Casp-9 cells (36.7% and 39.4%, respectively). Interestingly, after 18 h of treatment, both Anx and caspase-9 apoptotic cell populations reached similar values (40.0% and 36.7%, respectively).
Cell nucleus fragmentation analyzed by Hoechst 33342 confirmed that the apoptotic process was completed 6 h after staurosporine treatment, resulting in nuclei showing a typical apoptotic fragmented nuclear pattern in nearly all of the cells analyzed (data not shown). Therefore, apoptotic cells were used as nuclear donors 3 h after STP treatment to observe the continuation of the apoptotic process after SCNT prior to nuclear fragmentation.
Assessment of apoptosis in donor cells for cloning by JC-1 and TUNEL assay
To evaluate the cytoplasmic and nuclear status of fibroblasts at the moment of nuclear transfer (NT), a sample of cells were plated onto coverslips, treated with 10 μM of STP for 3 h and analyzed for mitochondrial potential and DNA fragmentation through JC-1 and TUNEL assays, respectively.
Results showed that at the moment of SCNT, cells treated with staurosporine exhibited typical apoptotic morphological changes at both the cytoplasmic and nuclear levels. Signs of nuclear fragmentation, determined by Hoechst 33342 staining, showed a marked change in nuclear morphology typical of apoptosis (nuclear shrinkage), but internucleosomal breaks were not observed by the TUNEL assay (Fig. 2). These results indicate that staurosporine treatment for 3 h did not induce internucleosomal breaks. Thus, cells used for cloning after 3 h of staurosporine treatment