
Abstract
Abstract
Adult cardiomyocytes have little ability to regenerate, thus cardiac regeneration therapy represents a potential method for treating severe heart failure. Human amniotic mesenchymal cells (hAMCs) have the potential to be a useful cell source for cardiac regeneration therapy. We attempted to isolate stem cells from hAMCs and differentiate them into cardiomyocytes. Nanog promoter-Cre plasmid and cytomegalovirus (CMV) promoter-loxP-STOP-loxP-Red-puror plasmid were co-transfected into immortalized hAMCs (iHAMs). Nanog-positive iHAMs were treated with 5-azacytidine (5-aza), trichostatin A (TA), activin A (AA), and bone morphogenetic protein-4 (BMP-4), or co-cultured with murine fetal cardiomyocytes for cardiomyocytes differentiation. Isolated Nanog-positive iHAMs were analyzed by quantitative RT-PCR and immunofluorescent staining before and after differentiation. Expression of Nanog, Oct3/4, Sox2, and Klf4 was significantly higher in Nanog-positive than in Nanog-negative iHAMs. Nanog-positive iHAMs were stained for Nanog and Oct3/4 in the nucleus. Nanog-positive iHAMs treated with 5-aza expressed Nkx2.5, GATA-4, human atrial natriuretic peptide (hANP), cardiac troponin T (cTnT), myocin light chain (Mlc)-2a, Mlc-2v, β-myosin heavy chain (β-MHC), hyperpolarization-activated cyclic nucleotide gated channels (HCN)-4, and inwardly rectifying potassium channels (Kir)-2.1. Although Nanog-positive iHAMs treated with TA, AA, or BMP-4 expressed several cardiac markers, no contraction was observed. Co-cultured Nanog-positive iHAMs with murine fetal cardiomyocytes spontaneously contracted in a synchronized manner and expressed the cardiac markers. In conclusion, Nanog-positive hAMCs with characteristics of stem cells were isolated and differentiated into cardiomyocyte-like cells, suggesting that these isolated hAMCs could be a useful cell source for cardiac regeneration therapy.
Introduction
We have reported that human amniotic mesenchymal cells (hAMCs) have some characteristics of cardiomyocytes (Zhao et al., 2005). hAMCs can be easily obtained from discarded tissues at parturition, thus resolving ethical issues. In addition, hAMCs appear to be relatively resistant to immune rejection because of their immature immune system (Miki and Strom, 2006; Toda et al., 2007; Walther et al., 2009). Therefore, we think that hAMCs have the potential to be a cell source of cardiac regeneration therapy. However, it has not been reported that hAMCs could differentiate into functional cardiomyocytes, probably because the stem cells existing in hAMCs had not been isolated. It has been reported that Oct3/4 and Sox2 were already expressed in almost all human amniotic cells. However, Nanog, which is described as one of the factors for maintaining stemness along with Oct3/4 and Sox2 (Kuroda et al., 2005; Rodda et al., 2005; Yates and Chambers, 2005), is expressed only in limited cells (Miki and Strom, 2006). On the basis of this fact, we focused on Nanog. In this study, the hAMCs in which Nanog promoter is working in the nucleus were named Nanog-“positive” hAMCs. We attempted to isolate Nanog-positive hAMCs by using the Cre/loxP system and then analyze their characteristics. Furthermore, we investigated the differentiation of Nanog-positive hAMCs into cardiomyocytes after treatment with chemical agents, such as 5-azacytidine (5-aza, a DNA demethylation agent that promotes cardiomyogenic differentiation), trichostatin A (TA, a histone deacetylase inhibitor), and activin A (AA), bone morphogenetic protein-4 (BMP-4), or after co-culture with murine fetal cardiomyocytes.
Materials and Methods
Cell preparation
hAMCs were isolated from human amnion, as previously described (Zhao et al., 2005). The cells were transfected with viral oncogenes E6 and E7, and transfected with human telomerase reverse transcriptase (hTERT). Stable transduced cells with an expanded life span were designated as immortalized hAMCs (iHAMs) (Teng et al., 2012, in press). The study and the use of the human amnion cells were approved by the Ethics Committee of the University of Toyama. Briefly, iHAMs expressed the stem cell markers such as Oct3/4, Sox2, Klf4, and SSEA-4, and showed adipogenic, osteogenic, and chondrogenic differentiation abilities. iHAMs were cultured with Dulbecco's modified Eagle medium (DMEM; Sigma, Irvine, UK) containing 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin mixture (Nacalai Tesque, Kyoto, Japan) under a humidified atmosphere of 5% CO2 at 37°C.
