
Abstract
Abstract
Porcine induced pluripotent stem cells (piPSCs) have the capacity to differentiate in vitro and in vivo and form chimeras. However, the lack of transgene silencing of exogenous DNA integrated into the genome and the inability of cells to proliferate in the absence of transgene expression are underlying reported problems, suggesting that reprogramming is not complete. The aim of the present study was to evaluate reprogramming events using a partially reprogrammed piPSC-like line expressing hOCT4, hNANOG, and hcMYC under tetracycline-regulated control to investigate the effects of these particular transgenes on the expression of the porcine endogenous pluripotency machinery. Endogenous and exogenous gene expression of OCT4, NANOG, SOX2, KLF4, and cMYC was determined at passages 5, 10, 15, and 20, both in cells cultured at 1 μg/mL doxycycline or 4 μg/mL doxycycline. Our results revealed that endogenous genes are repressed by their transgene counterparts in culture and that lack of expression of the transgenes, SOX2 and KLF4 allows for expression of endogenous SOX2 and KLF4. Furthermore, we report that alternate endogenous transcripts for pNANOG, pSOX2, and pKLF4 can also be detected in the pig. Despite the ability for some endogenous genes to be expressed in these lines, the piPSC-like cells still cannot be maintained without doxycycline, indicating that the culture system of piPSCs may not be optimal or that the reprogramming factor combination used may not currently be optimal for maintaining pluripotency in the pig. This may help to explain the difficulties in producing stable piPSCs and bona fide embryonic stem cell lines in this species.
Introduction
More recently, new technologies for producing piPSCs, which bypass transgene integration, have been developed. One of these approaches includes the use of episomal vectors (Telugu et al., 2010). Optimally, the episomal genes should be eliminated over a few passages, following the removal of antibiotics from the culture. In this particular case, however, genomic vector integration was evident in the piPSC clones, resulting in persistent expression, which was considered to be due to aberrations in methylation. Another nonviral and nonintegration approach for producing piPSCs was performed by Montserrat and colleagues using a polycistronic plasmid vector (Montserrat et al., 2011a). However, again, genomic integration of the plasmid was observed in the piPSC lines. A recent article has also shown that OCT4 may be indispensible for production of LIF-dependent piPSCs (Montserrat et al., 2011b). Given that the pig represents a good alternative mammalian model for investigating iPSC technology for potential future stem cell therapy, it is important that inserted transgenes are appropriately silenced and/or that nonintegrating plasmids actually remain in this condition and are subsequently eliminated. Investigating the mechanisms of reprogramming in this species and the ineffectiveness of transgene silencing will lead to improvements in the stability and reprogramming of piPSCs.
Tetracycline- or doxycycline-regulated expression of transgenes is one approach considered for silencing the transgenes effectively that has previously been applied in the pig (Wu et al., 2009). Interestingly, withdrawal of doxycycline resulted not only in expected loss of exogenous gene expression, but also loss of the capacity of the cells to proliferate and maintain pluripotency in vitro. In this case, differentiation occurred and the cells could not be maintained in culture, indicating incomplete, unstable reprogramming. Hence, the iPSC state could only be maintained in the presence of the transgene expression, driven by a high level of doxycycline (4 μg/mL). One consideration is that the level of transgene expression is relatively proportionate to the amount of doxycycline added to the culture (Blau and Rossi, 1999); therefore, it would be interesting to compare different levels of doxycycline and their impact on endogenous gene expression.
In our own research, we have produced partially reprogrammed piPSC-like lines using a Tet-On tetracycline-regulated expression system that expresses only some of the reprogramming transgenes (hOCT4, hNANOG, and hcMYC). We consider these lines to be a useful tool for evaluating reprogramming events related to production of piPSCs because they represent a partial state of reprogramming. It may be of further advantage to study these cell lines, without some of the transgenes present to determine if expression of endogenous pluripotency genes (e.g., pSOX2 and pKLF4) can be induced by manipulation of transgene expression with doxycycline. In this study, we assessed both the exogenous and endogenous expression of five pluripotency genes, OCT4, NANOG, SOX2, KLF4, and cMYC over long-term culture to determine the stability of exogenous versus endogenous expression over time. We also investigated the long-term endogenous expression of genes at low and high levels of doxycycline to determine the impact of this activator on the culture of piPSCs.
Materials and Methods
Production of piPSC-like line
Primary porcine embryonic fibroblasts were derived from day-24 Göttingen minipig embryos, in which the head, dorsal spine of the medial section, and tail were removed before collection of cell culture material. Adherent cells from the remains of the embryos were cultured in Dulbecco's modified Eagle's medium (DMEM; Sigma) containing 10% fetal calf serum (FCS; Invitrogen, Carlsbad, CA, USA) + penicillin/streptomycin (pen/strep; Sigma Aldrich, St. Louis, MO, USA). These cells were used as the background cell line for iPSC reprogramming. Embryonic porcine fibroblasts were transduced with a Tet-On inducible system, including a reverse tetracycline transactivator (rTA) driven by a ubiquitin promoter, together with lentivirus produced from five human reprogramming factors (OCT4, NANOGP8, SOX2, cMYC, and KLF4) (Addgene, Cambridge MA, USA) (Maherali et al., 2008) and 6 μg/mL polybrene/hexadimethrine bromide (Sigma Aldrich) (Fig. 1). Later, it was discovered the NANOG sequence used was a processed pseudogene, namely NANOGP8 as notified by Addgene.
