
Abstract
Abstract
The somatic cell nuclear transfer (SCNT) procedure requires nuclear remodeling to return differentiated somatic nuclei to the totipotent undifferentiated stage. We hypothesize that mechanical constraints might occur upon SCNT and thereby affect nuclear remodeling. Therefore, we analyzed the nuclear structures upon SCNT using as donors either wild-type fibroblasts with a dense vimentin network or vimentin-deprived cells [embryonic stem cells (ESCs) and fibroblasts invalidated for vimetin]. We demonstrated that following nuclear transfer of wild-type fibroblasts, vimentin intermediate filaments (IFs) persisted around the transplanted nuclei and 88% of them presented severe distortions. We also showed that the presence of vimentin filaments in the reconstructed embryos was correlated with DNA damage, as evidenced by γH2A.X foci. On the other hand, when ESCs or vimentin-null (Vim−/−) fibroblasts devoid of IFs were used as nuclear donors, no nuclear distortion and less DNA damage were observed. Altogether we believe that the introduction of vimentin into recipient oocytes during SCNT induces a mechanical constraint on the transplanted nucleus that is responsible for nuclear distortions and DNA damage. This could lead to incomplete reprogramming that would be detrimental to further embryonic development.
Introduction
It is now generally accepted that the development of cloned embryos receiving well-differentiated donor nuclei is less successful than with less differentiated ones (Heyman et al., 2002; Kato et al., 2000; Li et al., 2003; Miyara et al., 2006). For example, a 10- to 20-fold higher rate of development from blastocyst to term is observed in embryos reconstituted using embryonic stem cell (ESC) nuclei compared to somatic cell nuclei (Humpherys et al., 2002; Rideout et al., 2000). The differences between embryonic cells, ESCs, and somatic cells include the framework of intermediate filaments (IFs) (for review, see De Pianto and Coulombe, 2004). During SCNT, the foreign nucleus does indeed transmit the genome, but it also carries a variety of cytoplasmic factors into the recipient oocyte, including cytoskeletal elements that might induce mechanical constraints.
IFs are a significant component of the cytoskeleton and nuclear membrane in most types of eukaryotic cells (for review, see Erickson et al., 1992; Fuchs and Weber 1994; Steinert and Roop, 1988). One of the major functions fulfilled by IFs is to contribute to the maintenance of cellular integrity; IFs act as a scaffold that binds and regulates the activity of several effector proteins. The IF network is particularly stable and insoluble when compared with microfilaments and microtubules. IFs form an extensive and interconnected three-dimensional network that is distributed throughout the cytoplasm and functionally connected to the nuclear matrix, interacting closely with the nuclear lamina and its constituent filaments (Capco et al., 1982; Tolstonog et al., 2002; Traub and Shoeman, 1994). IFs vary in a cell–tissue and differentiation-dependent manner (for review, see Franke et al., 1982; Fuchs and Weber, 1994). The IF vimentin is expressed by endothelial and mesenchymal cells, such as fibroblasts, and appears only 7 days after fertilization in mouse embryos, in newly forming mesoderm. In fact, oocytes, early embryos, and ESCs are devoid of vimentin or any other IF (for review, see De Pianto and Coulombe, 2004). Therefore, we hypothesized that, during SCNT, the introduction of IFs into the recipient oocyte cytoplasm might promote mechanical constraints on the donor nucleus and thereby alter its remodeling.
On the other hand, it has emerged from different studies that nuclear organization and chromatin structure could potentially regulate genome function (Misteli, 2007; Misteli and Soutoglou, 2009; Thomas et al., 2002). A well-known marker for the analysis of DNA double-strand breaks (DSBs) and hence chromatin integrity is the phosphorylation of Serine139 in the H2A.X histone, termed γH2A.X (Paull et al., 2000). Microscopically visible γH2A.X foci were indeed initially described as the mark initiating the DNA DSB pathway to recruit DSB repair factors (Rogakou et al., 1998). Previous results have shown that in NT embryos micromanipulation with strong piezoelectric strength could induce DNA damage (revealed with γH2A.X foci) in reconstructed embryos, as well as a lower rate of development to term (Yu et al., 2007). Taken together, these data suggest that the nuclear distribution and number of γH2A.X foci will provide information on the DNA quality of NT embryos and help to predict SCNT efficiency.