Construction of plasmids
The plasmid pCMV-loxP-STOP-loxP-Red-puror was constructed as follows. The AgeI–NotI fragment was excised from pIRESpuro vector (Clontech, US). The AgeI-Red-NotI fragment was excised from pDsRed-Express plasmid (Clontech, US), and was inserted into pIRESpuro vector to generate plasmid 1 (Fig. 1A). The KpnI-IRESpuror-XbaI fragment was excised from pIRESpuro vector. The KpnI-loxP-STOP-loxP-XbaI fragment was excised from Lox-STOP-Lox TOPO plasmid (Adgene Inc) and was inserted into pIRESpuro vector to create plasmid 2 (Fig. 1B). Then, plasmid 2 was linearized at the XbaI site and treated with alkaline phosphatase (Escherichia coli c75) (Takara Bio Inc). The NheI-IRESpuror-XbaI fragment was excised from plasmid 1 and inserted into the linearized plasmid 2 (Fig. 1C).
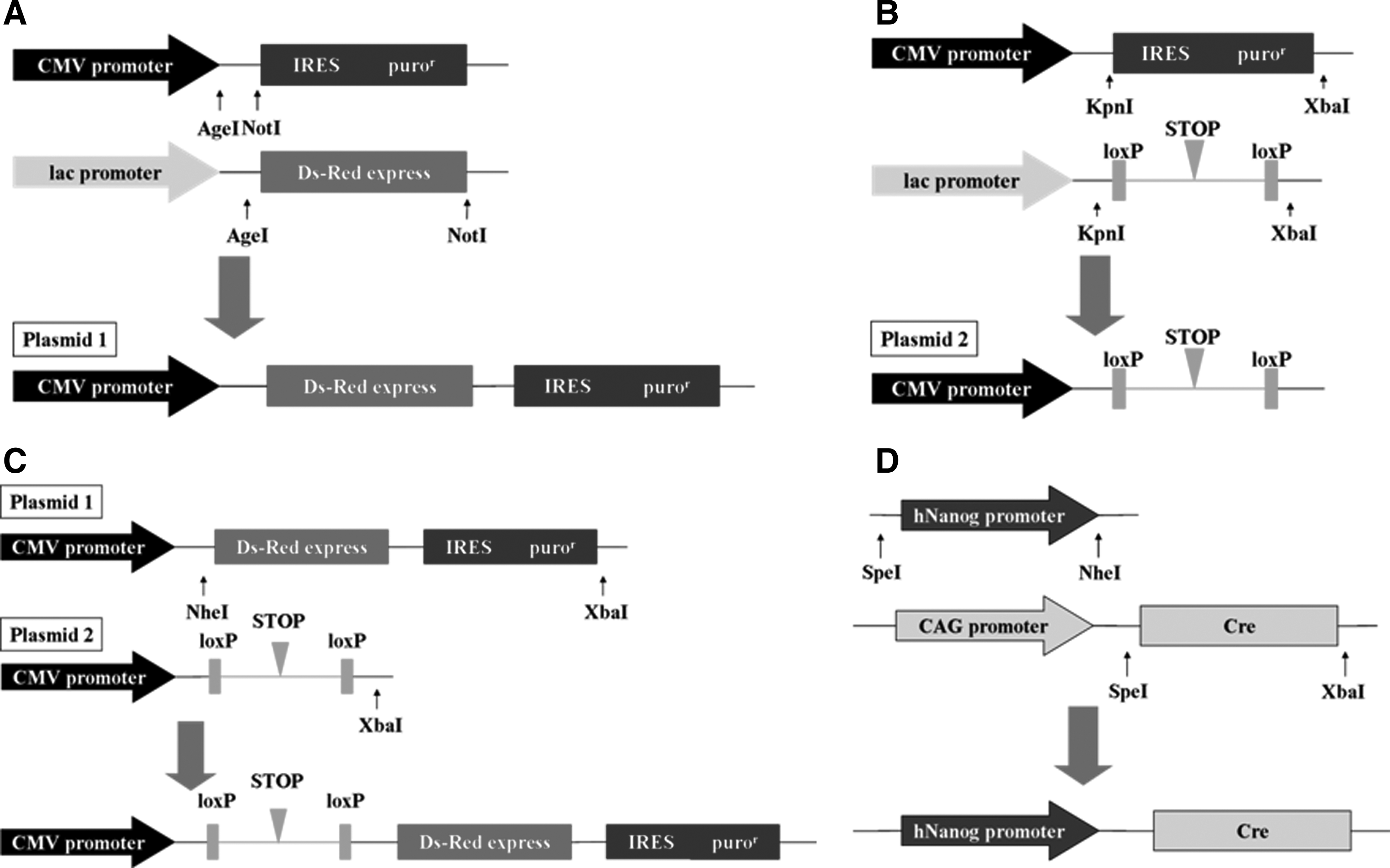
Plasmid construction. (
The plasmid human Nanog promoter-Cre was constructed as follows. The SpeI-Nanog promoter-NheI fragment was amplified from human genomic DNA (Toyobo, Osaka, Japan) using PrimeSTAR GXL DNA polymerase (Takara Bio Inc) (Kuroda et al., 2005). The SpeI–XbaI fragment was excised from pCAG-Cre vector (Adgene Inc). Then, the SpeI-human Nanog promoter-NheI fragment was inserted into the pCAG-Cre vector (Fig. 1D).
Plasmid extraction was carried out by using NucleoBond Xtra Midi Kit (Macherey and Nagel, Düren, Germany). After transformation and plasmid extraction, the concentration of the plasmid solution was adjusted to 1 μg/μL.
Plasmids transfection and selective isolation of Nanog-positive iHAMs