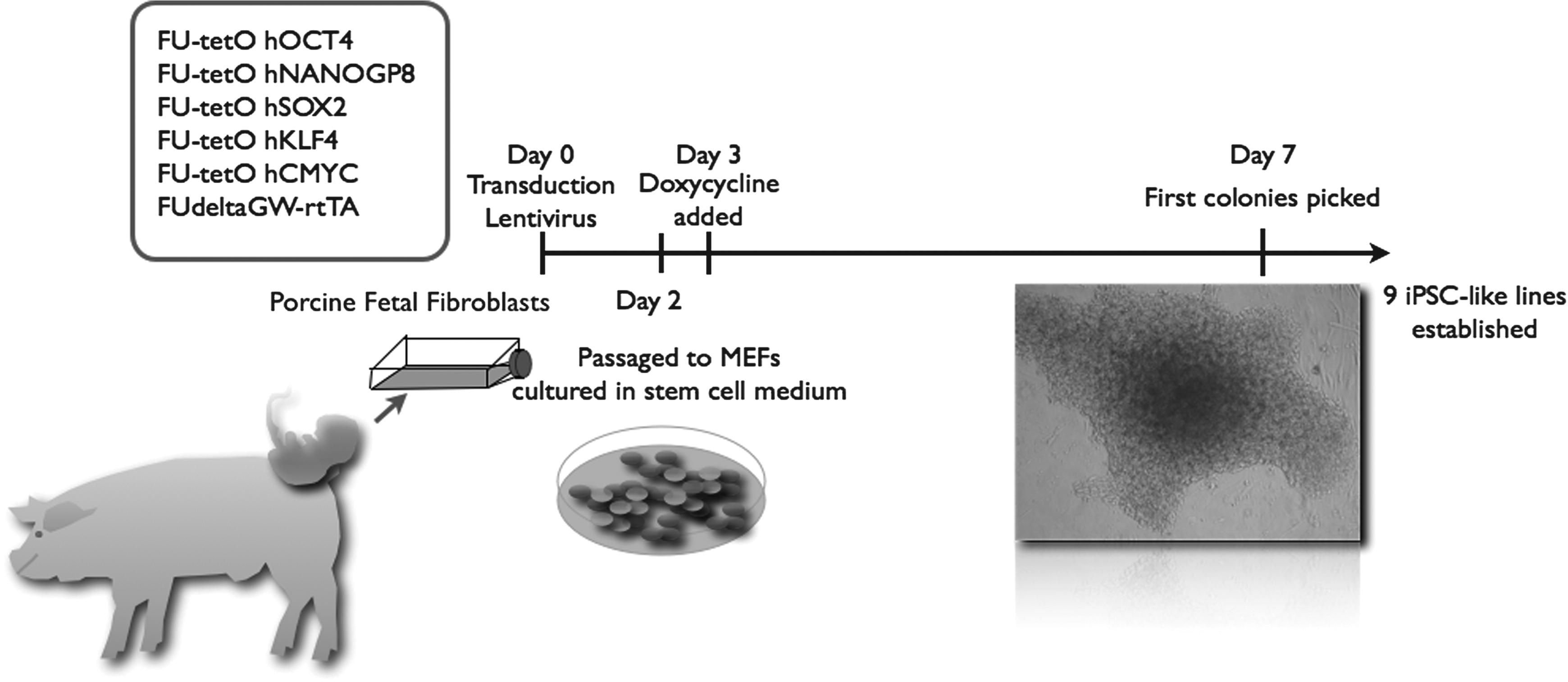
Schematic overview of reprogramming of porcine fetal fibroblasts using a Tet-On tetracycline-inducible expression system. MEFs, murine embryonic fibroblasts; IPSC, induced pluripotential stem cell.
A starting population of 0.5×106 cells was used for transduction and performed in triplicate. Transduced cells were passaged onto mitomycin C–treated (Sigma Aldrich) primary murine embryonic fibroblasts (MEFs) (background strain CF1) (Millipore, Billerica, MA, USA) on day 2 after transduction and cultured in knockout (KO)-DMEM (Invitrogen) containing 15% KnockOut™ Serum Replacement (Invitrogen), supplemented with 1× nonessential amino acids (Sigma Aldrich), 1×
Embryoid body assay
Cell colonies from piPSC3 were passaged mechanically (passage 8–12) into small clumps, which were cultured in 30 μL iPSC medium hanging droplets without bFGF, containing either 1 μg/mL or no doxycycline for 6 days at 5% CO2 in air. The hanging drop assay was repeated three times. Cells were harvested, and complementary DNA (cDNA) was prepared (as described below) and analyzed by reverse transcriptase PCR (RT-PCR) for expression of either markers of endoderm (AFP, FOXA2), mesoderm (HAND1, GATA2), ectoderm (NESTIN, NCAM, PAX6, TUJ1/BetaIII TUBULIN), or pluripotency (DNMT3B, GDF3, ZFP42/REX1) (Table 1).
Primer was also able to amplify a weak band in porcine tissue (indicative of cross-specificity).