were considered to have no fragmented DNA. Concerning membrane potential, cytometry results showed that ΔΨm (quantified by red/green ratio of JC-1 dye) was dramatically reduced by STP treatment (∼70% reduction) in comparison to control cells (Fig. 3; p<0.05).
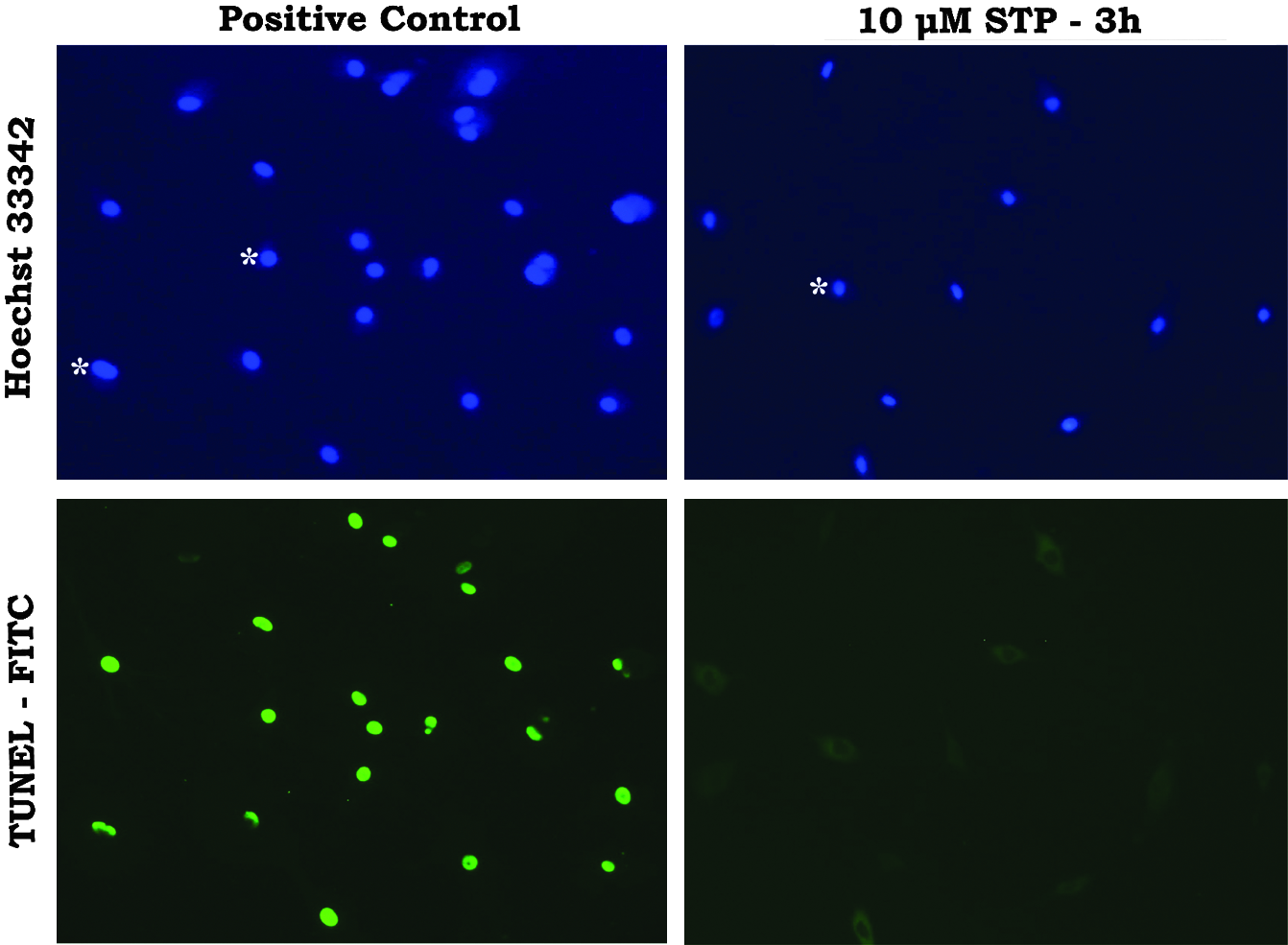
Analysis of DNA shrinkage and fragmentation in bovine adult fibroblasts treated with staurosporine (STP). Fibroblasts were treated with 10 μM of STP for 3 h and submitted to TUNEL assay (nuclei in green). Hoechst 33342 (5 μg/mL) was utilized for counterstaining (nuclei in blue). Results indicated that adult bovine fibroblasts treated with 10 μM of STP for 3 h showed signs of nuclear shrink (asterisks) but did not show signs of DNA fragmentation. Positive control group (incubated with DNAse I prior to TUNEL assay) was prepared with untreated fibroblasts which Hoechst staining shows no signs of nuclear shrink (asterisks). Negative TUNEL control cells (not shown) were incubated without TUNEL assay enzyme (TdT) (200× magnification).
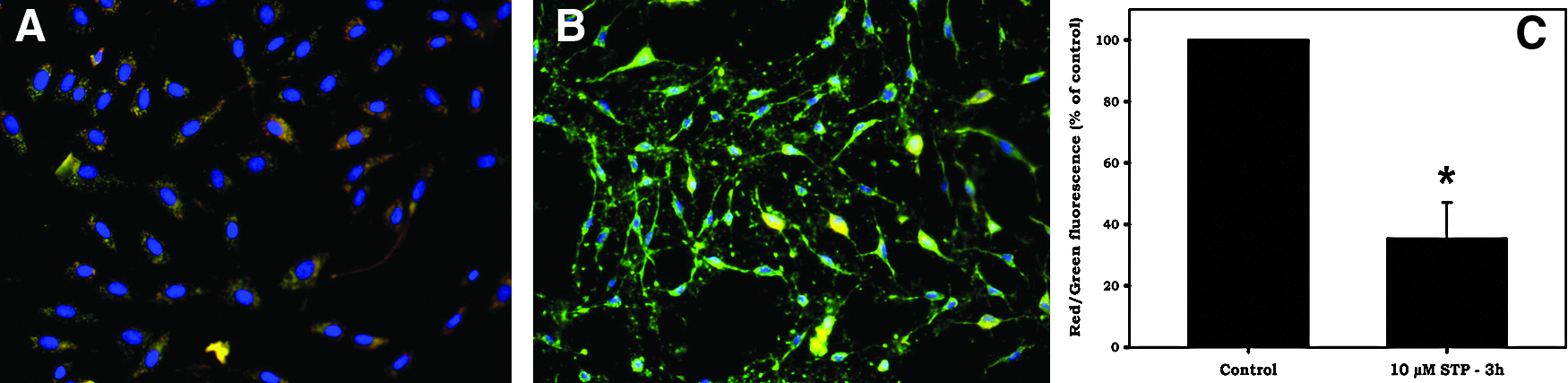
Mitochondrial membrane potential (ΔΨm) of adult bovine fibroblasts treated with STP for 3 h. Cells were stained with JC-1 and ΔΨm was analyzed by fluorescence microscopy and flow cytometry. In healthy untreated cells (
FACS and nuclear transfer of apoptotic cells
As mentioned previously, cells treated with 10 μM of STP for 3 h were separated and recovered by flow cytometry according to apoptotic markers (Anx-positive or Casp-9-positive) and used as nuclear donors for SCNT (Fig. 4).