Plasmids transfection into iHAMs was performed by using electroporation method. One million iHAMs in 90 μL of OPTI-MEM solution (GIBCO) were mixed with 10 μg plasmid solution (loxP:Cre molar ratio is 1:5) and then transfected by using a CUY Pro-Vitro electric pulse generator (NeppaGene, Tokyo, Japan). Initially, electroporation was performed with pCAG-Cre plasmid and pCMV-loxP-STOP-loxP-Red-puror plasmid to determine the optimal conditions. The setting of poring square electric pulses were 100, 125, 150, or 175 V; initial poring square electric pulse was applied, pulse width 10 msec, 1 pulse, followed by 20 V driving square electric pulse, pulse length 10 msec, 10 pulses. The transfected iHAMs were plated onto a 10-cm dish and cultured with DMEM containing 10% FBS and 1% penicillin-streptomycin mixture under a humidified atmosphere of 5% CO2 at 37°C. After 24 h, transfected cells were cultured with DMEM containing 30 μg/mL puromycin for 48 h to isolate Nanog-positive iHAMs. In this study, Nanog-“negative” iHAMs, which did not express red fluorescence after plasmid transfection, were collected with fluorescence-activated cell sorting (FACS) method and used as control.
Cardiomyogenic induction with chemical agents
Nanog-positive iHAMs were exposed to 10 μM 5-aza (Sigma-Aldrich) (Makino et al., 1999), 10 ng/mL TA (Sigma-Aldrich) (Hosseinkhani et al., 2007), 100 ng/mL AA (Sigma-Aldrich) (Burridge et al., 2007; Yao et al., 2006), or 25 ng/mL BMP-4 (Sigma-Aldrich) (Takei et al., 2009) for 24 h. The cells were cultured on a 24-well culture dish for 28 days with DMEM containing 20% FBS and 1% penicillin-streptomycin mixture under a humidified atmosphere of 5% CO2 at 37°C.
