
Abstract
Abstract
In somatic cell nuclear transfer (SCNT) procedures, exquisite enucleation of the recipient oocyte is critical to cloning efficiency. The purpose of this study was to compare the effects of two enucleation systems, Hoechst staining and UV irradiation (hereafter, irradiation group) and Oosight imaging (hereafter, Oosight group), on the in vitro production of bovine SCNT embryos. In the Oosight group, the apoptotic index (2.8±0.5 vs. 7.3±1.2) was lower, and the fusion rate (75.6% vs. 62.9%), cleavage rate (78.0% vs. 63.7%), blastocyst rate (40.2% vs. 29.2%), and total cell number (128.3±4.8 vs. 112.2±7.6) were higher than those in the irradiation group (all p<0.05). The overall efficiency after SCNT was twice as high in the Oosight group as that in the irradiation group (p<0.05). The relative mRNA expression levels of Oct4, Nanog, Interferon-tau, and Dnmt3A were higher and those of Caspase-3 and Hsp70 were lower in the Oosight group compared with the irradiation group (p<0.05). This is the first report to show the positive effect of the Oosight imaging system on molecular gene expression in the SCNT embryo. The Oosight imaging system may become the preferred choice for enucleation because it is less detrimental to the developmental potential of bovine SCNT embryos.
Introduction
To improve the enucleation procedure and cloning efficiency, several methods have been developed, including blind, Hoechst staining and ultraviolet (UV) irradiation, spindle imaging, and centrifugation (Liu et al., 2000; McGrath and Solter, 1983; Smith and Wilmut, 1990; Tatham et al., 1995). The Hoechst staining and UV irradiation approach is used routinely, particularly in domestic species, to ensure error-free enucleation by enabling clear visualization of the chromosomes before removal of the dense cytoplasm from the oocytes prior to nuclear transfer. This procedure involves the removal of a small amount of cytoplasm surrounding the spindle. However, even very brief UV exposure can have negative effects on the membrane integrity, mitochondrial DNA, and further embryo developmental potential (Smith, 1993).
As a more stable enucleation technique, spindle imaging was introduced for the direct enucleation of oocytes using polarized light microscopy (Liu et al., 2000). In human in vitro fertilization programs, spindle imaging (in this study, denoted as the Oosight imaging system) is mainly used to locate the meiotic spindle to avoid disrupting it while injecting oocytes (Kilani et al., 2011). This noninvasive and reliable technique has been used to improve cloning efficiency in pigs (Li et al., 2010). However, there has been no report of a positive effect of the Oosight imaging system on gene expression in SCNT embryos.
In this study, we examined the effect of two enucleation systems, Hoechst staining and UV irradiation (hereafter, irradiation group) and Oosight imaging (hereafter, Oosight group), on the in vitro production of Jeju Black Cattle (JBC; Korean Native Cattle) SCNT embryos and their developmental potential. We analyzed differences in cloning efficiency with different enucleation systems on the basis of apoptosis and specific gene expression by terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay and semiquantitative real-time PCR.
Materials and Methods
Unless stated otherwise, all chemicals were purchased from Sigma Chemical Company (St Louis, MO, USA).
Preparation of donor cells
Donor somatic cells were derived from JBC ear tissue. Sliced ear tissues were incubated in 0.1% collagenase type IV solution at 38°C for 1.5 h and cultured in donor cell culture medium [Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS; Gibco, Grand Island, NY, USA), 1 mM sodium pyruvate, 1% nonessential amino acids, 0.1% β-mercaptoethanol, and 1% penicillin-streptomycin]. The cells were grown and subcultured three to five times at intervals of 4–6 days and frozen with 1×106 cells in cryovials (1.5 mL) in a freezing medium [50% donor cell culture medium plus 45% FBS and 5% dimethyl sulfoxide (DMSO)].
For SCNT, frozen–thawed ear cells were washed twice with donor cell culture medium and treated with 3 mg/mL protease for 50 sec at room temperature. Treated cells were washed three times and resuspended in a donor cell preparation medium [tissue culture medium 199 (TCM-199)–HEPES (Gibco) supplemented with 0.2 mM sodium pyruvate].
