
Abstract
Abstract
Despite widespread application of intracytoplasmic sperm injection (ICSI) in human-assisted reproductive techniques (ART), the efficiency of this method is still far from satisfactory in livestock, particularly in the bovine species with its unique sperm condensation. On the basis of the natural chemical structure of chromatin in condensed sperm, we developed a novel combined heparin-reduced glutathione (GSH) sperm pretreatment that improves the efficiency of bovine ICSI via selection of the most appropriate sperm at the time of ICSI. Assessment of sperm DNA integrity revealed that this pretreatment can be considered as a safe and efficient approach for in vitro sperm decondensation when compared to conventional sperm pretreatments with dithiothreitol (DTT). Injection of completely decondensed bull sperm derived from this pretreatment significantly improved fertilization and blastocyst formation rates compared to untreated or intact sperm injection (34.8±2.7 and 29.1±1.5 vs. 12.0±3.2 and 15.9±1.2%, respectively; p<0.05). Real-time PCR analysis revealed that expression of pluripotent and anti-apoptosis markers in blastocysts derived by injection of completely decondensed sperm from heparin–GSH pretreatment were comparable to IVF when compared to the DTT pretreatment and control ICSI groups (p<0.05). The results of this study suggested that the degree of sperm decondensation derived from heparin–GSH pretreatment may affect ICSI efficiency in bovine.
Introduction
During the last stage of mammalian sperm maturation, the sperm nucleus is condensed through replacement of histones with protamines and stabilized by the formation of numerous disulfide bonds between protamines (Calvin and Bedford, 1971; Marushige and Marushige, 1975; Tavalaee et al., 2008). This process has great implications, such as the protection of the sperm's nucleus against mechanical and chemical damages, making sperm heads hydrodynamic, preserving imprinted genes, and regulation of gene expression during spermiogenesis (Braun, 2001; Kuretake et al., 1996; Martins et al., 2004; Sotolongo et al., 2003). This process is reversed during fertilization. However, unlike humans and mice, this process remains incomplete when the critical signaling pathways of sperm–oocyte interactions are subverted by the ICSI procedure in livestock (Malcuit et al., 2006). To overcome this phenomenon, the sperm has to be pretreated by various chemicals or methods either to remove the sperm membrane [freezing-thawing (Kolbe and Holtz, 1999), sonication (Hirabayashi et al., 2005), and Triton X-100 (Lee and Yang, 2004)] or partially reduce the degree of sperm nucleus condensation using dithiothreitol (DTT), as a disulfide bond reducer, with or without sodium dodecyl sulfate (SDS) (Rho et al., 1998a; Suttner et al., 2000). It has been suggested that removal of the sperm membrane improves oocyte activation and embryonic development through increasing the availability of sperm-borne oocyte-activating factor (SOAF) to the cytoplasm of the oocyte (Morozumi et al., 2006), whereas DTT partially decondenses sperm and thereby facilitates male pronucleus (PN) formation (Rho et al., 1998a; Wei and Fukui, 1999). However, the detrimental effects of these pretreatments on paternal genome integrity may adversely affect further embryo development (Perry et al., 1999; Szczygiel et al., 2003).
Reduced glutathione (GSH), the endogenous disulfide bond reducer, has fundamental roles in the oocyte development from germinal vesicle to metaphase II stage and cellular protection against oxidative stresses (Ali et al., 2003; De Matos and Furnus, 2000). GSH plays a critical role in sperm decondensation and male PN formation by reduction of S–S bonds to S–H groups in sperm chromatin structure during fertilization (De Matos and Furnus, 2000; Perreault et al., 1988b). Heparin, a member of the glycosaminoglycans (GAGs) family, is present in both male and female reproductive tracts, reproductive secretions, and gametes (Binette et al., 1996; Reyes et al., 2004; Sloan et al., 1996). It also modulates sperm capacitation and acrosome reaction through binding to its receptors on the sperm plasma membrane (Delgado et al., 1988; Sanchez-Prieto et al., 1996). Importantly, heparin binds tightly to intact nucleosomes and induces the release of histone H1 and protamine, which is a critical step in important for sperm chromatin decondensation (Delgado et al., 1999; Romanato et al., 2003, 2005; Villeponteau, 1992).