Therefore, the aim of this study was to compare nuclear behavior after NT using ESCs (deprived of IFs) and fibroblasts with a dense vimentin IF network through immunofluorescent detection of vimentin. To further test the involvement of IFs in the nuclear remodeling after SCNT, we used vimentin-null fibroblasts. We saw that when wild-type fibroblasts were used as donors, the nuclei appeared distorted, which was not the case with ESCs or vimentin-null fibroblasts. We were able to demonstrate that when vimentin is maintained around the nucleus, it mechanically disturbs nuclear remodeling and induces DNA DSBs as shown by γH2A.X localization analysis.
Materials and Methods
All experiments involving animals were carried out according to European regulations of animal welfare.
Cell culture
The fibroblasts were derived from day-14 129S2/SvPas mouse fetuses. They were routinely cultured in Dulbecco's modified Eagle medium (DMEM +4.5 g/L glucose+Glutamax), supplemented with 15% fetal calf serum (FCS) and 0.1 mM β-mercaptoethanol. The Vim−/− fibroblasts were derived from day-14 Vim tm1Cba/Vim tm1Cba 129S2/SvPas fetuses (a generous gift from Dr. Emma Colucci-Guyon; Colucci-Guyon et al., 1994) and were cultured as described above. We did not detect any histological differences between the wild-type and mutant mice, as previously described (Colluchi-Guyon et al., 1994). Using immunofluorescence (Fig. 1A) and western blotting (not shown), we confirmed that these Vim−/− cells expressed neither vimentin nor other IFs. For the NT procedure, fibroblast cells were used at passages 3–4 and kept at confluence for 2 days to synchronize at the G0/G1 stage by contact inhibition.
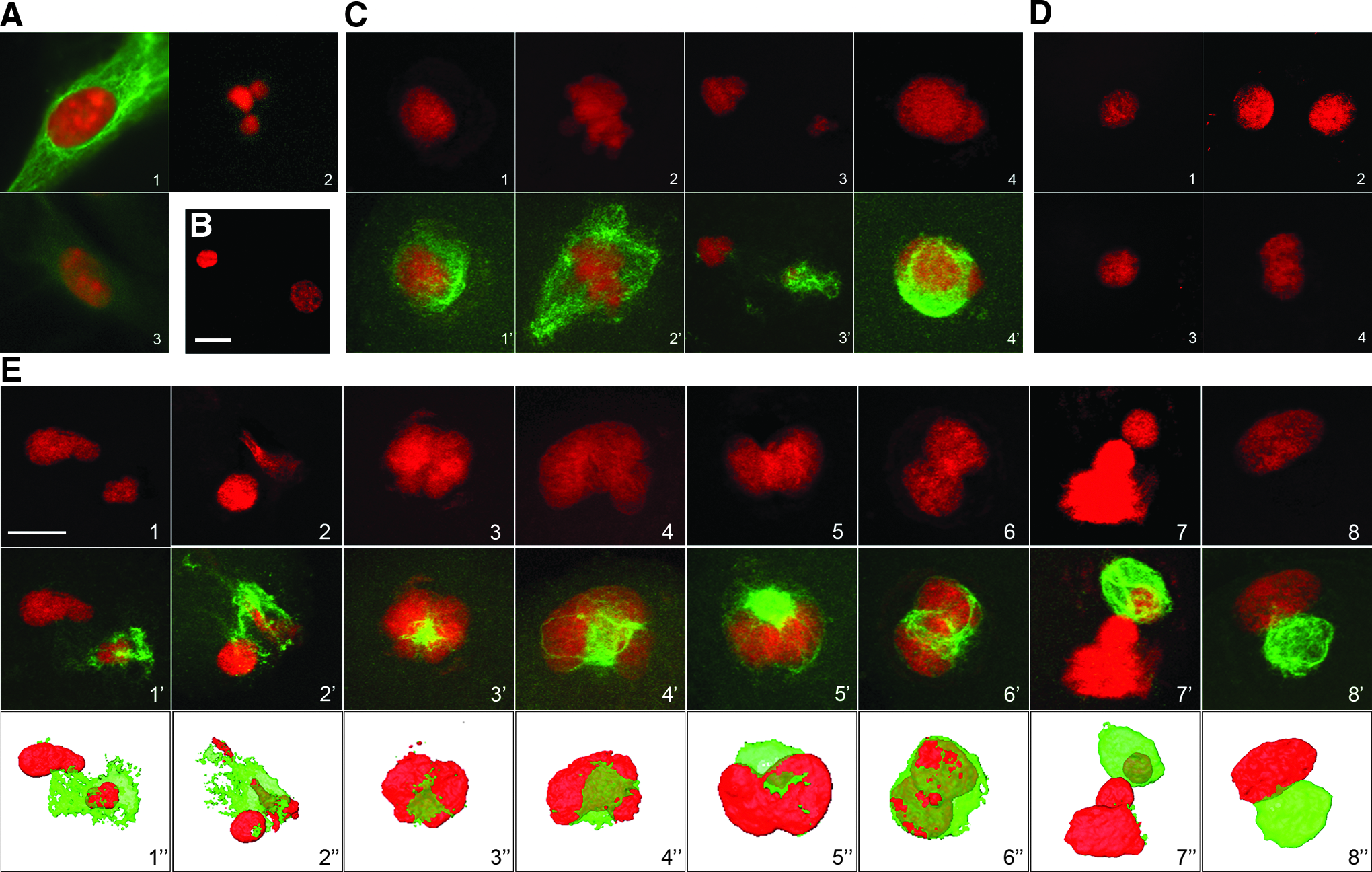
Nuclear behavior and immunolocalization of vimentin after somatic cell nuclear transfer. Each sample was double-stained for vimentin (green) and DNA (red). (