Co-culture with murine fetal cardiomyocytes
Fetal cardiomyocytes were obtained from the hearts of day-15 mouse fetuses. Hearts were minced and washed with phosphate-buffered saline (PBS), and incubated in PBS with 0.2% collagenase II (Worthington Biochemical Corporation, Freehold, NJ) and 0.1% deoxyribonuclease I (DNase I) (Sigma-Aldrich) for 20 min at 37°C. After the addition of DMEM with 10% FBS, cells were centrifuged at 1500 rpm for 5 min. The pellet was then resuspended in 10 mL of DMEM with 10% FBS and incubated on culture dish for 1 h. The floating cardiomyocytes were collected and replated at 5×105 per cm2.
Direct co-cultivation was established by seeding Nanog-positive iHAMs treated with 5-aza for 24 h on murine fetal cardiomyocytes at 3×103 per cm2. DMEM including 20% FBS was replaced every 2 days during the 14 days of co-cultivation.
Reverse transcription-polymerase chain reaction and quantitative RT-PCR
Total RNA from iHAMs was extracted using Isogen reagent (Nippongene Co., Ltd.) and treated with DNase I (Invitrogen, Carlsbad, CA) at room temperature for 15 min. cDNAs were synthesized with RNA using ReverTra AceR qPCR RT Kit (Toyobo, Osaka, Japan). cDNAs were subjected to PCR using Taq PCR Core Kit (QIAGEN). PCR was performed as follows: Denaturation at 94°C for 5 min; 30 cycles of 94°C for 30 sec, 62°C for 30 sec (except for hyperpolarization-activated cyclic nucleotide gated channels-4 [HCN4] and inwardly rectifying potassium channels-2.1 [Kir2.1]), 72°C for 30 sec; final extension was performed at 72°C for 10 min (Yoon et al., 2006). The annealing temperature for HCN4 and Kir2.1 was 60°C (Heubach et al., 2004).
Quantitative RT-PCR was conducted with Brilliant II Fast SYBR® Green qPCR Master Mix (Stratagene). Reaction mixtures contained 10 μL of 2×SYBR Green master mix, 1 μL of cDNA, 250 nM of each primer and nuclease-free water in final volume of 20 μL. All reactions were repeated independently three times in duplicate to ensure the reproducibility of the results. Data were analyzed by using MxPro software (Stratagene). Primer sequences are shown in Table 1.
F, forward; R, reverse.
Immunofluorescent staining
Immunofluorescence assays were performed as follows: iHAMs were washed in PBS and fixed by 4% paraformaldehyde for 10 min. Cells were permeablized by 0.1% Triton-X/PBS for 15 min, and blocked with BlockingOne (Nacalai Tesque, Inc.) for 30 min. The cells were incubated with primary antibodies against Nanog (1:100), Oct3/4 (1:100), Nkx2.5 (1:100), HCN4 (1:100), Kir2.1 (1:100) (all antibodies are from Santa Cruz Biotechnology, Santa Cruz, CA, USA); cardiac troponin T (cTnT, 1:100) (AbCam, Cambridge, United Kingdom); and connexin43 (1:50) (Cell Signaling Technology) at 4°C overnight. Cells were stained with fluorescein isothiocyanate (FITC)-conjugated anti-rabbit secondary antibody (1:200), FITC-conjugated anti-mouse secondary antibody (1:200), or FITC-conjugated anti-goat secondary antibody (1:200) (all of the secondary antibodies are from Beckman Coulter, Fullerton, CA, USA) at room temperature for 1 h. Nuclear staining was performed with Hoechst 33342 (Dojindo Laboratories, Japan). All samples were visualized using fluorescence microscopy (Leica DM/RBE, Wetzlar, Germany), and figures were analyzed with a DP-70 digital microscope camera (Olympus, Japan).
Results
Selective isolation of Nanog-positive iHAMs
Survival rates of iHAMs after electroporation with pCAG-Cre plasmid and pCMV-loxP-STOP-loxP-Red-puror plasmid, which were investigated to determine the optimal conditions, were 91.1±0.96% at 100 V, 70.5±2.01% at 125 V, and 61.8±2.48% at 150 V. iHAMs electroporated at 175 V almost died. Transfection efficiencies were 55.5±2.43% at 100 V, 67.7±3.00% at 125 V, and 93.7±3.45% at 150 V. From these results, we adopted a 150 V poring square electric pulse. Transfection efficiency with pCMV-loxP-STOP-loxP-Red-puror plasmid and human Nanog promoter-Cre plasmid was 23.05±3.80%.