The ability of combined heparin–GSH treatment for in vitro decondensation of bull sperm was first introduced by Delgado et al. (2001) for purposes other than ICSI. Subsequently, Lee and Yang (2004) showed that pretreatment of sperm with heparin could improve the efficiency of ICSI in bovine species. However, to our knowledge, there is no report regarding sperm pretreatment with heparin–GSH to improve the efficiency of ICSI in the bovine species. Therefore, the main objective of this study is to introduce combined heparin–GSH as an innate treatment to facilitate sperm decondensation and thereby improving the ICSI fertilization rate. In addition, we showed that sperm selected based on the degree of decondensation significantly impacts fertilization and embryo development post-ICSI.
Materials and Methods
Unless indicated otherwise, all chemicals and media were obtained from Sigma Chemical (St. Louis, MO, USA) and Gibco (BRL, Grand Island, NY, USA) companies, respectively.
Sperm preparation and treatments
Commercial frozen sperm from three different bulls was thawed, pooled together, and washed twice by centrifuging at 400×g for 10 min in 2 mL of HEPES-tissue culture medium 199 (H-TCM) supplemented with 10% fetal calf serum (FCS). Completely motile sperm were obtained by pure sperm 100® (Nidacon, Gothenburg, Sweden) gradients (40:80). For the treatment of sperm with heparin–GSH, 50 μL of completely motile sperm were introduced into 1 mL of Tris medium (100 mM, pH 8.0) supplemented with 80 μM heparin-sodium salt (Hep) and 15 mM L-GSH. Sperm were also treated with 80 μM heparin and 15 mM L-GSH alone, as described by Delgado et al. (2001). Treated sperm were incubated for 7 h at 39°C in 20% O2 and 5% CO2 according to Sanchez-Vazquez et al. (2008). Sperm pretreatment with DTT was carried out according to Rho et al. (1998a). In brief, 50 μL of the motile sperm were introduced into 5 mM DTT diluted in H-TCM supplemented with 10% FCS and incubated for 1 h at 39°C in 20% O2, and 5% CO2. Sperm were washed twice by centrifugation for 10 min at 350×g in 10 mL of H-TCM to remove DTT.
Sperm analyses
Acridine Orange staining
Acridine Orange (AO) staining was performed to evaluate the effect of sperm decondensation on sperm DNA denaturation under different pretreatments. AO is a metachromatic dye that intercalates with double-stranded and single-stranded DNA to form pattern staining from green to orange and red fluorescence. The extent of chromatin integrity posttreatments was assessed by this procedure according to Kheirollahi-Kouhestani et al. (2009).
TUNEL assay
The terminal deoxynucleotidyl transferase dUTP nick end-labeling (TUNEL) assay was carried out to establish the effects of various pretreatments on sperm DNA fragmentation. The assay was conducted via a TUNEL assay kit (Apoptosis Detection System Fluorescein; Promega, Mannheim, Germany) according to the manufacturer's protocol. Red sperm were considered intact, whereas green sperm were considered fragmented. The percentage of sperm DNA fragmentation was assessed in all experimental groups by fluorescence microscopy (Olympus BX51, Tokyo, Japan).
Sperm typing
Sperm typing at the time of ICSI was carried out using image analyzer software (Motic Images Plus 2.0, British Columbia, Vancouver, Canada) on the basis of the size and morphology of decondensed sperm heads. The intact sperm (head size ≤7 μm) was regarded as type I (TI; Fig. 1B1), the partially decondensed sperm (8 μm≤head size ≤11 μm) as type II (TII; Fig. 1C1), and the completely decondensed sperm (head size≥12 μm) was regarded as type III (TIII; Fig. 1D1). The proportions of TI, TII, and TIII after combined heparin–GSH and DTT treatment were determined by microscopic analysis of the slides on by fluorescence microscopy (Olympus BX51, Tokyo, Japan). On each slide, 200 sperm cells were evaluated.