The ESCs (line R1, a generous gift from Dr. A. Nagy, Mount Sinai Hospital, Toronto, Canada; Nagy et al., 1993) were derived from 129/SvJx129/Sv mouse blastocysts. As described by Wakayama et al. (1998), they were routinely cultured on mitomycin C-inactivated embryonic fibroblasts in DMEM+4.5g/L glucose+Glutamax supplemented with 20% FCS, 500 U/mL leukemia inhibitory factor (LIF; Esgro), and 0.1 mM β-mercaptoethanol. For NT, the ESCs were used at relatively low passage (15–20) to avoid any karyotypic changes and were synchronized by an overnight incubation with 0.02 μg/mL demecolcin. The mitotic cells were then collected, washed twice with culture medium, and incubated for 1–2 h to allow resumption of the cell cycle before use. G1 cells were identified as small doublets (Zhou et al., 2001).
Collection of mouse oocytes and NT
NT was performed as described previously (Maalouf et al., 2009; Zhou et al., 2001). The oocytes were prepared by inducing the superovulation of 8-week-old C57Bl/6×CBA/J F1 mice through injection of equine chorionic gonadotropin (eCG; 7 IU) and human chorionic gonadotropin (hCG; 5 IU). Oocytes were collected from the oviducts 13 h after hCG injection. Cumulus cells were removed with hyaluronidase (300 IU/mL), and oocytes were washed with M2 medium (Sigma) several times. They were then cultured in M16 medium (Sigma) at 37.5°C, under 5% CO2 in air. All manipulations (Nikon-Narishige Micromanipulators MO-188; Nikon, Tokyo, Japan) were performed under differential interference contrast [(DIC)-equipped Olympus IX70] at 20× (objective) magnification. The oocytes were incubated for 5 min at 37.5°C in M2 medium containing 5 μg/mL cytochalasin B, and the chromatin spindle was then removed as previously described by Zhou et al. (2001). Donor nuclei were derived from ESCs or fibroblasts by gentle aspiration in and out of the injection pipette (inner diameter 7–8 μm) and were then microinjected into the cytoplasm of the enucleated oocytes. Only small cells were selected as nuclear donors. The reconstituted embryos were incubated for 2 h in M16 medium before activation.
Oocyte activation, embryo culture and development
Reconstituted embryos were activated by incubating them for 6 h in Ca2+-free CZB medium containing 10 mM Sr2 and 5 μg/mL cytochalasin B. Embryos with visible nuclei were considered to be activated. They were then transferred into fresh M16 medium and cultured at 37.5°C under 5% CO2 up to the blastocyst stage (4 days).