PCR, Polymerase chain reaction; NCBI, National Center for Biotechnology Information; F, forward; R, reverse.
Transmission electron microscopy
Three individual colonies from piPSC3 were harvested at passage 8 for ultrastructural evaluation. Specimens were fixed in 3% glutaraldehyde in 0.1 M Na3PO4 buffer (pH 7.3) for 1 h at room temperature, then transferred and stored in 0.1 Na3PO4 buffer at 4°C. Samples were embedded in Bacto-agar (Difco Laboratories, Detroit, MI, USA), dehydrated, and embedded in Epon TAAB 812 embedding resin (VWR, West Chester, PA, USA), and processed for semithin and ultrathin sections as previously described (Hall et al., 2010; Hyttel and Madsen, 1987; Kallos, 2011). Semithin sections were observed using bright-field microscopy (Leica Microsystems, Bensheim, Germany), and ultrathin sections were observed under a transmission electron microscope (CM100, Philips, Darmstadt, The Netherlands).
Alkaline phosphatase assay
Detection of alkaline phosphatase in the piPSC lines piPSC3, piPSC7, and piPSC8 was performed using an Alkaline Phosphatase Detection kit (Millipore) according to the manufacturer's guidelines. piPSC3 was analyzed at passage 6 and piPSC7 and piPSC8 were analyzed at passage 7. Cell colonies were allowed to form for 5 days following passage. Fixation of cells was performed in 4% paraformaldehyde for 2 min.
Immunocytochemistry
For fluorescence immunocytochemistry, lines piPSC3, piPSC7, and piPSC8 were cultured on poly-L-lysine/laminin-treated (Sigma) glass cover slips (Hounisen, Aarhus, Denmark) and fixed in 4% paraformaldehyde (PFA) for 15 min at room temperature. Cells were stored in phosphate-buffered saline (PBS) at 4°C until analysis. They were permeabilized in 0.1% Triton-X 100 (Sigma Aldrich) for 1 h at room temperature, and antigen retrieval was performed by adding boiling citrate buffer (0.01 M) to the cells for 30 sec. Cells were washed two times with PBS and incubated in blocking buffer [5% normal donkey serum (Sigma Aldrich) in PBS] for 1 h at room temperature. They were cultured in diluted primary antibodies overnight at 4°C.
Primary antibodies were diluted in blocking buffer at the following dilutions: Goat polyclonal anti-OCT4, 1:250 (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA), rabbit polyclonal anti-NANOG, 1:500 (Peprotech, Rocky Hill, NJ, USA), mouse monoclonal anti-SOX2, 1:100 (R&D Systems, Minneapolis, MN, USA), goat polyclonal anti-KLF4, 1:400 (R&D Systems), mouse monoclonal anti-cMYC, 1:100 (Abcam, Cambridge, UK), mouse monoclonal SSEA1 1:100 (Biolegend, San Diego, CA, USA), rat monoclonal SSEA3 1:25 (Biolegend), mouse monoclonal SSEA4 1:100 (Biolegend), mouse monoclonal TRA-1-60 1:400 (Biolegend), and mouse monoclonal TRA-1-81 1:100 (Biolegend). It is important to mention that in the case of OCT4, NANOG, SOX2, KLF4, and cMYC, it was impossible to determine whether protein expression resulted from either porcine endogenous or human exogenous genes. Secondary antibody negative controls were performed by supplementation of primary antibody with blocking buffer. Cells were washed two times with PBS and then incubated in either fluorescein isothiocyanate (FITC) or Cy3 donkey-conjugated secondary antibodies raised in the appropriate species (i.e., mouse, rabbit, or goat) (Jackson Immunoresearch, Suffolk, UK), which were diluted at 1:200 in blocking buffer. Cells were washed two times in PBS and incubated in 0.1 μg/mL Hoechst 33342 (Sigma Aldrich). Cells were mounted on glass slides using fluorescent mounting medium (DAKO, Glostrup, Denmark) and observed by immunofluorescence imaging using a DMRB fluorescent microscope (Leica Microsystems). Images were captured using Leica Application suite (2.8.1) capture software (Leica Microsystems). A commercial human iPSC line (System Biosciences) was used as a positive control for SSEA3, SSEA4, TRA-1-61, and TRA-1-81. Porcine embryos were used as a positive control for SSEA1. A Danish ESC line LRB101 (kind gift from Claus Yding Andersen) and a mouse ESC line derived in Hungary by Biotalentum Ltd. were used as positive controls for OCT4, NANOG, SOX2, KLF4, and cMYC.
For chromogen immunocytochemistry (used for testing viral transduction efficiency), cells (including mouse ESCs and transduced/nontransduced porcine embryonic fibroblasts, p11) were fixed and permeabilized as described above. Endogenous peroxidase blocking was performed by incubating cells in 3% hydrogen peroxide (H2O2) in water for 10 min at room temperature. Blocking was performed in 10% normal animal (goat/rabbit) serum (Sigma Aldrich) in PBS for 30 min at room temperature. Cells were incubated in primary antibodies anti-SOX2, anti-KLF4, anti-OCT4, and anti-NANOG, diluted in blocking buffer, and incubated as described above. Biotinylated secondary antibodies were used at 1:250 (DAKO), and cells were incubated for 30 min at room temperature prior to incubation in strepavidin/horseradish peroxidase (HRP) (DAKO) at 1:500 for 30 min at room temperature. Cells were then stained with diaminobenzidine (DAB; DAKO) according to manufacturer's recommendations, and cell nuclei were counterstained with Mayer's hematoxylin for 30–40 sec. Images were taken using an inverted Leica DM IL microscope (Leica Microsystems), and a minimum of 500 cells were scored for assessment of transduction efficiency.