Preparation of recipient oocytes
Bovine ovaries were collected from a slaughterhouse and transported to the laboratory within 2 h in 0.9% saline at 35°C. Cumulus oocyte complexes (COCs) were aspirated from visible follicles (2–6 mm in size) with an 18-gauge needle attached to a 10-mL disposable syringe. The medium used for COCs collection was HEPES-buffered Tyrode's medium (TL-HEPES). Sets of ten COCs were in vitro matured in TCM-199 (Gibco) supplemented with 10% FBS, 0.2 mM sodium pyruvate, 1 μg/mL follicle-stimulating hormone (Folltropin™, Bioniche Animal Health, Belleville, ON, Canada), 1 μg/mL estradiol-17β, and 1 mM epidermal growth factor (EGF) in mineral oil at 38.8°C in an incubator (5% CO2, 5% O2, and 90% N2) for 18–20 h.
For enucleation in the irradiation group, denuded oocytes were labeled with 5 μg/mL Hoechst 33342 for 10 min, washed three times, and transferred into enucleation medium (TCM-199–HEPES containing 20% FBS and 7.5 μg/mL cytochalasin B). The metaphase (M) II plate and first polar body (PB1) were visualized by exposure to UV irradiation for 10 sec and removed by squeezing (Fig. 1A). In the Oosight group, the MII plate and PB1 were visualized with an inverted microscope (Olympus, Tokyo, Japan) equipped with a Oosight spindle-check system (CRi, Hopkinton, MA, USA) and removed by the same squeezing method (Fig. 1A).
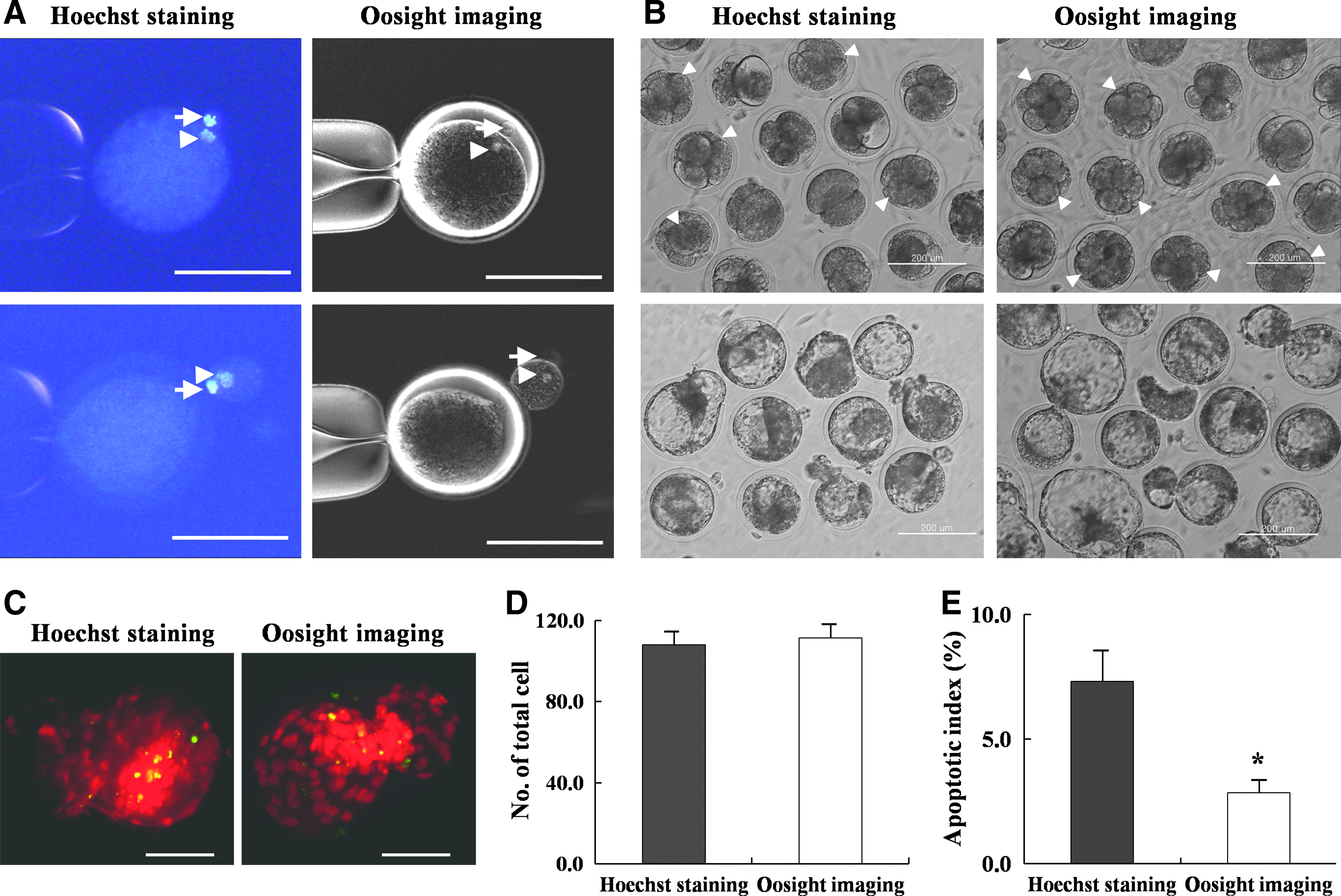
JBC-SCNT embryos produced in vitro with different enucleation systems and their TUNEL-stained fluorescence microscopic images. (
SCNT
A single treated donor cell was placed in the perivitelline space of the enucleated oocytes in nuclear transfer medium [TCM-199—HEPES containing 0.06% fatty acid-free (FAF)-bovine serum albumin (BSA), and 10 μg/mL phytohemagglutinin] through the opening made during enucleation. Oocyte–cell couplets were placed in cell fusion medium (0.3 M mannitol, 0.5 mM HEPES, 0.05 mM CaCl2, and 0.1 mM MgSO4) and subjected to an electrical pulse of 1.3 kV/cm for 20 μsec with an Electro Cell Fusion Generator (LF101, NEPAGENE, Chiba, Japan). After fusion, the reconstructed embryos were kept in TCM-199–HEPES supplemented with 20% FBS for 1 h, activated in 10 μM calcium ionophore for 5 min, and exposed to 2 mM 6-dimethylaminopurine for 3 h.