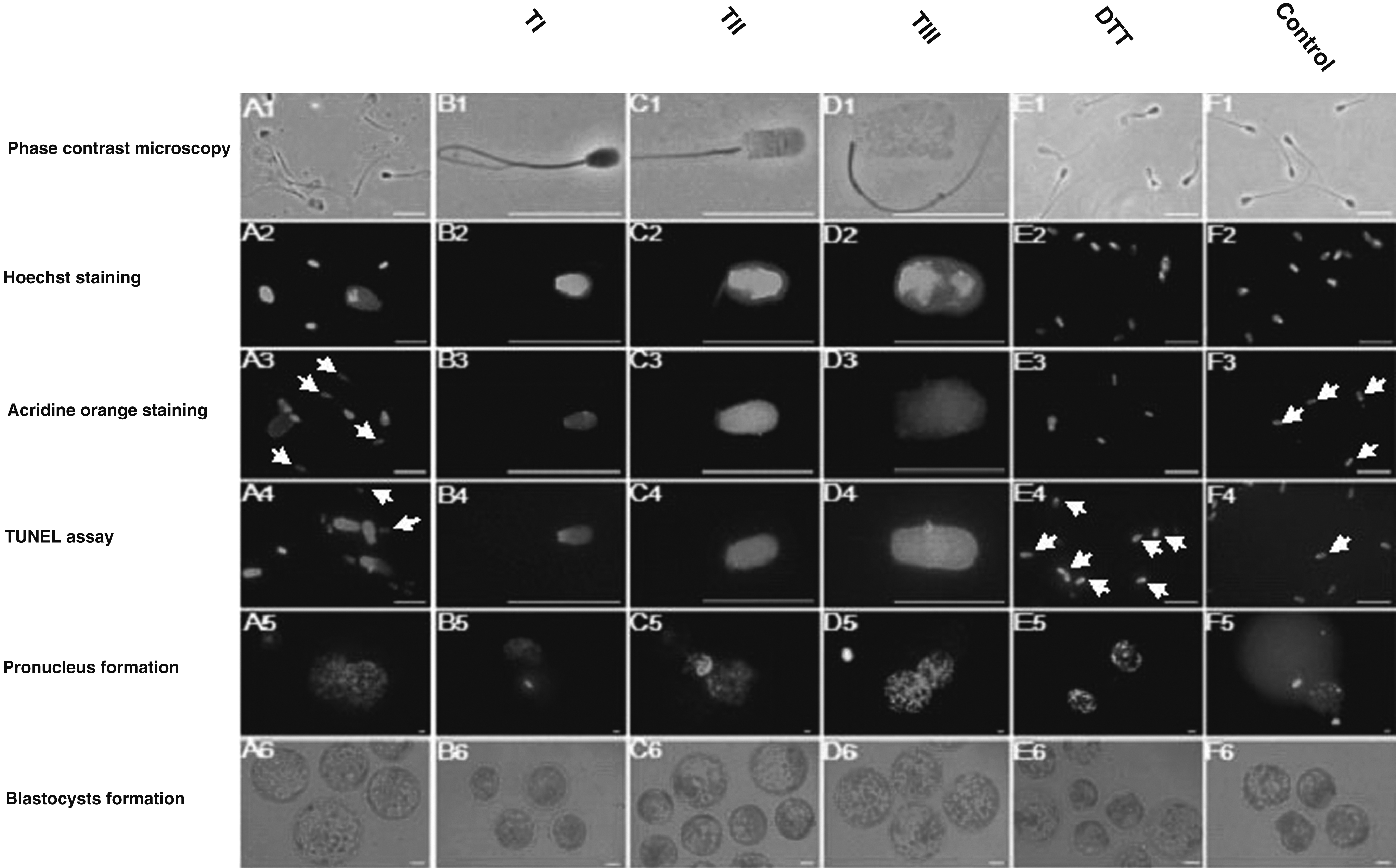
Phase-contrast and fluorescence microscopic analyses. (
Oocyte preparation
The process for oocyte in vitro maturation was as described previously (Moulavi et al., 2006). In brief, cumulus–oocyte complexes (COCs) were aspirated from antral follicles (2- to 8-mm diameter) of abattoir-derived ovaries using 18-gauge needles attached to a vacuum pump (80 mmHg). COCs with homogeneous cytoplasm and more than three layers of cumulus cells were then matured in tissue culture medium 199 (TCM-199) that contained 2.5 mM sodium-pyruvate, 100 IU/mL penicillin, 100 μg/mL streptomycin, 1 mg/mL estradiol-17β, 10 μg/mL follicle-stimulating hormone (FSH), 10 μg/mL luteinizing hormone (LH), 100 ng/mL epidermal growth factor (EGF), 0.1 mM cysteamine, and 10% FCS at 39°C in a humidified atmosphere of 5% CO2 in air.