Production of genomic DNA, cDNA, and polymerase chain reaction
For assessment of transgene integration, genomic DNA was collected from cells from all nine lines isolated at passages 4–9. Cells were mechanically picked from MEF feeders and resuspended in PBS. DNA was isolated using the Qiagen Blood DNeasy kit (Qiagen, Hilden, Germany). For assessment of exogenous and endogenous gene expression, cells were mechanically harvested from piPSC3 cultured in either 1 μg/mL or 4 μg/mL doxycycline and harvested at passages 5, 10, 15, and 20. Cells were lysed in 50 μL of RLT buffer (Qiagen) containing β-mercaptoethanol (Sigma Aldrich) and snap frozen in liquid nitrogen; they were stored at −80°C until required for RNA extraction.
RNA was extracted using the RNAeasy Micro Kit (Qiagen) according to the manufacturer's instructions and included a DNA digest step with DNase I. Twenty nanograms of RNA was then amplified and converted into cDNA using the Qiagen QuantiTect® Whole Transcriptome Kit (QIagen) using the long amplification cycle according to the manufacturer's instructions. cDNA was also produced from a single isolated porcine epiblast (day 11 of development) and from human ESC line LRB101.
Polymerase chain reaction (PCR) was performed with diluted cDNA (100 ng/mL) using a 25 μL master mix composed of 5× reaction buffer (Fermentas/Thermo Scientific, Waltham, MA, USA), 2 mM deoxyribonucleotide triphosphates (dNTPs; Fermentas/Thermo Scientific), 1.5 mM MgCl2, (Fermentas/Thermo Scientific), 10 pmol/μL of both sense and antisense oligonucleotides (TAGC Oligos, Denmark), and 200 U/μL HotStart Taq polymerase (Fermentas) in RNase-free water. PCR conditions were as follows: Initial denaturation 95°C for 4 min followed by 35 cycles of 95°C for 1 min, 55°C for 1 min, 70°C 1 min. A further extension was performed at 70°C for 15 min. Porcine- and human-specific primers were designed and are listed in Table 1. OCT4 primers were designed to amplify the stem cell–specific isoform OCT4A (Wang and Dai, 2010). PCR products were mechanically isolated from agarose gels and cDNA purified using the Wizard® SV Gel and PCR Clean-up System (Promega, Madison, WI, USA) according to the manufacturer's instructions. The PCR products were sequence-confirmed by DNA Technology A/S (Risskov, Denmark).
Comparative RT-PCR
Comparative RT-PCR was performed on cDNA (produced as described above) obtained from line piPSC3 cells obtained at passages 5 and 20, both from low and high doxycycline cultures and from lines piPSC7 and piPSC8 cells obtained from passages 6 and 10, also cultured in both low and high doxycycline culture. Reactions were set up in triplicate using the Lightcycler 480 SYBR Green I Master Kit (F-Hoffman La Roche, Basal, Switzerland) in accordance with the manufacturer's instructions. Target genes were normalized to the Ct value of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and then normalized further to the Ct value of the positive calibrator tissue (for porcine genes, porcine epiblast cDNA; and for human genes, human ESC cDNA). The fold change in expression was determined using the calculation 2−ΔΔCt. Standard deviations were calculated from the 2−ΔΔCt value. Statistical analyses were also performed on the 2−ΔΔCt values, using an independent, unpaired, two-tailed students t-test and significance was determined when p≤0.05.