Culture of JBC-SCNT embryos
After activation, the reconstructed embryos were cultured in CR1aa medium supplemented with 0.03% FAF-BSA for 2 days. They then were co-cultured on the same JBC ear feeder cell drop in CR1aa medium containing 10% FBS, 1 μM EGF, and 1 μM insulin-like growth factor (IGF) at 38.8°C in an incubator (5% CO2, 5% O2, and 90% N2) for 6 days.
Blastocyst differential staining
The blastomere, inner cell mass (ICM), and trophectoderm (TE) cell numbers in blastocysts were counted by differential staining, according to the method of Thouas et al. (2001). Zona-intact blastocysts were incubated in 500 μL of Solution 1 [TL-HEPES containing 1% Triton X-100 and 100 μg/mL propidium iodide (PI)] for 30 sec. Blastocysts were then immediately transferred into 500 μL of Solution 2 (100% ethanol with 25 μg/mL bisbenzimide; Hoechst 33258) and stored at 4°C overnight. Blastocysts were mounted onto a slide glass and observed by fluorescence microscopy equipped with an ultraviolet (UV) filter. The PI- and bisbenzimide-labeled TE nuclei appeared pink or red. Bisbenzimide-labeled ICM nuclei appeared blue.
TUNEL assay
The numbers of apoptotic cells of day-8 JBC-SCNT blastocysts produced in the two different enucleation systems were determined with the In Situ Cell Death Detection Kit (Roche, Mannheim, Germany). Briefly, SCNT blastocysts were fixed in 3.7% paraformaldehyde in phosphate-buffered saline (PBS) for 1 h, permeabilized in 0.3% Triton X-100 for 1 h, and incubated with fluorescein-conjugated deoxyuridine triphosphates (dUTPs) and terminal deoxynucleotidyl transferase enzyme in the dark for 1 h. This process was followed by incubation in 50 μg/mL RNase A for 1 h at 37°C, while the nuclei were simultaneously counterstained with 40 μg/mL PI. Stained blastocysts were loaded onto glass slides and observed by fluorescence microscopy equipped with a UV filter. Red, green, and yellow (merged) indicate chromatin, fragmented DNA, and fragmented DNA of an apoptotic blastomere, respectively. The apoptotic index was determined as the percentage of yellow blastomeres among the total number of red blastomeres.