To evaluate developmental potential, SCNT blastocysts were transferred into pseudopregnant (C57Bl/6×CBA) F1 females, and fetuses were dissected at day 10. The morphology of these fetuses was recorded as “normal” if turning had occurred and if somites as well as a beating heart were present.
For naturally in vivo–fertilized embryos, superovulated females were mated with male mice at the time of the hCG injection, The collection and culture of these embryos were carried out according to the same method as previously described (Maalouf et al., 2009).
Immunocytochemistry
For vimentin immunodetection, reconstituted embryos and cells grown on glass coverslips (ESCs, fibroblasts) were fixed in Recta Pur methanol (Prolabo, France) at −20°C for 10 min. The samples were then washed three times in phosphate-buffered saline (PBS). For γH2A.X immunodetection, reconstituted embryos were fixed with 2% paraformaldehyde (PFA) in PBS for 30 min at room temperature and permeabilized with 0.5% Triton X-100 (30 min, room temperature).
All samples were blocked with 2% bovine serum albumin (BSA) in PBS for 1 h and then incubated for 2 h with the first antibody [goat polyclonal anti-vimentin antibody; diluted 1/300; Sigma; rabbit polyclonal anti-phospho-histone H2A.X (Ser 139), antibody; diluted 1/300; Cell Signalling] and washed extensively for 1 h in PBS-BSA. The samples were then incubated for 1 h in fluorescein isothiocyanate (FITC)-labeled anti-rabbit immunoglobulin G (IgG) or anti-goat IgG (Biosys) diluted 1/500 in PBS–BSA and exposed for a few minutes to propidium iodide (10 mg/mL; Invitrogen, France) before mounting with a PBS–glycerol mixture (City University, London).
For vimentin immunodetection, 40 one-cell–stage embryos reconstituted with fibroblasts, were collected between 30 min to 2 h after injection or 1 h after activation (two experiments). Embryos reconstituted with fibroblasts (n=91, seven experiments), with Vim−/− fibroblasts (n=34, three experiments) and ESCs (n=20, three experiments) were analyzed by immunocytochemistry 2 h after activation. In vitro-fertilized (IVF) embryos at 4 h postfertilization were used as controls (n=34, two experiments). Confocal microscopy was performed using a Zeiss LSM 510 confocal laser scanning microscope equipped with an oil immersion objective [Plan Apochromatic 63× numerical aperture (n.a.) 1.4], with 488- and 535-nm wavelength lasers. Image analyses were performed using the LSM 5 browser and ImageJ and Amira software.
For γH2A.X immunodetection, embryos reconstituted with ESCs (n=23), with Vim−/− fibroblasts (n=21), and with wild-type fibroblasts (n=20) were analyzed at the two-cell stage (20 h after activation); in vivo–fertilized embryos were used as controls (n=15) (three experiments). Embryos were analyzed using a Zeiss Axiovert 200M inverted microscope equipped with the Apotome fluorescence module and a B/W digital camera for imaging. The images were captured and merged using AxioVision software (Zeiss). Quantitative analyses of γH2A.X levels were performed using ImageJ software as follows: The area of each nucleus was outlined manually on the DNA counterstaining images, the area was pasted on the γH2A.X images, and the mean fluorescence intensity was then measured (it should be underlined that all images were captured with equal acquisition times).
Results
NT with wild-type fibroblasts
When wild-type fibroblasts were analyzed with anti-vimentin antibody prior to NT, we observed a dense vimentin network distributed throughout the cytoplasm as previously described (Coulombe et al., 2004) (Fig. 1A1). These fibroblast nuclei were then observed following NT, between 30 min postinjection and 2 h postactivation.
During the first hour after injection, the somatic nucleus remained intact and IFs were still located around the nucleus, forming a dense interconnected network (Fig.1C1). Between 1 h and 2 h after the injection (n=40), PCC occurred in 80% of the embryos (Fig. 1C2). In these embryos, the IF network was located around the condensed chromosomes (80%) or around the intact nucleus (20%). One hour after activation, the condensed chromatin started to decondense and separate, forming two deformed pronucleus-like structures of unequal size (Fig. 1C3) or one pronucleus-like structure with an irregular shape (Fig. 1C; 4). The IF network was always located around the nuclei, forming a cage (Fig. 1C3,4).