ICSI
To avoid the mechanical transfer of disulfide bond–reducing agents (DTT or heparin–GSH) via injection pipette, immediately before ICSI the treated sperm in each group were washed twice in 1 mL of H-TCM by centrifuging at 150×g for 10 min. Finally, 3-μL aliquots of precipitate were introduced into 20μL droplets of phosphate-buffered saline (PBS) in ICSI dishes. ICSI was performed using an injection pipette with an inner diameter at the tip of 8 μm for injection of control sperm and ≥12 μm for injection of decondensed sperm. ICSI was carried out as described previously (Shadanloo et al., 2010). Chemical activation of injected oocytes was performed out according to Nasr-Esfahani et al. (2008) with minor modifications. Briefly, 20 min after sperm injection, oocytes were activated using 5 μM calcium-ionophore prepared in H-TCM plus 1 mg/mL bovine serum albumin (BSA) for 5 min in the dark, followed by washing in H-TCM plus 3 mg/mL BSA. For extrusion of the second polar body, activated oocytes were left to rest for 3 h in synthetic oviduct fluid (SOF) medium, and then incubated in 2 mM 6-dimethyl aminopurine (6-DMAP) for 4 h. The presumptive zygotes were then cultured in a sequential formulation of SOF medium (Tervit et al., 1972) at 39°C, 5% CO2, 5% O2, and maximum humidity for 8 days.
Parthenogenetic oocyte activation
The protocol for parthenogenetic activation of matured oocytes was similar to the activation process for ICSI, but to prevent extrusion of the second polar body, the 3-h period between activation with calcium-ionophore and 6-DMAP was omitted. After several washings in H-TCM, activated oocytes were cultured as described for the ICSI group.
In vitro fertilization
The process of oocyte in vitro fertilization (IVF) was as described by Moulavi et al. (2006) with minor modifications. Accordingly, the same semen samples used for sperm treatments and ICSI were also used for IVF. Highly motile sperm were collected using a pure sperm gradient, washed once by centrifugation (70×g for 10 min). Meanwhile, matured COCs were washed twice and transferred in groups of 25–30 per 200-μL drops of fertilization medium under mineral oil. The IVF medium consisted of Fert-TALP (Parrish et al., 1986) supplemented with penicillamine (20 μM), hypotaurine (10 μM), epinephrine (1 μM), and heparin (0.56 μg/mL). COCs and sperm were coincubated for 24 h at 39°C, 5% CO2, and maximum humidity. Subsequently, oocytes were cleaned off from sperm and cumulus cells using vortexing and cultured as described for ICSI oocytes.
Assessment of pronucleus formation
Pronucleus (PN) formation analysis was carried out to determine the efficacy of the activation protocol and to evaluate the efficiency of the male PN formation for each treatment. This was suspected to be the main reason for ICSI failure in the bovine species.
The assessment of PN formation was carried out according to Bevacqua et al. (2010). In brief, at 16 h after initial activation, oocytes were fixed in an acetic acid:ethanol (3:1) solution and incubated for 5 h at room temperature (RT). The staining was performed by incubation of the fixed embryos in 1 mg/mL Hoechst for 5 min. For assessment of PN formation, stained embryos were mounted and analyzed by fluorescent microscope. In each group, the presumptive zygotes were classified as 2PN (formation of both male and female PN), 1PN (have a female PN but with a condensed sperm head), or nonactivated (without male and female PN).
Gene expression
RNA extraction
Total mRNA was extracted using the RNeasy Micro Kit (Qiagen™, Germany), according to the manufacturer's instructions. In brief, at day 8 of embryo culture, 10 blastocysts from each group were pooled, lysed, treated with DNase I, and finally the total extracted mRNA was eluted in RNase-free water and immediately used for cDNA synthesis.
Reverse transcription
Total RNA was used for cDNA synthesis by the RevertAid ™ First Strand cDNA Synthesis Kit (Fermentas™, Germany) according to the manufacturer's recommendation. cDNA syntheses was conducted in a 20 μL final volume that contained: total extracted RNA, 1 μL of random hexamer primer (100 μM), 1 μL of RNase inhibitor (20 U/μL), 4 μL of 5×reaction buffer [250 mM Tris-HCl (pH 8.3), 250 mM KCl, 20 mM MgCl2, 50 mM DTT], 2 μL of dNTP (10 mM), 1 μL of Moloney murine leukemia virus (Mo-MuLV) reverse transcriptase (200 U/μL), and RNase-free water.
The thermostatic program for cDNA synthesis consisted of a 10-min incubation at 70°C, annealing at 25°C for 10 min, and finally reverse transcription at 42°C for 60 min. The samples were then cooled at 4°C.