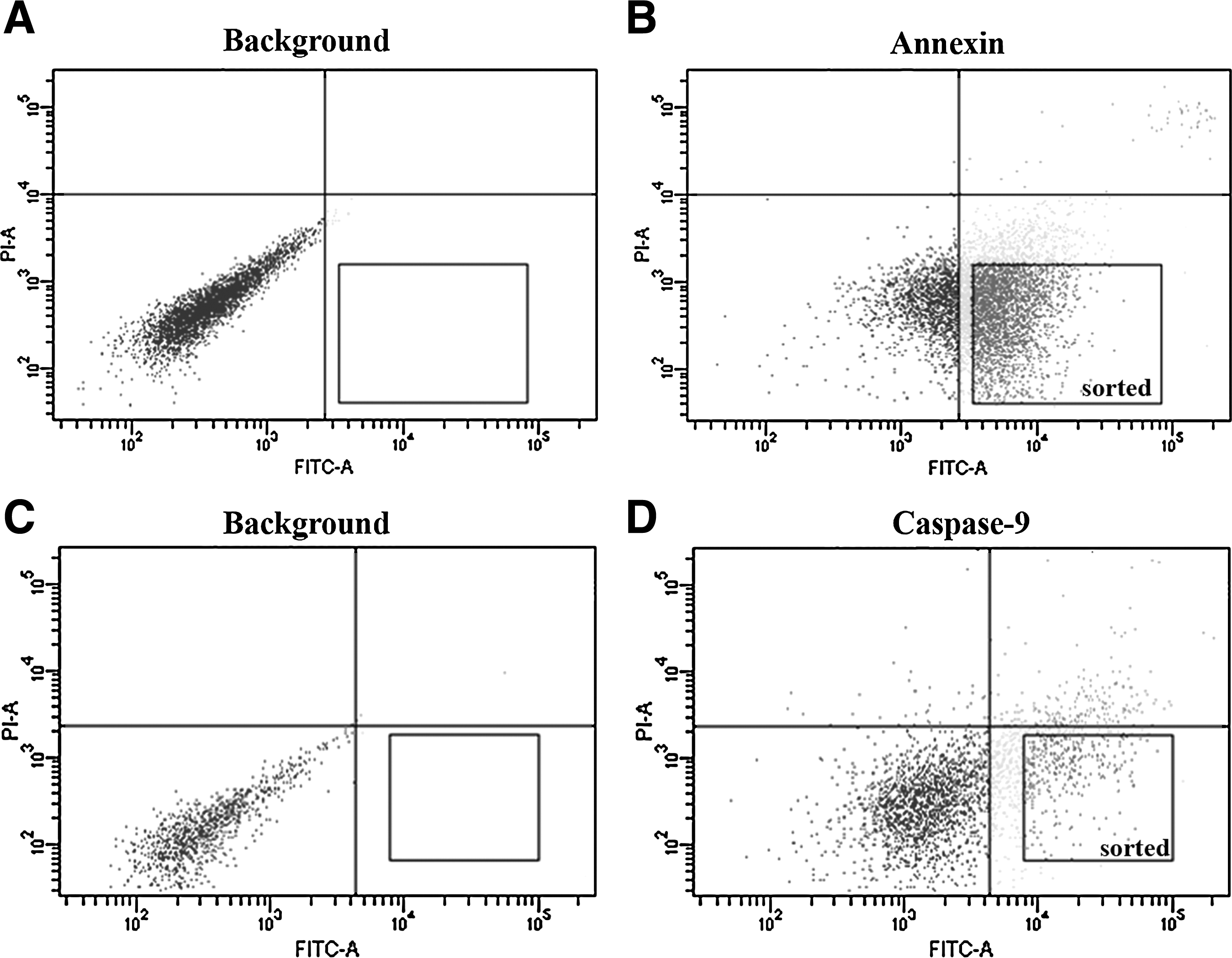
Flow cytometry dot plots showing cell populations, quadrants and sorted cell population after STP treatment. (
Data on the development of embryos reconstructed with Anx-positive or Caspase-9-positive cells are shown in Table 1. Two hundred sixty-two oocytes were micromanipulated in three experimental replicates. Fusion rate was not affected by the use of either Anx-positive or Casp-9-positive apoptotic cells compared to unsorted and untreated control cells (p>0.05). Similarly, no effect on cleavage rates was observed after two days in culture (p>0.05). However, the use of Caspase-9-positive cells as nuclear donors for cloning decreased blastocyst development in comparison to Anx-positive cells (12.3 vs. 26.4%; p<0.05) and control cells (12.3 vs. 22.9%; p<0.05). Anx-positive cells showed similar results to the control (p>0.05).
Different superscripts in the same column mean statistical difference (ANOVA; p<0.05).
Discussion
There are two well-established apoptotic pathways: the extrinsic death receptor-mediated pathway and the intrinsic mitochondrial-mediated pathway (Feng and Kaplowitz, 2005). In the intrinsic pathway, loss of ΔΨm induces cytochrome c release into the cytoplasm, which then binds to apoptotic protease activating factor 1 (APAF-1), resulting