Two hours after activation, the pronucleus-like structures had formed and swollen (n=91). We obtained 66% of embryos with one pronucleus-like structure and 34% with two pronucleus-like structures (Table 1). In most cases, the transferred nucleus was enclosed within an interconnected three-dimensional cage of vimentin. All of the surrounded pronucleus-like structures presented a distorted shape (Fig. 1E), unlike the circular pronuclei observed in fertilized embryos (Fig. 1B). Among the 91 reconstituted embryos thus analyzed, we determined that 88% of the nuclei were distorted and surrounded by IFs (Table 1; Fig. 1E). Some of these embryos displayed two deformed pronucleus-like structures of unequal size (Fig. 1E1,2), whereas others presented a formation of chromatin lobes trying to extrude from the filament cage (Fig. 1E3,4) or presented severe torsions (Fig. 1E5,6). In some embryos, part of the chromatin had escaped from the IF cage, probably inducing a scission (Fig. 1E7). When the nucleus was surrounded by the IF cage, its swelling was altered so that it could not escape the vimentin cage as highlighted by the 3D representations (Fig 1E; bottom row).
We observed that only 34% of reconstituted embryos presented two pronucleus-like structures. Among the 91 reconstituted embryos, 88% displayed an irregular shape.
PN, pronucleus.
In a few cases (12%), the nuclei were able to escape from the IF network; in this situation, the shape of the nucleus seemed to be more regular and the pronucleus-like structure lay outside the vimentin network (Fig. 1E8).
NT with vimentin-deprived cells
We next performed NTs using mouse ES cells and vimentin-null fibroblasts (Vim−/−). Prior to NT, the ESCs and Vim−/− fibroblasts were, as anticipated, negative with anti-vimentin antibody (Fig. 1A2,3). We did not detect any morphological difference between wild-type and mutant (Vim−/−) fibroblasts cultured in vitro. Using western blotting (not shown) and immunofluorescence (Fig. 1A3), we were able to confirm that no vimentin filament or other IFs were expressed in these fibroblasts, as previously described (Colucci-Guyon et al., 1994).
Following NT, embryos reconstituted with ESCs did not present any IF network or nuclear distortion (Fig. 1D1,2). When NT was performed with Vim−/− fibroblasts, the pronucleus-like structure presented a regular, circular shape as with ESCs (Fig. 1D3,4). During these SCNT experiments, we also noticed that the cytoplasm of the Vim−/− cells was more flexible and supple when compared to wild-type fibroblasts, thus enabling easier nucleus recovery and injection, as with ESCs.
DSBs induced by IFs
Because cell type and IF presence appeared to influence nuclear shape following NT, we investigated whether this could be associated with nuclear damage. Therefore, we explored the presence of DNA DSBs under these different experimental conditions. It is well known that in response to DNA damage, a rapid modification of histone composition within the chromatin leads to accumulation of phospho-H2A.X on DSBs (Sedelnikova et al. 2002; Yu et al 2007) that can be detected with anti-γH2A.X-specific antibodies. In the donor cells, no γH2A.X labeling was detected prior to SCNT, as previously described (not shown; Rogakou et al., 1998). We next explored the presence of γH2A.X in two-cell-stage embryos as previously performed by Yu et al. (2007). Very few γH2A.X-positive foci were found in two-cell in vivo–fertilized embryos used as controls (n=15; Fig. 2a) as expected. By comparison the number of γH2A.X-positive foci increased in all cloned embryos (Fig. 2b–d and histogram). Remarkably, labeling was equivalent in cloned embryos reconstituted with Vim−/− fibroblasts (n=21; Fig 2c) and with ESCs (n=23; Fig. 2b). By contrast, labeling increased dramatically in embryos reconstructed with wild-type fibroblasts (n=20; Fig. 2d). These observations indicate that the presence of IFs was correlated with the formation of DSBs upon SCNT.