As shown in Figure 2, transfection with only pCMV-lox-STOP-lox Red-IRESpuror showed no red-colored cells by fluorescence microscopy at 24 h after transfection (Fig. 2a, b). When iHAMs were co-transfected with pCMV-lox-STOP-lox Red-IRESpuror and human Nanog promoter-Cre, some cells were visualized with red fluorescence (Fig. 2c, d). After 48 h of culture with DMEM containing puromycin, only Nanog-positive iHAMs survived (Fig. 2e, f).
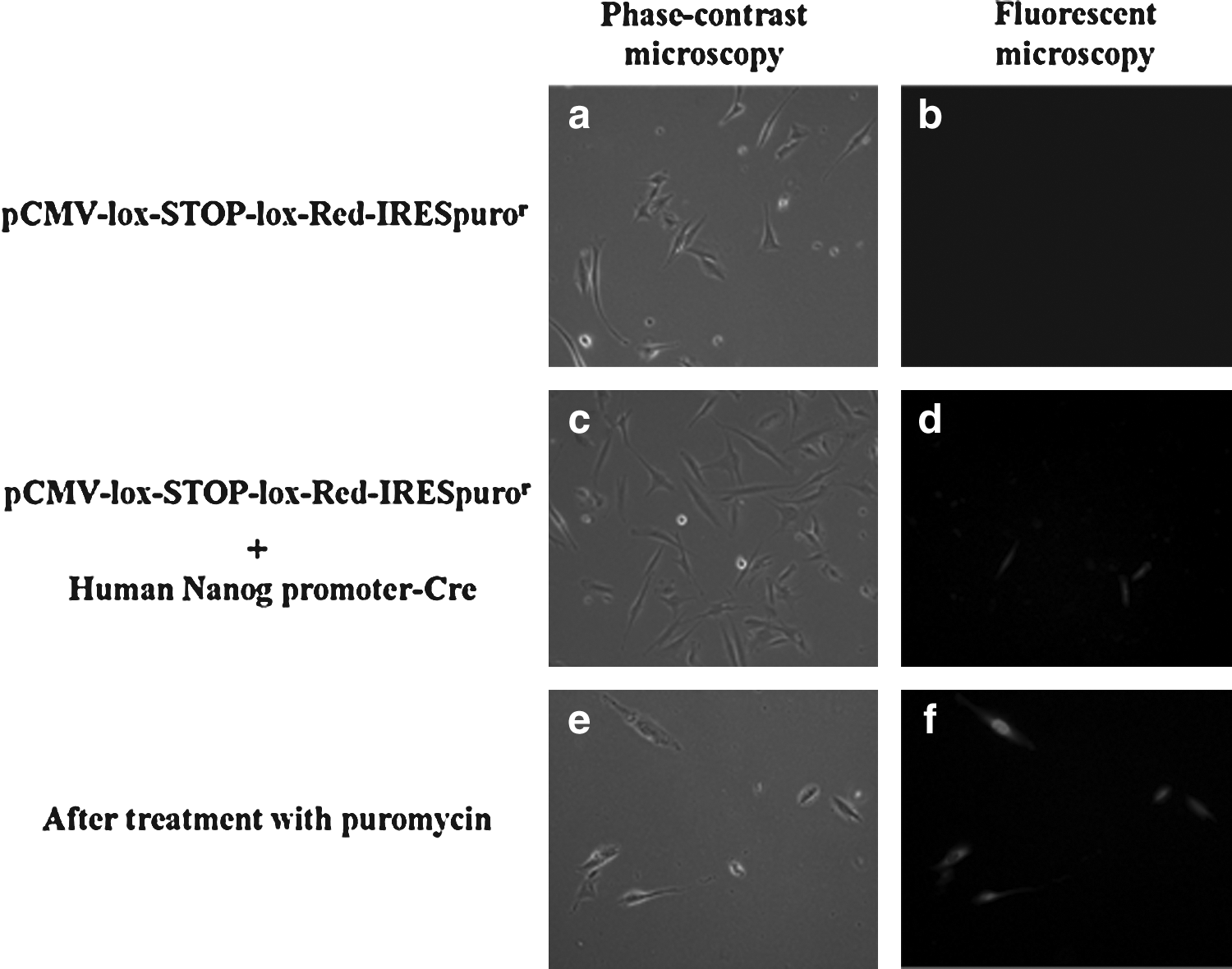
iHAMs after plasmid tranfection. (
Characteristics of Nanog-positive iHAMs
To examine the characteristics of Nanog-positive iHAMs, quantitative RT-PCR and immunofluorescent staining were performed after selection with puromycin. Quantitative RT-PCR analysis showed that the expression levels of Nanog, Oct3/4, Sox2, and Klf4 were significantly higher in Nanog-positive iHAMs than in Nanog-negative iHAMs (p<0.05; Fig. 3A). Immunofluorescent staining confirmed that Nanog-positive iHAMs expressed Nanog and Oct3/4 both in the nucleus and cytoplasm (Fig. 3B, e and k) of those cells, while they were weakly detected at the cytoplasm, but not detected at the nucleus, of Nanog-negative iHAMs (Fig. 3B, b and h).