in recruitment of pro-caspase-9 by APAF-1. APAF-1 then serves as a scaffolding protein to bring pro-caspase-9 molecules together to promote self-activation of pro-caspase-9 to active caspase-9 in the presence of dATP (Li et al., 1997). Active caspase-9 then cleaves downstream executioner caspases, such as caspase-3 (Kuwana et al., 1998).
According to Yoshida et al. (1997) and Chae et al. (2000), staurosporine is a potent inhibitor of protein kinase C, which is capable of blocking cell cycle progression when used in low concentrations, as well as quickly inducing programmed cell death (PCD) in a variety of cell lines, including fibroblasts, when used at high concentrations (0.1–10 μM). STP triggered intrinsic pathway apoptotic stimuli in most cell lines through phosphatidyl serine exposure and caspase-9 activation, particularly in cancer cell lines (Andersson et al., 2000; Feng and Kaplowitz, 2002; Mannherz et al., 2006; Samraj et al., 2007). In this study, treatment of bovine fibroblasts with 10 μM of STP for 24 h induced phosphatidyl serine exposure at the external face of the plasma membrane (Anx staining), reduced ΔΨm, activated caspase-9, and demonstrated a typical chromatin fragmentation pattern by Hoechst staining, all indicating that the intrinsic pathway of apoptosis was activated.