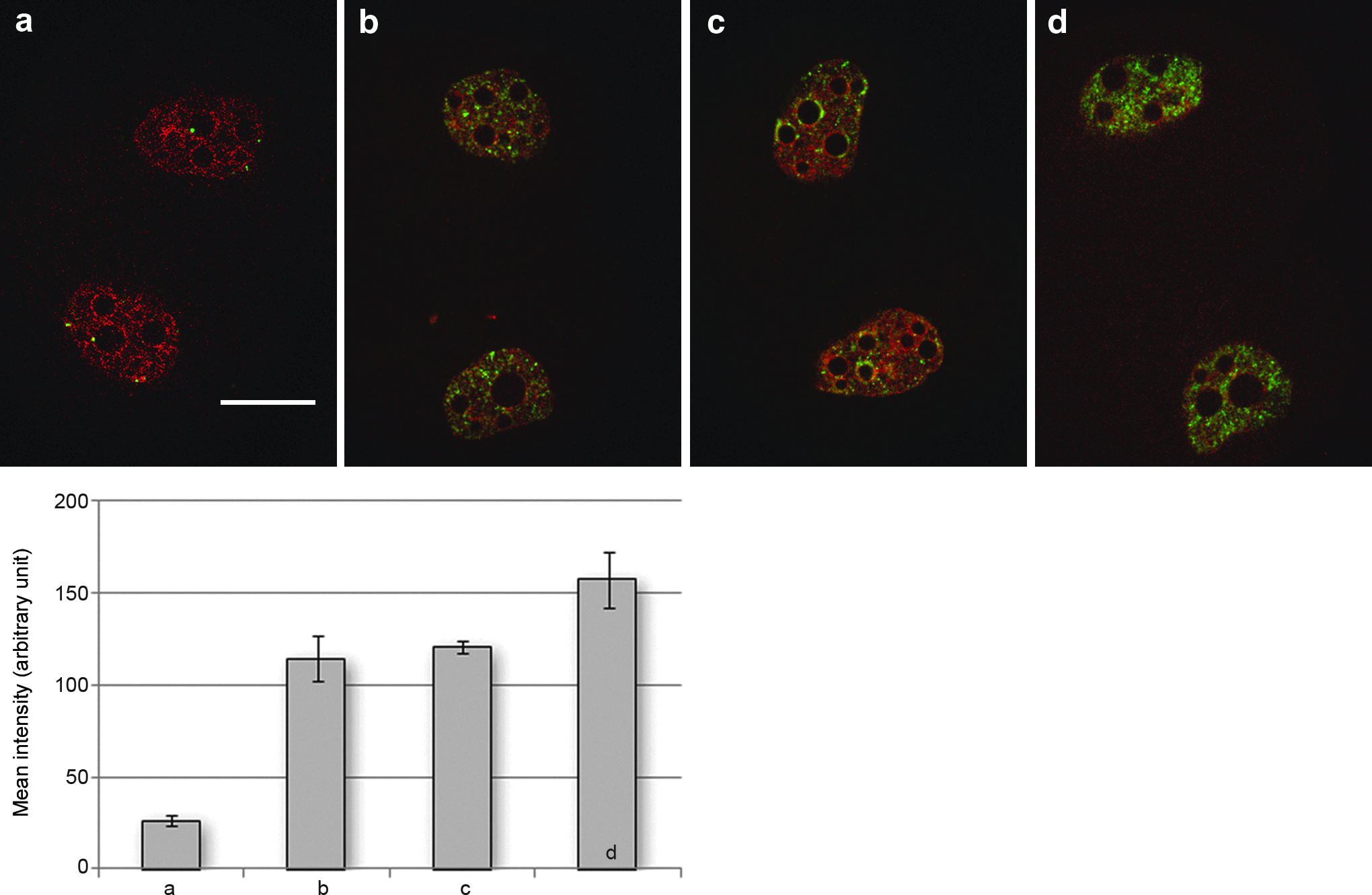
DNA double-strand fragments in two-cell embryos revealed by γH2A.X staining. Embryos were double stained for γH2A.X (green) and DNA (red) at the two-cell stage. In vivo–fertilized embryo (
Development after SCNT with wild and Vim−/− fibroblasts
Finally, we evaluated cleavage rates, blastocyst formation, and implantation after NT of ESCs and wild and Vim−/− fibroblasts. In all groups, the cleavage rate was high, ranging from 70% to 87%, with no statistically significant difference between the wild and Vim−/− fibroblast-derived embryos (Table 2). Remarkably, differences appeared at the blastocyst stage. Fewer NT embryos were reaching this stage with Vim−/− fibroblast donors than with wild-type ones (12.5% vs. 20.6%, p<0.05). We also noted that fewer NT embryos derived from Vim−/− fibroblasts developed to the day-10 normal fetus (as defined in Material and Methods), than those derived from wild-type fibroblasts (4% vs. 10%, p<0.05). However, numbers of implantation sites and live fetuses were very low in both cases, especially in comparison to ESC-derived embryos, as could be expected with SCNT (Table 2).