Real-time RT-PCR quantification
Comparative real-time PCR was performed using a Chromo 4 (Bio-Rad) and DyNAmo HS SYBR Green qPCR kit (Finnzymes), according to the manufacturer's instructions, with the primers described in Table 1. Messenger RNA (mRNA) for real-time reverse transcriptase (RT)-PCR was prepared from 15 JBC-SCNT blastocysts from each of the two enucleation systems by using magnetic beads (Dynabeads mRNA purification kit; Dynal, Oslo, Norway). Gene expression was quantified by the 2−ΔΔCt method (Livak and Schmittgen, 2001).
Experimental design
To examine the effect of the two different enucleation systems on JBC-SCNT embryo production, oocytes enucleated by either Hoechst staining and UV irradiation or by the Oosight imaging system were treated under the same SCNT conditions with the same donor cells. The fusion rate, cleavage rate at day 2, blastocyst rate at day 8, total cell and ICM cell number in blastocysts, and overall efficiency were compared between the two groups. The apoptotic index for in vitro–produced 8-day-old JBC-SCNT blastocysts and the relative mRNA expression levels for seven candidate genes (Oct4, Sox2, Nanog, Interferon-tau, Caspase-3, Dnmt3A, and HSP70) were determined in both groups. Experiments were repeated six times.
Statistical analysis
Differences in the fusion rate, developmental rate, total cell and ICM cell number in blastocysts, overall efficiency, apoptotic index, and gene expression levels between the irradiation group and the Oosight group were examined. Differences were evaluated by analyses of variance (ANOVA) with the general linear model (PROC-GLM) in the SAS software program. Differences of p<0.05 were considered significant.
Results
Effect of the two different enucleation systems on in vitro development of JBC-SCNT embryos
When SCNT was used with two different enucleation systems, as shown in Figure 1A, the enucleation volumes between the two groups were not different. After enucleation by Hoechst staining and UV irradiation or the Oosight imaging system, the cell fusion rates between JBC donor cells and enucleated recipient oocytes were 62.9% (204/324) and 75.6% (242/320), respectively (p<0.01). At day 2 postactivation, the cleavage rates of embryos reconstructed from the fused oocytes were 63.7% (130/204) and 78.0% (189/242) for the irradiation and Oosight group, respectively (p<0.05). When the day-2 JBC-SCNT embryo quality was compared, the good-quality four- to eight-cell (≥four cell) JBC-SCNT embryo development rates were higher in the Oosight group (71.4%, 135/189) than in the irradiation group (50.8%, 66/130), whereas the two- to three-cell (<four cell) JBC-SCNT embryo development rates were higher in the irradiation group (49.2%, 64/130) than in the Oosight group (28.6%, 54/189) (each p<0.01) (Table 2 and Fig. 1B).
Means with different superscripts in the same column are significantly different (a,b p<0.01; c.d p<0.05).
JBC-SCNT, Jeju Black Cattle– somatic cell nuclear transfer; ICM, inner cell mass.
At day 8 postactivation, the blastocyst development rates of JBC-SCNT embryos were higher in the Oosight group (40.2%, 76/189) than in the irradiation group (29.2%, 38/130) (p<0.01). When the expanding numbers of 8-day-old JBC-SCNT blastocysts were compared, the hatching blastocyst rate was higher in the Oosight group (48.6%, 37/76) than in the irradiation group (39.5%, 15/38), as shown in Figure 1B (p<0.05). The average total cell number in the Oosight group (128.3±4.8) was higher than that in the irradiation group (112.2±7.6) (p<0.05), but the ICM cell numbers were not different between the two groups. Finally, the overall efficiency in producing JBC-SCNT embryos was higher in the Oosight group (23.7%, 76/320) than in the irradiation group (11.7%, 38/324). Therefore, cloning efficiency was significantly different between the two groups (p<0.05).
Effect of two different enucleation systems on cell death in JBC-SCNT embryos
Approximate values for the apoptotic index per blastocyst were determined by fluorescence microscopy with the TUNEL assay (Fig. 1C–E). The apoptotic index of the Oosight group (2.8±0.5) was lower than that of the irradiation group (7.3±1.2) (p<0.05).