In this study, the time course of phosphatidyl serine exposition (Anx-staining) and intrinsic pathway activation (active caspase-9 staining) were monitored at 3, 12, and 18 h after STP treatment. These experiments indicated that 3 h after STP treatment most of the cells were apoptotic (∼90% Anx-positive and ∼25.0% caspase-9-positive) with concomitant loss of ΔΨm, typical chromatin condensation, but no internucleosomal DNA fragmentation as evaluated by TUNEL staining. At 12 and 18 h after STP treatment, typical morphological changes, DNA fragmentation and internucleosomal cleavage were detected by Hoechst and TUNEL staining. These results are in agreement with other studies in which the apoptotic time course was monitored after artificially induced PCD, confirming that phosphatidyl serine (PS) exposure is an initial event in the apoptotic process leading to more advanced stages, and that caspase activation is a downstream event in comparison to PS exposure (Clodi et al., 2000; Span et al., 2002).
An interesting observation was made when the percentage of cells positive for apoptotic markers was compared to the percentage of cells with nuclear fragmentation signals (Hoechst staining) 12 h after STP treatment. Although 100% of nuclei showed a fragmented pattern at this time point, only 52.2% of cells were positive for active caspase-9 (taking together apoptotic and late apoptotic/necrotic groups) (Fig. 1). At the same 12-h time point, around 93.5% of cells were positive for annexin (taking together apoptotic and late apoptotic/necrotic groups) (Fig. 1). Studies have shown that in other cell types, the mechanism of death induced by STP can occur independently of caspase-9 activation through a mechanism called AIF (apoptosis induction factor). AIF has been well described in the literature, and its expression has been shown to lead to the externalization of phosphatidyl serine in the cell membrane (Deas et al., 1998; Dumont et al., 2000; Susin et al. 1999). This finding may help to explain why we observe universal nuclear fragmentation but only ∼50% active caspase-9.
Previously, our group reported that naturally occurring Anx-positive fibroblasts negatively affected SCNT embryo production (Miranda Mdos et al., 2009). We hypothesized that this negative effect could be related to the use of donor cells at various phases of the apoptotic process. However, it has been demonstrated that even in cells committed to the apoptotic pathway, this process can be reversed before the cells reach some critically advanced stage of apoptosis (Morita and Tilly, 1999); in other words, before cells pass the so-called “point of no return” in the apoptotic cascade. In this study, we proposed a new approach to investigate these two dichotomous themes by evaluating if donor fibroblasts at different phases of the apoptotic pathway could be reprogrammed by SCNT.
Because it is assumed that DNA integrity is a prerequisite for nuclear reprogramming of somatic cells by SCNT, we decided to use apoptotic cells early enough in the apoptotic process that DNA fragmentation could not be detected by TUNEL assay, which ended up being 3 h after STP treatment. Interestingly, our results showed that the use of Anx-positive donor cells for SCNT did not affect embryo production compared with parthenotes and nontreated controls. The apoptotic process of early Anx-positive fibroblasts seemed to be either reversed or interrupted after their transfer into enucleated bovine oocytes. Interestingly, besides being positive for annexin staining, donor cells showed a reduction in mitochondrial membrane potential and clear cytoplasmic and nuclear changes, including chromatin condensation, which are phenomena occurring during the cell death process.
These results are consistent with a study by Span et al. (2002), who showed that detection of early PS exposure, detected by annexin fluorescein isothiocyanate (FITC) assay, is not always associated with irreversible apoptosis, and that the proliferative capacity of Anx-positive CD34 cells was maintained after the apoptotic stimulus was removed. (Dumont et al., 2000) studied the reversibility of apoptosis by treating activated T lymphocytes with STP for 2 h followed by removal of the treatment, and reported that in the presence of a weak stimulus, rescue mechanisms present in the cell are able to save most of the cells in the apoptotic process, while in the presence of a stronger stimulus (or very long exposure), these rescue mechanisms can be overcome.
When compared to our previous study (Miranda Mdos et al., 2009) results obtained in this experiment seems to be conflicting at first glance, because we previously showed that Anx-positive cells had a negative effect on cloning and in the present study we showed that Anx-positive cells did not impaired embryo production. However, in fact, both results are complementary. In the first article, apoptosis was not induced by STP and “natural” Anx-positive cells were detected just after 24 h of serum starvation (15.7% of Anx-positive). In the present study cells were first serum starved, as we did previously, and then apoptosis was induced by STP (89.7% of Anx-positive cells at 3h of STP treatment).