Chi-squared test gives statistical difference (p<0.05) between all groups at the blastocyst and fetus stages. At the two-cell stage, there is however no statistical difference between Vim−/− and wild-type fibroblasts.
Discussion
In this study, we showed that when fibroblasts were used as nuclear donors for SCNT, the transplanted nuclei remained enclosed in a dense vimentin network that induced important morphological distortions and DNA damage, affecting the foreign nucleus.
Two main procedures are used to transfer donor nuclei into enucleated oocytes—electrofusion and piezoelectric microinjection. In the first case, the donor cell membrane is fused with the oocyte membrane by an electric current, whereas in the latter case, the cytoplasmic membrane of the donor cell is broken and the donor nucleus is directly injected into the oocyte cytoplasm. The electrofusion method is used extensively in most species, and the piezoelectric microinjection method is mainly applied in rodents. The data presented here indicate that the solid vimentin IF network remained strongly bound to the transferred nucleus during SCNT. Furthermore, when vimentin-null fibroblasts were used as nuclear donors, we observed that the cytoplasm seemed to be more flexible when compared with fibroblasts containing vimentin, thus enabling easier nucleus recovery and injection, as with ESCs. Altogether this suggests that the piezoelectric microinjection method was not sufficient to remove IFs from around the fibroblast nuclei, probably because of their sturdiness and their interaction with the nuclear membrane. On the other hand, if electrofusion implies the transfer of the whole cell into the oocyte cytoplasm, the whole IF network of the donor cell would probably be left even more intact.
Following injection and activation, the foreign nucleus needs to be remodeled to reach a state similar to that of a pronucleus. One of the more notable events during this process is the dramatic swelling of the foreign nucleus. It appeared that the vimentin network, forming a solid cage around the nucleus, impeded this swelling process and induced severe nuclear distortions. Likewise, the IF network was also probably responsible for the formation of pronuclei of different sizes. On the other hand, only a small percentage of embryos with two pronucleus-like structures were observed during this study (34%). We hypothesized that the vimentin cage impeded chromosomal separation after PCC and thus induced formation of single pronucleus-like structure. Moreover, only nuclei that were able to escape the IF network displayed a circular regular shape. When NT was performed with IF-deprived ESCs or vimentin-null fibroblasts, no nuclear distortion was observed. We concluded that vimentin IFs introduced during SCNT and persisting around the transplanted nucleus were directly responsible for the nuclear distortions seen when fibroblasts were used as nuclear donors.
We postulated that the nuclear distortions observed after fibroblasts NT could be detrimental to the chromatin structure. To evaluate chromatin integrity, we analyzed the nuclear distribution and number of γH2A.X foci in NT embryos at the two-cell stage. Indeed, the phosphorylated form of H2A.X, which appears in response to the appearance of DNA DSBs, acts both as a DNA damage sensor and as a signal that prevents cells with damaged DNA from entering mitosis (Rogakou et al., 1998; Sedelnikova et al., 2002). When DSBs caused by genotoxic factors occur, cell cycle checkpoint mechanisms function to stop the cell cycle until all DSBs are repaired. Thus, looking at the nuclear distribution and number of γH2A.X foci was a way to obtain information on the DNA quality in our NT embryos (Yu et al 2007). We demonstrated that whichever cells were used as donors for NT—ESCs, Vim−/− fibroblasts, or wild-type fibroblasts—more γH2A.X foci were detected at the two-cell stage than in control in vivo–fertilized embryos. Nevertheless, the rate of embryos affected by DSBs (evidenced by γH2A.X foci) was clearly higher when wild-type fibroblasts were used for SCNT. This could be linked to the nuclear distortion observed in these SCNT embryos. Indeed if the transplanted nucleus is subjected to strong shear forces produced by the IF network, its DNA double-stranded structure may be altered. Altogether this suggests that the use of fibroblasts as nuclear donors for transfer is directly correlated to nuclear distortion and important DNA breaks that could also be linked to inadequate remodeling. However, there are a number of other parameters that could induce DSBs, including manipulation. Indeed, we previously demonstrated in reconstituted embryos that piezoelectric injection can induce DSBs on its own (Yu et al., 2007). In the present study, we used the lowest piezoelectric strength possible to avoid this bias.