Relative mRNA expression of candidate genes in JBC-SCNT embryos created by different enucleation systems
The relative mRNA expression levels of genes related to pluripotency (Oct4, Sox, Nanog), maternal recognition of pregnancy (Interferon-tau), apoptosis (Caspase-3), de novo methylation (Dnmt3A), and stress (Hsp70) were analyzed. As shown in Figure 2 and Table 3, the mRNA expression level for the core pluripotency marker gene Oct4 was significantly higher in the Oosight group (1.49-fold) compared with that of the irradiation group. However, the expression levels of Sox2 and Nanog mRNA were not different between the two groups (p<0.05). The relative abundance of Interferon-tau gene expression was significantly higher (1.88-fold) in the Oosight group than in the irradiation group. Conversely, the mRNA expression levels of Caspase-3 and Hsp70 mRNA were lower (0.22- and 0.27-fold, respectively) in the Oosight group than in the irradiation group (p<0.05). The mRNA expression level for the de novo methylation and methylation maintenance marker gene Dnmt3A was slightly higher (1.20-fold) in the Oosight group than in the irradiation group.

Relative mRNA expression of candidate genes of 8-day-old JBC-SCNT embryos produced by different enucleation systems. Expression levels of genes related to pluripotency (Oct4, Sox2, and Nanog), implantation (Interferon-tau), proapoptotic activity (Caspase-3), demethylation (Dnmt3A), and stress (Hsp70) were examined. Bars with different superscripts within a panel differ significantly (p*<0.05). Error bars indicate standard deviation.
JBC-SCNT, Jeju Black Cattle– somatic cell nuclear transfer.
Discussion
This study demonstrates that the direct enucleation of oocytes using the Oosight imaging system can significantly improve the in vitro developmental potential of bovine SCNT embryos compared with the more conservative approach of enucleation using Hoechst staining and UV irradiation. In this study, SCNT embryos were reconstructed using JBC (a species of Korean Native Cattle) ear cells as donor cells. Significant differences between the two enucleation methods were observed in the cell fusion rate, cleavage rate at day 2, blastocyst rate at day 8, and total cell number in the blastocysts. The overall efficiency after SCNT was twice as high in the Oosight group as that in the irradiation group (p<0.05). Comparative TUNEL and semiquantitative RT-PCR assays indicated that the developmental potential of the Oosight group was elevated, with a high expression level of genes related to pluripotency (Oct4 and Nanog), implantation (Interferon-tau), and de novo methylation (Dnmt3A) and a low expression level of proapoptotic (Caspase-3) and stress (Hsp70) genes, compared with the irradiation group. This is the first report to determine the positive effect of the Oosight imaging system on molecular gene expression in the SCNT embryo.
Enucleation of a recipient oocyte is crucial to cloning efficiency. The use of proper measures in enucleation can avoid problems such as aneuploidy and the related, subsequent detrimental effects on later development, genetic interference of the recipient cytoplasm, and possible parthenogenetic activation and embryo development without the participation of a newly introduced nucleus (Dominko et al., 2000). Enucleation may also affect the ultrastructure of the remaining cytoplast, resulting in the decline or destruction of its cellular compartments (Greising et al., 1999).
Among the numerous enucleation methods, we selected Hoechst staining and UV irradiation as a conservative (control group) and Oosight imaging as a new (comparison group) technique. The cloning efficiency of Hoechst-stained oocytes is affected negatively by a simultaneous increase in UV irradiation, and some researchers have indicated that oocytes should not be exposed to UV for more than 30 sec (Smith, 1993). On the other hand, Westhusin et al. (1992) found that exposure to UV irradiation for 10 sec had no effect on embryo viability or the production of live calves. In the present study, we attempted to decrease the UV exposure time of Hoechst-stained oocytes to the briefest time possible (<10 sec), but differences in cloning efficiency remained between the Hoechst staining and UV irradiation system and the Oosight imaging system. Recently, we produced one cloned female Jeju Black cow through the Oosight imaging system from cells of an elite animal dead before 3 years (see Supplementary Figs. 1 and 2). (Supplementary Data are available at www.liebertonline.com/cell/.) The transfer numbers of SCNT embryos were too low to allow valid comparison of the efficiency of cloned animal production between the Hoechst staining and UV irradiation system and the Oosight imaging system. However, TUNEL assay and molecular gene expression analysis results indicated that Oosight imaging may be a more efficient technique than Hoechst staining and UV irradiation.