Results
Three cell lines display many typical characteristics of piPSCs
Stem cell-like colonies emerged as early as 7 days following transduction. Nine colonies were isolated from days 7 to 11 and cultured further and independently to form nine different cell lines. These lines were cryopreserved at early passages. Three lines, piPSC3, piPSC7, and piPSC8, were selected for further culture and in-depth analysis. In the case of piPSC3, the cells grew rapidly as compact, tight colonies with a domed-shape appearance, which appeared to be a number of layers thick; however, the borders of the colonies were irregular (Fig. 2A, B, D–F). No difference in morphology could be observed when this line was grown in either low (1 μg/mL) or high (4 μg/mL) doxycycline; however, cells were unable to proliferate in the absence of doxycycline (Fig. 2C). Line piPSC3 was able to grow beyond 20 cell passages without changes in colony morphology or proliferation rate (Fig. 2F). Lines piPSC7 and piPSC8 had a more flattened and less well-delineated appearance, and the cells contained lipid droplets

Morphology of porcine iPSC-like lines piPSC3, piPSC7, and piPSC8. Cell morphology of line piPSC3 revealed the formation of compact, domed-shaped colonies with irregular cell borders in low doxycycline (1 μg/mL) (
Immunocytochemistry was performed on all three lines for the pluripotent markers OCT4, NANOG, SOX2, and KLF4 and the surface markers, SSEA1, SSEA3, SSEA4, TRA-1-60, and TRA-1-80 from passages 5 to 7. cMYC was also analyzed in piPSC3. Results for expression in line piPSC3 are shown in Figure 3, and piPSC7 in Supplementary Figure 1 (Supplementary Data are available at www.liebertonline.com/cell). Results of expression in line piPSC8 were identical to that in piPSC7 and, therefore, data for piPSC8 are not shown. In the case of piPSC3, OCT4 and NANOG expression could be detected (Fig. 3A–F), whereas, mosaic SOX2 expression was initially observed, but became undetectable after passage 3 (Fig. 3G–I). A mosaic expression of both KLF4 and cMYC could be detected (Fig. 3J, L, M). This line was found to be negative for the cell-surface markers, SSEA1, SSEA3, SSEA4, TRA-1-60, and TRA-1-81 (Fig. 3K, L, O–R). RT-PCR revealed however, that this line expressed both DNMT3b and REX1 (detected at passage 10) (Suppl. Fig. 2C). In the case of piPSC7 and piPSC8, these cell lines expressed OCT4, NANOG, SOX2, and KLF4 until passage 6 (Suppl. Fig. 1A–I). Again, these lines were negative for SSEA3, TRA-1-60, and TRA-1-81 (Suppl. Fig. 1P–R); however, a few individual cells did stain positively for SSEA1 within colonies (Suppl. Fig. 1J–L) and some areas of the colonies expressed SSEA4 (Suppl. Fig. 1M–O). All three lines were found to be negative for alkaline phosphatase activity at passages 6 to 7 (data not shown).

Expression of pluripotent and cell-surface markers in line piPSC3. Line piPSC3 expresses OCT4 and NANOG (
Colonies from piPSC3 were harvested at passage 8 and analyzed for details of cell ultrastructure. The iPSC colonies were composed of tightly apposed cells of intercellular spaces observed between groups of cells (Fig. 4A). In the latter spaces, accumulation of apoptotic bodies and cell debris was noted (Fig. 4B). The individual iPSC cells were rounded and presented a spherical-to-ovoid nucleus, with abundant euchromatin and one or two prominent nucleoli, surrounded by sparse cytoplasm with mitochondria, sparse rough endoplasmic reticulum, and prominent polyribosomes (Fig. 4C). The presence of coated pits and vesicles indicated receptor-mediated endocytotic signaling. In some cells, the cytoplasmic compartment was larger and included prominent Golgi complexes and lipid droplets (Fig. 4D). Phagocytosis of apoptotic bodies and cellular debris, found between the cell chords, was not observed practically. Adjacent cells in the chords displayed closely apposed plasma membranes, but only very sporadic tight junctions were observed between them (Fig. 4E). Gap junctions were never observed. Occasionally, primary cilia were observed protruding into the intercellular space (Fig. 4F).

Light (
When subjected to embryoid formation assay, piPSC3 cells formed large embryoid body (EB)-like structures, but only in the presence of doxycycline. RT-PCR analysis of endoderm markers (AFP, FOXA2), mesoderm markers (HAND1, GATA2), and ectoderm markers (NESTIN, NCAM, PAX6, TUJ1/BIII TUBULIN) revealed that the expression of differentiation markers was less pronounced than in the undifferentiated piPSC-like cells (Suppl. Fig. 2). Interestingly, expression of ectoderm markers (NCAM, PAX6, TUJ1/BIIITUBULIN) could be observed in both undifferentiated P10 piPSC-like cells and in their EB counterparts. Moreover, piPSC-like cells also expressed other differentiated markers, including the endoderm marker FOXA2 and mesoderm marker GATA2, which indicates spontaneous differentiation may be more prevalent in standard culture conditions (Suppl. Fig. 2).
Induction of five human factors in porcine embryonic fibroblasts led to generation of porcine iPSC-like cells lacking hSOX2 and hKLF4 integration
When analyzed for incorporation of the exogenous human transgenes, it was determined that line piPSC3 was negative for hSOX2 and hKLF4 expression (Suppl. Fig. 3) (note that KLF4 bands visible in the figure are pKLF4). To determine whether silencing had occurred, or whether this was due to lack of genomic integration, we performed genomic PCR and confirmed that hSOX2 and hKLF4 had not integrated (Suppl. Fig. 4). Following this result, we performed further genomic PCR analyses on the eight other cell lines and found that in every case these lines also lacked hSOX2 and hKLF4 (data not shown) in spite of a high lentiviral transduction activity of hSOX2 and hKLF4 (SOX2 88%, KLF4 68% transduction efficiency), which was comparable to the transduction activity of OCT4 (81%) and NANOG (82%) (Suppl. Fig. 4).
Endogenous porcine genes are repressed in iPSC-like cells when the equivalent transgene is present, whereas when the transgene is lacking, the equivalent endogenous gene is not suppressed
We further analyzed lines piPSC3, piPSC7, and piPSC8 to gain additional insight into the mechanisms and effects of particular transgenes on the reprogramming of their endogenous counterparts using RT-PCR and comparative RT-PCR in line piPSC3 (Fig. 5 and Suppl. Fig. 3) and comparative RT-PCR in lines piPSC7 and piPSC8 (Fig. 5). In particular, we addressed whether endogenous expression of genes that lack a transgene counterpart (i.e., pSOX2 and pKLF4) could be induced. We also investigated whether endogenous gene expression over time in culture could change.