On the other hand, when vimentin-null fibroblasts were used, the transferred nucleus never appeared to be distorted and γH2A.X labeling was similar to that seen with ESCs. Once again this suggests that the presence of IFs was correlated with nuclear distortion and chromatin alterations. Therefore, we postulate that the cytoskeleton, and particularly IFs, applies forces that modulate nuclear shape, probably through contacts between the nuclear envelope and cytoplasmic IFs (Herman and Aebi, 2004; Tolstonog et al., 2002; Traub and Shoeman, 1994). Of particular interest is the idea that heterochromatin assembly could be coupled to a nuclear scaffold, which in turn connects it to the nuclear envelope (Cremer, 2007; Nelson et al., 1986; Nickerson, 2001; Traub and Shoeman, 1994; Zhang et al., 2005).
It should be noted that we obtained 10% of live fetuses with wild-type fibroblasts under conditions that induced nuclear distortion. We could hypothesize that the repair machinery acted efficiently and enabled embryos with distorted nuclei to overcome IF effects and DNA damage. It could also be that the 12% of embryos that were not distorted after SCNT corresponded to the embryos that were able to develop further. However, when Vim−/− fibroblasts were used as nuclear donors, embryonic development was not enhanced and remained very limited. Vimentin starvation did not promote better development. It is possible that the absence of IF from Vim−/− fibroblasts might have affected other processes. For example, IF deprivation might modify regulation and/or expression of other cytoskeletal elements, such as tubulin and actin, that could then be detrimental to the reprogramming of the transplanted nuclei. Indeed, because these cytoskeleton elements are really important during cellular compaction at the eight-cell stage and differentiation at the blastocyst stage, we could postulate that their deregulation might contribute to a poorer development.
Finally, it is known that IFs in mitotic somatic cells are partially depolymerized through phosphorylation, especially by the M-phase promoting factor (MPF), allowing cellular division (for review, see Coulombe and Wong, 2004; Inagaki et al., 1996). During the SCNT procedure, although the donor nuclei are transplanted in metaphase cytoplasm with active MPF, we did not observe major IF depolymerization. We hypothesize that oocyte MPF activity was insufficient to promote enough vimentin phophorylation and thus depolymerization of the foreign vimentin. On the other hand, MPF is not the only kinase required for IF depolymerization (Erikson et al., 1992; Inagaki et al., 1996). The oocyte, which is usually devoid of IF, probably does not contain all the machinery required to promote IF phosphorylation and depolymerization.
In conclusion, our observations could have important consequences regarding the choice of SCNT strategies. Indeed, studies on chromatin remodeling after NT need to take into account the mechanical constraint. The link between chromatin structure and the surrounding cytoskeleton should be studied more, particularly regarding IFs, because of their sturdiness and their functional connections to the nuclear matrix. It is also necessary to consider the regulation of cytoplasmic factors present in the recipient oocyte (especially MPF), which might be able to assist in IF depolymerization.
Footnotes
Acknowledgments
We are thankful to Dr. E. Colluchi-Guyon for providing Vim−/− cells. We thank Dr. Pierre Adenot for advice and use of the INRA MIMA2 platform. We thank UEAR for animal care and embryo transfer. We are indebted to J.P. Renard for fruitful discussions and critical reading of the manuscript. This work was supported by an ACI Phase grant from INRA (Institut National de la Recherche Agronomique).
Author Disclosure Statement
The authors declare that there is no conflict of interest that would prejudice the impartiality of this scientific work.