DNA fragmentation in oocytes associated with apoptotic evidence may indicate poor oocyte quality and lower fertility (Fuijino et al., 1996). Using the TUNEL assay, we confirmed that the DNA fragmentation frequency was significantly lower in the Oosight group than in the irradiation group. In addition, the expression of the proapoptotic gene Caspase-3 was significantly lower in the Oosight group compared with the irradiation group. During SCNT procedures, oocytes were exposed to several instances of heat shock stress during the course of the many mechanical and chemical treatment steps involved in these procedures. Heat shock induces apoptosis in preimplantation embryos in a developmentally regulated manner (Paula-Lopes and Hansen, 2002). We observed that heat shock gene (Hsp70) expression was significantly lower in the Oosight group than in the irradiation group. This result is similar to that of the proapoptotic Caspase-3 gene expression. These results indicate that the Oosight imaging system is a less stressful method for the production of SCNT embryos than the Hoechst staining and UV irradiation system.
Important genes, such as those related to pluripotency (Oct4, Nanog), de novo methylation (Dnmt3A), and implantation (Interferon-tau), which affect the in vitro and in vivo developmental potential of SCNT embryos, were relatively highly expressed in the Oosight group compared with the irradiation group. The octamer-binding transcription factor Oct-4 is a master key regulator that is expressed at the beginning of mammalian embryogenesis. Observed variations in the level and pattern of Oct-4 expression might be responsible for at least some of the problems related to cloning (Boiani et al., 2002). The level of Oct-4 expression might regulate cell lineage commitment in that a critical level of expression is required to maintain pluripotency (Niwa et al., 2002). Nanog is also a crucial pluripotency factor, expressed in ICM cells with Oct-4. The Nanog expression level is directly related to epiblast formation at later stages (Messerschmidt and Kemler, 2010). The present study found that the two pluripotency factors, Oct4 and Nanog, were highly expressed in SCNT embryos of the Oosight group compared with those of the irradiation group.
DNA methylation in embryos changes in a very organized manner to set up imprinting patterns that are vitally important for numerous biological events. Dnmt3A is thought to be essential for the establishment of de novo methylation patterns during gametogenesis and mammalian development (Golbabapour et al., 2011; Okano et al., 1999). Abnormalities observed in cloned animals suggest that imprinted genes and problems related to unsuccessful epigenetic reprogramming might be serious contributors to the failed development of cloned embryos (Beyhan et al., 2007). The Dnmt3A expression level was higher in the Oosight group compared with the irradiation group, although this difference was not significant. This result suggests that the Oosight imaging method may be more helpful in furthering the developmental potential of SCNT embryos than the Hoechst staining and UV irradiation method.
Interferon-tau is exclusively secreted by the trophectodermal cells of blastocysts. It primarily functions in the maternal recognition of pregnancy in cattle (Roberts et al., 1992). Higher mRNA expression levels of Interferon-tau indicate good-quality embryos. In this study, there was a significant difference in the Interferon-tau mRNA expression level between the two groups, suggesting that SCNT embryos of the Oosight group had better developmental potential than those of the irradiation group.
Taken together, the results of this study demonstrate that direct enucleation using the Oosight imaging system has positive effects on the production of SCNT embryos compared with Hoechst staining and UV irradiation. Using real-time imaging with the easy-to-use software of the Oosight imaging system, the MII plate and PB1 of recipient oocytes were easily removed without the damaging membrane or cytoplasmic integrity (Liu et al., 2008). In conclusion, Oosight imaging is an efficient and reliable enucleation technique that could improve the cloning efficiency and developmental potential of SCNT embryos by being less damaging to unknown cellular mechanisms.
Footnotes
Acknowledgments
This study was supported by grants from the Bio-industry Technology Development Program (308008-5); Ministry for Food, Agriculture, Forestry and Fisheries, Next-Generation BioGreen 21 Program (PJ009075); and Cooperative Research Program for Agriculture Science & Technology Development (PJ009103), Rural Development Administration, Republic of Korea.
Author Disclosure Statement
The authors declare that there are no conflicting financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.