Real-time PCR
Gene transcription was quantified by quantitative real-time PCR (qRT-PCR) using the Rotorgene 2000 Real Time Cycler (Corbett Research, Sydney, Australia). For each sample, qRT-PCR was performed in triplicate. Control samples (no template control) were also run in parallel with the casework DNA samples.
PCR was conducted by adding 1 μL aliquot of the synthesized cDNA to the 20 μL of PCR mixture that contained 1×SYBR® Premix Ex Taq™ (Takara Bio Inc., Otsu, Japan), 0.4 μM of each specific primer, and RNase-free water. The PCR protocol included an initial step of 94°C (4 min), followed by 40 cycles of 94°C (30 sec), 58–62°C (30 sec), and 72°C (30 sec).
To quantify gene expression levels, the comparative CT method was performed (Gutierrez-Adan et al., 2004). The gene expression pattern of each experimental group was compared with IVF as an indicator. The candidate genes for evaluating the different aspects of embryo quality were: GAPDH, POU5F1 (Oct4), SOX2, VEGF, ATP1A1, HSP40, and GMNN (Table 1).
Transcription factor involved in embryonic stem cell pluripotency (Kirchhof et al., 2000).
Transcription factors involved in the regulation of embryonic development and in the determination of cell fate (Masui et al., 2007).
Creation of new blood vessels during embryonic development (Krussel, et al., 2001).
Glucose transporter, which is critical for early embryo development (Moley, 1999).
Cellular stress-associated factor, which has antiapoptosis effects (Mosser et al., 1997).
Regulation of accumulation of fluid in the blastocoele, differentiation of the TE (Barcroft et al., 2004).
Induction of global histone methylation in the heterochromatin region (Sylvestre et al., 2010).
Housekeeping gene as a reference (Smolkina and Karus, 2004).
Statistical analysis
Cleavage and blastocyst rates were modeled by ArcSin transformation and the transformed data were analyzed by one-way analysis of variance (ANOVA) using SPSS version 17 software (SPSS Science, Chicago, USA, IL). Differences were compared by the Tukey multiple comparison post hoc test. PN formation, and TUNEL assay data (categorical data) were analyzed with the logistic regression procedure of STATA 9 (College Station, TX). All data were presented as means±standard error of the mean (SEM), and differences were considered significant at p<0.05.
Results
Sperm analysis
The results of sperm analysis revealed that decondensation under combined heparin–GSH treatment and injection of more decondensed sperm heads (TIII) (Fig. 1) improved the rate of male PN formation and increased the blastocyst formation rate per injected oocyte (Fig. 2 and Table 2). AO staining demonstrated that under combined heparin–GSH treatment, with increased rate of decondensation from TI to TII and TIII, DNA unwinding via acid treatment increased (Fig. 1). According to the results of the TUNEL assay, under combined heparin–GSH treatment, the proportion of sperm DNA fragmentation in TIII in comparison to control was not significantly increased (Table 3) and the majority of completely decondensed sperm remained intact (Fig. 1A4). However, DTT treatment led to extreme DNA damage in most treated sperms (90%) that invariably affected embryo quality (Table 3, Fig.1E4 and E6). After 7 h of sperm pretreatment with combined heparin–GSH, the proportion of TI was 10.2±1.1%, TII was 29.6±2.3%, and TIII was 61.0±2.1%. However, like Delgado et al. (2001), we also observed no sperm nuclear decondensation when sperm were pretreated with heparin or GSH alone (data not shown). The rate of sperm head decondensation under DTT treatment was rare and there was no visible degree of nuclear decondensation (Fig. 1E1 and E2). The majority of sperm heads folded near their equatorial segments (data was not shown).

Effects of different sperm pretreatments and the degree of decondensation on pronucleus formation in bovine ICSI-derived embryos. (Nonactivated) without male and female PN; (1PN) have a female pronucleus but with condensed sperm; (2PN) formation of both male and female PN. Percentages not sharing a common letter are statistically different (p≤0.05). The number of replicates for each experimental group was 50 fertilized oocytes.
Percentages not sharing a common letter within each column are statistically different (p<0.05).