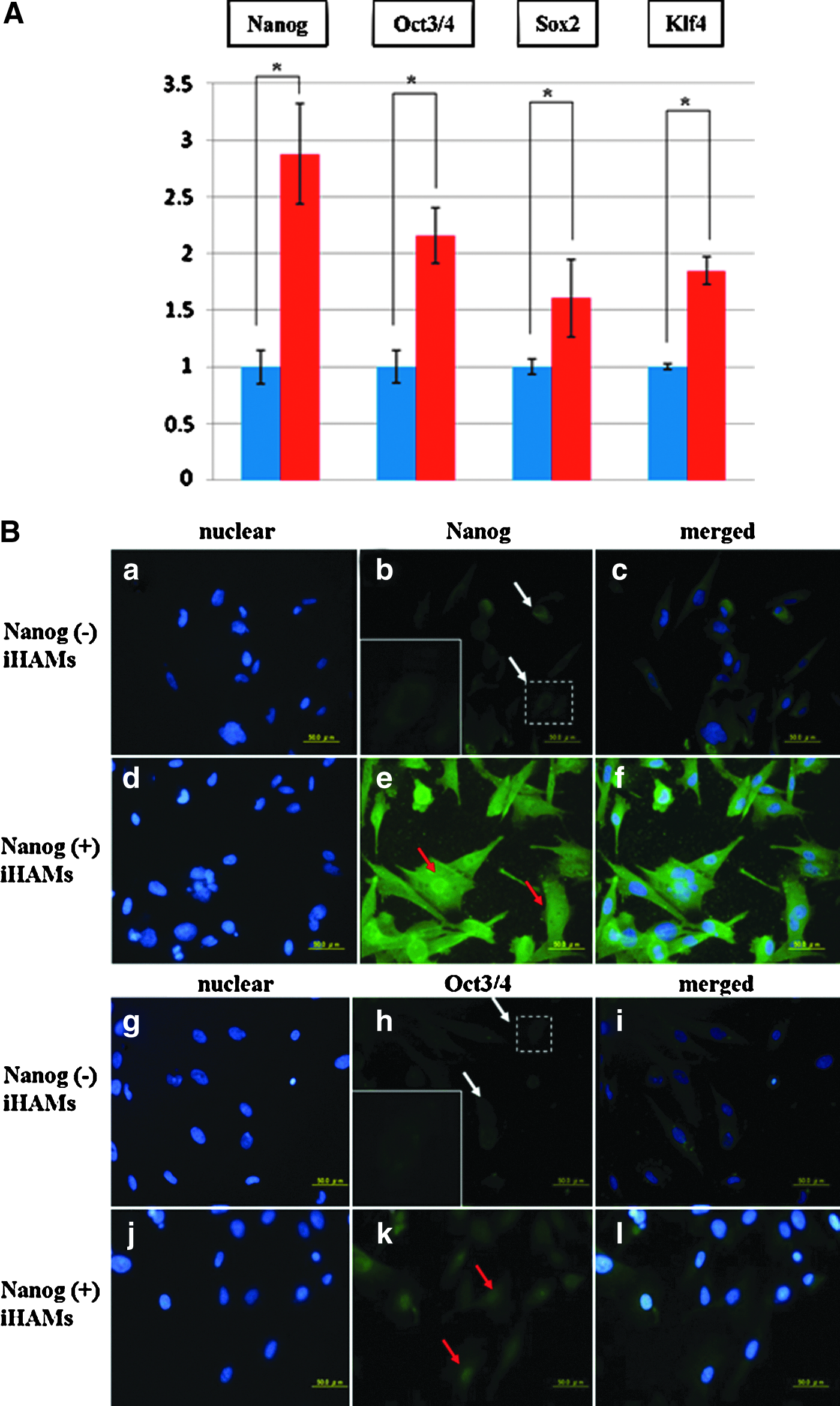
The characteristics of Nanog positive iHAMs. (
Differentiation of Nanog-positive iHAMs into cardiomyocytes by chemical agents
To characterize the cardiomyogenic differentiation potential of Nanog-positive iHAMs, RT-PCR and immunofluorescent staining were performed after 28 days of cardiomyogenic induction. RT-PCR results showed that Nanog-positive iHAMs treated with 5-aza expressed Nkx2.5 and GATA4 (myocardial precursor determination factors in early heart development). Human atrial natriuretic peptide (hANP), cTnT, myosin light chain-2a (Mlc-2a), Mlc-2v, and β-myosin heavy chain (β-MHC) were also expressed in Nanog-positive iHAMs treated with 5-aza. β-MHC and cTnT are considered important structural elements of cardiac tissue and play a role in the contraction of muscle cells. In addition, the cardiac ion channel genes HCN4 and Kir2.1 were also expressed (Fig. 4A). On the other hands, only Nkx2.5 and cTnT were expressed in Nanog-negative iHAMs treated with 5-aza.
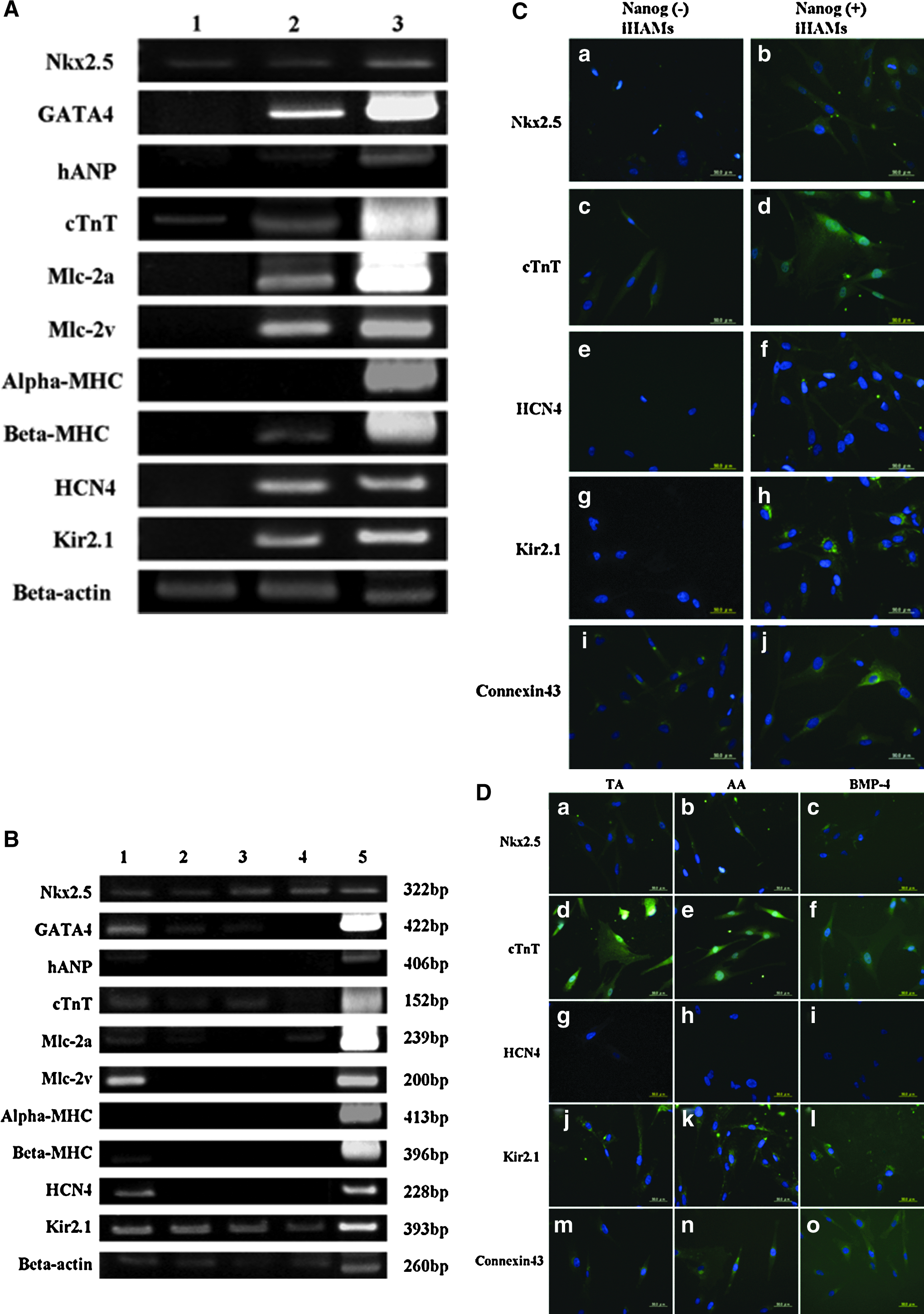
Cardiomyogenic differentiation after treatment with 5-aza, TA, AA, or BMP-4. (