Comparative real-time PCR of exogenous and endogenous OCT4, NANOG, SOX2, KLF4, and cMYC was performed in three porcine iPSC-like lines, piPSC3, piPSC7, and piPSC8. piPSCs grown in either low or high levels of doxycycline. Graphs depict relative expression of genes in piPSC3 at p5 and p20 and relative expression of genes between piPSC7 and piPSC8 at p6 and p10. Relative expression is shown as the fold change (calculated using 2−ΔΔCt). Y error bars depict standard deviations of the 2−ΔΔCt value. Statistical differences between low and high doxycycline are depicted for comparisons within the same passage and within the same cell line. Statistical analysis was performed using a Student independent, unpaired, two-tailed t-test. Statistical significance is shown when p≤0.05.
Porcine- and human-specific primers were designed to amplify either the endogenous or exogenous genes, respectively. These primers were tested in pooled porcine epiblast and human ESCs. All primers were found to be species specific with the exception of the human SOX2 and KLF4 primers, which were found to amplify endogenous genes in the porcine positive control tissue (Suppl. Fig. 3A). However, given that all of the lines lacked human SOX2 and KLF4, we assumed any expression would be indicative of porcine endogenous expression. When these primers were actually tested in the piPSC-like line, endogenous pKLF was amplified using the human primers, but pSOX2 was not amplified by the hSOX2 primers (Suppl. Fig. 3A).
RT-PCR analyses revealed that hSOX2 and hKLF4 were lacking from passage 5 to passage 20 in the piPSC3 line (as expected from our genomic PCR analyses); however pSOX2 and pKLF4 could be detected throughout the culture (Suppl. Fig. 3). Considering that the human SOX2 and KLF4 transgenes were missing, we attribute the protein expression of SOX2 and KLF4 in our piPSCs to be endogenous (Fig. 3). Further analysis by comparative RT-PCR confirmed this downregulation of hSOX2 and hKLF4 and the upregulation of the equivalent endogenous genes (pSOX2 and pKLF4) occurred (Fig. 5). To be certain this was not an effect in an individual line, we performed comparative RT-PCR on two other lines, piPSC7 and piPSC8, which also revealed an upregulation of pSOX2 and pKLF4 in the absence of the human transgenes SOX2 and KLF4 (Fig. 5). In the case of OCT4, NANOG, and cMYC, we found the opposite to be the case in all three lines, where the endogenous porcine genes were downregulated compared to their equivalent human transgenes (Fig. 5 and Suppl. Fig. 3). There were only two observed exceptions to this during analysis of the varying different passages and low and high doxycycline conditions between all three lines. This was when endogenous pNANOG was found to be more upregulated than hNANOG. This was the case at passage 20 in piPSC3 when cells were cultured in high doxycycline, and at passage 10 in piPSC7 when cells were cultured in low doxycycline.
Expression of endogenous genes reveals presence of spliced variants or alternate transcripts for pSOX2, pKLF4, and pNANOG
Expression of endogenous porcine genes by RT-PCR in piPSC3 cells revealed the presence of spliced variants or alternate transcripts (Suppl. Fig. 3). Interestingly, under low doxycycline, endogenous pSOX2 and an alternate transcript of pKLF4 could be detected at passage 5. Endogenous expression of pSOX2 and pKLF4, however, varied during culture. By passage 10, a splice variant of pSOX2, as well as two bands for pKLF4, were detected. The pSOX2 band and larger pKLF4 bands were sequenced and confirmed to be alternate transcripts.
To gain further understanding of the two pKLF4 transcripts detected, an alignment was performed between pig (NM_001031782.1), mouse (NM_010637.3), and human KLF4 (NM_004235.4) mRNA [sequences obtained from National Center for Biotechnology Information (NCBI)]. The pig mRNA contains an additional exon (which was amplified in our primer set), which is not found in either mouse or human KLF4 (Fig. 6). This exon is found directly between the mouse and human exons 3 and 4. DNA sequencing confirmed that the two transcripts obtained for pKLF4 contain a variant, including the additional exon and a variant excluding this exon, indicative of the presence of two isoforms.

An alignment of mouse (NM_010637.3), human (NM_004235.4), and pig (NM_001031782.1) sequences from GenBank NCBI of KLF4 indicate that an extra exon exists in the porcine that lies between the mouse and human exons 3 and 4.
A further interesting note should be made regarding endogenous expression of pNANOG. From the RT-PCR analyses on piPSC3, the correct expected-size band of pNANOG could only be detected in low doxycycline conditions at passage 10 and at passages 10 and 15 in high doxycycline. A larger PCR product of 300 bp was observed at passages 5 and 20 in low doxycycline and at passage 20 in high doxycycline. Considering that we obtained a correct-size band at other passages, we believe that this could be a potential pseudogene, spliced-variant, or alternate transcript; however, further analyses are needed to clarify this.