ICSI, intracytoplasmic sperm injection; TI, injection of intact sperm (head size ≤7 μM); TII, injection of partially decondensed sperm (8 μM<head size ≤11 μM); TIII, injection of completely decondensed sperm (head size≥12 μm) obtained under combined heparin–glutathione (Hep-GSH) pretreatment; DTT, injection of sperm pretreated with dithiothreitol; control, injection of untreated sperm; parthenogenetic, blastocysts derived by chemical activation.
Percentages not sharing a common letter within each column are statistically different (p<0.05).
TUNEL, terminal deoxynucleotidyl transferase dUTP nick end-labeling; TI, Intact sperm (head size ≤7 μM); TII, partially decondensed sperm (8 μM<head size ≤11 μM); TIII, completely decondensed sperm (head size≥12 μm) obtained under combined heparin–glutathione (GSH) pretreatment; DTT, sperm pretreated with dithiothreitol; control, untreated sperm.
Pronucleus formation
The analysis of PN formation revealed that in all experimental groups, the activation protocol induced acceptable rates of oocyte activation that was around 60–80%. This rate is similar to the previously reported rate (Rho et al., 1998a) (see Fig. 2). The rate of nonactivated oocytes was highest and similar to control sperm when TI sperm were used for insemination of oocytes, which were significantly different from TII sperm and DTT-treated sperm (Fig. 2).
As shown in Figure 2, despite the highest rate of activation observed in TII sperm compared to all treatments groups, the efficiency of male PN formation in this group remained lower than TIII sperm. The lowest rate of 2PN oocytes was observed in the TI (7.1±2.1%) and control (12.0±3.2%) groups, which were both significantly lower than the TIII (34.8±2.7%) group. According to Figure 2, the 2PN formation rate increased with the higher order of decondensed sperm used in this study.
In vitro developmental competence of ICSI-derived embryos
The results from ICSI of 852 in vitro-matured oocytes using sperm with different degrees of decondensation (TI, II, III), DTT, and control indicated that the cleavage rates at day 3 postinjection were not significantly different between the groups. We observed the maximum rate of blastocyst formation per cleaved embryo in TIII sperm (29.1±1.5%), which significantly differed from the control sperm (15.9±1.2%). The lowest rates of blastocysts per injected oocyte were observed in the control and TII sperm. The highest rate of blastocyst formation per oocyte in TIII sperm was significantly higher than the other groups, with the exception of DTT sperm (Table 2).
Gene transcription analysis
To determine the effects of various sperm pretreatments on the developmental potency and quality of ICSI-derived blastocysts, the expression patterns of some developmentally important genes were investigated. As indicated in Figure 3, while the POU5F expression rates were not significantly different among blastocysts derived from IVF, parthenogenetic, and TI–III ICSI, its expression in DTT and control-derived blastocysts was significantly lower than TIII and IVF groups. The lowest expression of SOX2, as the second marker of pluripotency, was observed in DTT sperm; however, the expression of only this gene in TIII sperm was comparable to IVF. GMNN is an important reprogramming gene that directs cell lineage commitment of the early embryo toward ICM and TE production (Gonzalez, and Tachibana, 2006; Sylvestre, et al., 2010). Importantly, the expression of the GMNN gene in ICSI-derived blastocysts in TIII sperm was significantly higher than all the other groups, except for the TII sperm group. We observed the lowest rate of GMNN gene expression in DTT sperm, where it was significantly lower than all other groups, except for parthenogenetically derived embryos. Expression of GLUT1, as a postcompaction developmentally important marker, was highest in IVF embryos. In comparison with IVF, GLUT1 expression was significantly lower in all groups, except for control and TIII sperm.

Relative mRNA transcription analysis of in vitro-fertilized, parthenogenetically activated, and bovine ICSI-derived blastocysts. (TI) Blastocysts derived by injection of the intact sperm obtained after combined heparin–GSH pretreatment (head size ≤7 μm); (TII) blastocysts derived by injection of partially decondensed sperm obtained after combined heparin–GSH pretreatment (8 μm<head size ≤11 μm); (TIII) blastocysts derived by injection of completely decondensed sperm obtained after combined heparin–GSH pretreatment (head size≥12 μm); (DTT) blastocysts derived by injection of sperm pretreated with dithiothreitol; (Control) blastocysts derived by injection of untreated sperm; (Parthenotes) blastocysts derived by chemical activation; (IVF) blastocysts derived by in vitro fertilization. Columns not sharing a common letter are statistically different (p≤0.05). For each sample, qRT-PCR was performed in triplicate.