We speculate that Anx-positive cells in first study were a very heterogeneous apoptotic population including cells in which apoptosis had been just started as well as cells which had bypassed the “nonreturn” point therefore explaining the 50% reduction in blastocyst rated, while in the present study STP induced Anx-positive cells represented a more homogeneus population in which most of cells had not bypassed the “nonreturn” point in the apoptotic process. Interestingly in the present study, when cells at more advanced stages of apoptosis (casp-9-positive) were sorted and used as nuclear donors, blastocyst production was impaired in ∼50%, similarly to results found for us previously.
Because the critical juncture defining the “point of no return” has not yet been clearly defined (Kroemer et al., 2009), our aim was to characterize this process by evaluating whether donor cells in more advanced stages of the apoptotic process could be reversed after introduction into oocyte cytoplasm. During the cell death process, the activation of caspase-9 is responsible for the activation of other effector caspases (e.g., caspase-3) that perform the major structural changes indicative of cell death, such as DNA fragmentation, etc.
We observed that donor cells treated with STP for 3 h containing activated caspase-9 decreased the production of cloned embryos approximately by half (12% blastocyst rate) in comparison to Control and Anx-positive groups. We speculate that different from the Anx-positive cell population, casp-9-positive cells are actually a heterogeneus cell population concerning the “apoptotic step” in which the cells are at the exact moment of the fusion (mixing of cytoplasms) to oocyte cytoplasm. As mentioned above, active caspase-9 is considered as the signaling caspase occuping a key site in apoptotic cascade. Therefore, we think that despite the fact that casp-9-positive cells share a common feature utilized for sorting, they are, in fact, a group of cells at different time point of the downstream apoptotic cascade, maybe at the limit of the “decision” about cell life or death in which only half of them were able to be reprogrammed by SCNT.
Considering the dramatic effect caused on blastocyst development we also speculated about the effect of casp-9-positive cells present within the Anx-positive group, which did not show blastocyst reduction. Taken together, the results of the time course experiment (Fig. 1) showing that around 25% of Anx-positive cells were casp-9-positive and the ∼50% reduction in blastocyst development observed after using casp-9-positive cells for SCNT, we would expect around 12.5% of reduction in blastocyst yelds in Anx-positive group (∼23.1% instead of 26.4%) caused by presence of casp-9-positive cells. Because the expected reduction did not change results at all, we conclude that SCNT itself did not influence the results.
Whether other caspases were already activated within the caspase-9-positive cell population sorted is unknown, but considering that the use of casp-9-positive cells as nuclear donors allowed blastocyst production after SCNT, it is tempting to speculate that until activation of caspase-9 some cells (maybe half of them) are on a reversible phase of the apoptotic pathway, with cell death becoming irreversible once at further steps are initiated. In future studies one could address this possibility by transferring caspase-3-positive cells into the oocyte's cytoplasm in order to determine what is the “cytoplasmatic limit” in the reversibility of apoptotic cells by SCNT.
Finally, these results confirm that adult fibroblasts at early stages of apoptosis may be reprogrammed by SCNT without affecting the developmental rate of cloned embryos, whereas donor cells with active caspase-9 negatively affected the production of cloned embryos. Because these caspase-9-positive cells had no indication of internucleosomal DNA fragmentation, we hypothesize that the “point of no return” for the apoptotic process is located between caspase-9 activation and DNA fragmentation.
Footnotes
Acknowledgments
The authors acknowledge Patrícia V.B. Palma for flow cytometry assistance and Ed Reyes for technical support and reagent purchasing. This work was supported by the Foundation for the Support of Research of the State of Sao Paulo (FAPESP), Brazil, and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPQ), Brazil.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.