The level of doxycycline used does not alter the general expression levels of either the transgenes or endogenous genes
We considered it would be worthwhile to investigate how the level of doxycycline may impede the endogenous gene expression of OCT4, NANOG, SOX2, KLF4, and cMYC. Therefore, lines piPSC3, piPSC7, and piPSC8 were cultured in either 1 μg/mL (low) or 4 μg/mL (high) doxycycline culture over time. A temporal fluctuation in both transgene and endogenous gene expression was observed in all three lines. Although significant differences could be determined when comparing low and high doxycycline at the same cell passage within the same cell line, no consistent trend was observed (Fig. 5); i.e., low doxycycline conditions did not result in consistently lower expression of the exogenous genes. Similarly, low doxycycline did not result in consistently higher expression of the porcine endogenous genes. Therefore, we conclude that the level of doxycycline does not significantly impact on either the level of transgenes or endogenous genes and that temporal changes in the levels of both transgenes and endogenous genes over time in culture may be due to other factors.
Discussion
Significant problems remain in the generation of bona fide porcine iPSCs; they include the lack of transgene silencing of plasmid DNA integrated into the genome and an inability of cells to proliferate in the absence of transgene expression. These issues suggest that reprogramming of these cells is not complete. In addition, bona fide ESCs have not been generated from this species. Reasons for the difficulties have been postulated to be due to differences in preimplantation development, differences in expression of pluripotent markers, as well as inadequate culture conditions (Hall, 2008; Hall et al., 2010). Despite clear differences in expression of OCT4 and NANOG during development (Gao et al., 2011; Hall et al., 2010; Wolf et al., 2011), a pluripotent state can be induced in porcine cells overexpressing mouse or human reprogramming factors, including OCT4 and NANOG.
Two possible scenarios may be inferred. The human or mouse genes are functionally different from their porcine counterparts, or there is no difference in gene functionality, but that in the overexpressed state, these cells may override inadequacies in the culture medium to sustain pluripotency. The latter, or a combination of both, may likely be correct, considering piPSCs cannot be maintained in vitro in the absence of transgene expression (Wu et al., 2009). This also suggests that culture conditions have not yet been optimized to enhance reprogramming and create stable piPSC lines relying on the endogenous pluripotency machinery. The effect of culture on reprogrammed cells is significant. This has recently been eloquently shown, when neural progenitor cells were formed, by reprogramming mouse fibroblast cells with the reprogramming factors Oct4, Sox2, Klf4, and cMyc; but in this case, the ESC medium was swapped to a neural medium (Kim et al., 2011). Thus, it is likely that the reprogramming factors may prime cells to an open state that can then be altered by the culture conditions used (Kim et al., 2011).
In this study, tetracycline-regulated piPSC-like cells (with genomic hOCT4, hNANOG, and hcMYC integration) were produced using Göttingen minipig embryonic fibroblasts. The piPSC3 line was characterized by transmission electron microscopy and was shown to share several major ultrastructural characteristics with the inner cell mass cells of the day 6 porcine blastocyst (Hall et al., 2010). Both cell types have a large nucleus, characterized by abundant euchromatin and prominent nucleoli, and a relatively sparse cytoplasm. Most of the iPSC-like cells displayed a very sparse cytoplasm presenting few organelles, features that are also characteristic for the pluripotent cell population in human embryonic stem cell colonies (Jacobsen et al., 2011). Some of the individual iPSC-like cells were more rich in cytoplasm and had a higher abundance of Golgi complexes and lipid droplets. We hypothesize that the degree of reprogramming within an iPSC population is diverse, and that the cytoplasm-rich cells may represent less well reprogrammed, more somatically imprinted cells in the population. Immunocytochemistry revealed that pluripotent genes could be detected, including OCT4, NANOG, SOX2, KLF4, and cMYC, but did not express any of the stage-specific embryonic antigen (SSEA) or TRA cell-surface markers analyzed. This line could produce EBs but was unable to differentiate well in vitro. Two other piPSC lines with different morphology (i.e., flattened) were also characterized by immunocytochemistry. These two lines expressed OCT4, NANOG, SOX2, and KLF4 and were also found to express very low levels of SSEA1 and SSEA4 in some areas of some colonies. These lines did not express alkaline phosphatase. Endogenous SOX2 and KLF4 could be detected in all three lines (which was confirmed due to the absence of the human SOX2 and KLF4 transgenes), although SOX2 was lost at early passages in piPSC3. These cell lines, despite their lack of true pluripotency, were found to be useful for evaluating how transgenes may affect endogenous gene expression in this species.
Despite the high activity of hSOX2- and hKLF4-carrying lentiviruses, transgene integration was not observed in nine different clones. Reasons for this lack of integration remain unclear. It could be postulated that the cell background may have an influence on the reprogramming ability. However, this cannot necessarily be the case, because other researchers have been able to establish porcine iPSC lines from D37-derived fetal fibroblasts from the Tibetan minipig (Esteban et al., 2009). Future evaluation of other cell lines may be the only way to ascertain if this is the case. In addition, it could be worth in future experiments to retransduce these cells with hSOX2 and hKLF4 to determine if integration may occur. Despite the fact that the evaluated cell lines only expressed the transgenes OCT4, NANOG, and cMYC, endogenous expression of both pSOX2 and pKLF4 could be detected through to passage 20 and expression of endogenous SOX2 and KLF4 could be detected in early culture. In addition, it was later discovered that the NANOG plasmid obtained from Addgene and produced by Konrad Hochedlinger's group (Maherali et al., 2008), was in fact a pseudogene, NANOGP8. Evidence suggests, this pseudogene is highly similar to the NANOG gene, is human specific, and processed (Fairbanks and Maughan, 2006). In addition, this pseudogene is translated into protein (considered a retrogene), affects cell proliferation, and plays a role in tumorigenesis (Zhang et al., 2006).