Expression of ATP1A1 as an essential factor for differentiation of the trophectoderm (TE) significantly decreased in DTT sperm compared with other groups, except for TI and control sperm. The expression level of the ATP1A1 gene in TII and TIII sperm was not significantly different from IVF. Despite the differential expression level of the latter two genes, no significant differences in VEGF gene expression were observed between the groups.
Transcription analysis for HSP40 as an anti-apoptosis gene demonstrated that sperm pretreatment with DTT prior to ICSI resulted in the minimum level of HSP40 expression, which was significantly lower than the other groups, except for control sperm. However, the rate of HSP40 expression in TIII, parthenogenesis, and TI sperm was not significantly different from the IVF group. The expression of GAPDH as a reference gene was not significantly different between the experimental groups (Supplementary Fig. S1).
Discussion
The present study shows that combined heparin–GSH pretreatment is a valuable approach for sperm decondensation and provides a unique opportunity to select appropriate sperm for ICSI in the bovine species and thereby improves ICSI efficiency.
The rate of blastocyst formation in TIII was improved slightly in comparison to the DTT group as conventional sperm pretreatments in this study. The rate of blastocyst formation in DTT in this study was close to the rate previously reported for DTT by Rho et al. (1998). Considering the significant improvement in blastocyst rate compared to the control group and the insignificant increase in blastocyst formation compared to the DTT group, one may suggest that further optimization is required to obtain a blastocyst formation rate comparable to the rate observed for parthenogenic activation.
Analysis of the expression of candidate genes, enveloping the different aspects of embryo quality, indicated that in comparison to IVF the gene expression pattern of blastocysts in TIII sperm was more appropriate than the other groups. It is well known that the swollen sperm nuclei in oocytes tend to show more active transcription than condensed nuclei, suggesting that chromatin decondensation is not a merely morphological event and that it may expose the genome to proper reprogramming during the early stage of fertilization, affecting sperm remodeling and transcriptional status in the oocyte cytoplasm (Carmo-Fonseca, 2002; Tamada et al., 2006).
Lack of sperm decondensation in bovine ICSI is a major drawback, particularly when signaling pathways during the sperm–oocyte interaction have been bypassed (Malcuit et al., 2006). Therefore, it can be concluded that the improved candidate gene transcription patterns obtained in TIII sperm were derived from proper sperm chromatin accessibility by oocyte-associated remodeling factors. In contrast, although DTT treatment slightly improved both male PN and blastocyst formation rates, the pattern of candidate genes expression remained low. Moreover, observations by microscopy showed that DTT treatment apparently decreased some morphological aspects of the derived blastocyst. AO staining and the TUNEL assay also supported these results. Using AO-stained sperm, Kosower et al. (1992), in a study on hamsters, reported that the nucleoli of naturally fertilized sperm chronologically changed from green to red during decondensation within the ooplasm and in the PN stage. They suggested that this transition was related to the degree of presence of disulfide bonds. In the light of the above facts, it is possible that reduction of S–S bridges in the sperm chromatin structure that occurred under the combined heparin–GSH sperm pretreatment is a natural process that could be essential for both the initiation of PN formation and progression of reprogramming.
On the other hand, generally under combined heparin–GSH pretreatment, the proportion of sperm DNA fragmentation in comparison to DTT was lower, which could be due to the natural features of this pretreatment and cellular protection of GSH against oxidative stresses (Ali et al., 2003; De Matos and Furnus, 2000). Interestingly, despite complete sperm head swelling in TIII, the percentage of fragmented head sperm was considerably low. It is likely that the most appropriate sperm respond to proper decondensation and form the largest decondensed head or TIII. In this regard, in TII that have partial decondensation, the rate of DNA fragmentation was higher. TUNEL assessments for DTT were in concordance with the findings of Ward and Ward (2004), who have shown that DTT treatment destabilizes the structural protein of sperm nuclei, making them more susceptible to DNA damage.
In conclusion, the results of the present study indicate for the first time that the degree of bovine sperm decondensation at the time of ICSI could affect its efficiency. This may have practical implications for other species. We have shown that combined heparin–GSH treatment provides a unique opportunity to select the most appropriate sperm for bovine ICSI because it fully supports male PN formation and in vitro embryo development.
Footnotes
Acknowledgment
The present study was funded by a grant from Royan Institute of IRI. The authors would like to express their gratitude to Royan Institute for their full support.
Author Disclosure Statement
The authors declare that there is no conflict of interest in this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.