Despite this lack of transgene integration, the piPSC3 line was found to express REX1 and DNMT3B and cells had an epithelial-like morphology. Cells could be grown for at least 20 cell passages and could also form EB-like structures in vitro. Furthermore, two additional piPSC-like lines, piPSC7 and piPSC8, also expressed endogenous SOX2 and KLF4, and this was also confirmed at the RNA level by comparative RT-PCR up until passage 10. Furthermore, in all three piPSC-like lines, we show clearly that when the transgene is expressed (i.e., OCT4, NANOG, and cMYC), the counterpart endogenous gene is repressed. This may be the first evidence to reveal that in the pig, endogenous gene expression may only be possible when the reprogramming transgene is absent.
These lines may be useful for further investigation of the regulatory mechanisms of reprogramming in this species. Reasons for the lack of integration of hSOX2 and hKLF4 remain a mystery; however, the future use of polycistronic vectors containing a minimum of four transgenes (e.g., OKSM; Carey et al., 2009; Gonzalez et al., 2009; Sommer et al., 2009), which have been applied to mouse and human cells, may help to override future issues regarding lack of integration of individual factors in porcine cells. A major issue that still needs to be overcome in this species is how to silence all the transgenes, induce the expression of all the endogenous genes, and maintain proliferation and pluripotency of piPSCs in culture.
Another insight from this study related to the expression of alternate endogenous transcripts of pSOX2, pKLF4, and pNANOG, which has not been described previously. An alternate pSOX2 transcript was observed that varied temporally during culture within the same cell line. In addition, two KLF4 alternated transcripts were detected. Coincidently, the KLF4 primers, which were designed to span the third intron, were able to delineate between two different-sized KLF4 PCR products. These two KLF4 transcripts are identical, with the exception of one containing an additional exon. This exon is lacking in both mouse and human KLF4 gene sequences. Functional assays looking at the roles of these two transcripts would be intriguing. The transcripts are not likely to be alternatively spliced variants, because alternative splicing has not been detected for KLF4 in the mouse; rather, six different pseudogenes have been described (Godmann et al., 2005). This is the first case in which an alternate KLF4 transcript has been described in porcine stem cell–like cells. An alternate transcript was also detected for pNANOG. Further analysis of these transcripts may reveal insight into the roles they play in regulating gene function and their possible role in stem cell regulation.
A previous study reportedly cultured piPSCs using high levels of doxycycline to regulate iPSC transgene expression (Wu, et al., 2009). Therefore, we decided to investigate if doxycycline levels could affect endogenous gene expression. In the present study, the expression of endogenous genes was dynamic during culture, and differences could be observed when altering the level of doxycycline. However, these differences were inconsistent with the level of doxycycline used and differed between cell lines, suggesting that doxycycline may not be an important consideration in inducing endogenous gene expression in piPSCs.
Our piPSC lines could not be sustained in the absence of doxycycline, as previously reported by other researchers. This outcome may be due to insufficient culture conditions, which do not allow for activation of endogenous genes and maintenance of cell signaling events and which are important for sustaining the culture of porcine pluripotent cells in vitro. This may also explain why bona fide porcine ESCs have not been produced to date. Another consideration may relate to differences in cell signaling in porcine pluripotent cells compared to the mouse and human. Recently, we have reported that the porcine epiblast expresses low levels of NANOG and cMYC (Gao et al., 2011). It may be that these genes do not play a significant role in pluripotency in this species, and the discovery of alternate genes, which regulate pluripotency and stem cell renewal, may potentially improve future reprogramming of piPSCs.
To conclude, we have been able to demonstrate important aspects of reprogramming porcine cells using a tetracycline-regulated expression system. In this study, we have shown that the embryonic fibroblasts used in this study could not be reprogrammed into bona fide piPSCs. In three piPSC-like cell lines, we identified the integration of hOCT4, hNANOG, and hcMYC, and showed that the expressed transgenes repress their endogenous counterparts. In the absence of hSOX2 and hKLF4, we could see the upregulation of their endogenous counterpart genes, suggesting that the transgenes themselves directly repress the endogenous counterparts. These cell lines, however, could not be sustained without transgene expression. Hence, appropriate culture conditions for sustaining piPSCs on the basis of their endogenous pluripotency machinery need to be developed and further cell signaling experiments need to be performed to reveal whether other pluripotent genes are needed for maintenance of pluripotency in this species.
Footnotes
Acknowledgments
We would like to thank Professor Claus Yding Andersen for donation of the human ESC line LRB101 for these experiments. We would also like to thank the following for financial support: European Union (EU) projects EU FP7 PartnErS, PIAP-GA-2008-218205, InduVir, PEOPLE-IRG-2009-245808, AniStem, PIAP-GA-2011-286264 and PluriSys, HEALTH-2007-B-223485, the Danish National Advanced Technology Foundation, Pigs and Health Project, and the Danish Council for Independent Research, Technology and Production Science in Denmark. We also thank Ellegaard, Denmark for supply of Göttingen minipig tissues for this project.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.