Immunofluorescent staining also showed that Nanog-positive iHAMs treated with 5-aza expressed cardiac-specific proteins, such as Nkx2.5, cTnT, and cardiac ion channels HCN4 and Kir2.1 (Fig. 4C, b, d, f, and h). Furthermore, connexin43, which is the main protein of gap junctions essential for the transmission of the contraction signal, was detected in Nanog-positive iHAMs treated with 5-aza (Fig. 4C, j). On the other hand, Nkx2.5, cTnT, and connexin43 were detected, but HCN4 and Kir2.1 were not detected in Nanog-negative iHAMs treated with 5-aza (Fig. 4C, a, c, e, g, and i).
As shown in Figure 4B, Nanog-positive iHAMs treated with TA expressed Nkx2.5, GATA4, cTnT, and Kir2.1. Other genes, such as hANP, Mlc-2a, Mlc-2v, α-MHC, β-MHC, and HCN4 were not detected in Nanog-positive iHAMs with TA. Nanog-positive iHAMs treated with AA expressed Nkx2.5, GATA4, cTnT, and Kir2.1, while hANP, Mlc-2a, Mlc-2v, α-MHC, β-MHC, and HCN4 were not detected. Nanog-positive iHAMs treated with BMP-4 expressed Nkx2.5, Mlc-2a, and Kir2.1, while GATA4, cTnT, hANP, Mlc-2v, α-MHC, β-MHC, and HCN4 were not detected.
Immunofluorescent staining also showed that Nanog-positive iHAMs treated with TA, AA, or BMP-4 expressed Nkx2.5 (Fig. 4D, a–c), cTnT (Fig. 4D, d–f), Kir2.1 (Fig. 4D, j–l), and connexin43 (Fig. 4D, m–o), but did not express HCN4 (Fig. 4D, g–i). These results indicated that 5-aza was the most effective chemical agent for differentiating Nanog-positive iHAMs into cardiomyocytes.
Differentiation of Nanog-positive iHAMs into cardiomyocytes by co-culture with murine fetal caridomyocytes
From the results of cardiomyogenic induction by chemical agents, Nanog-positive iHAMs treated with 5-aza were used in co-culture experiment. After 7 days of co-cultivation, some Nanog-positive iHAMs had elongated and formed myotube-like structures. In addition, Nanog-positive iHAMs around murine fetal cardiomyocytes looked like they were beating spontaneously. RT-PCR and immunofluorescent staining were performed after 14 days of co-cultivation. To identify iHAMs in immunofluorescent staining, they were stained with human CD44-PE (Fig. 5B). RT-PCR showed that all of the analyzed genes, such as Nkx2.5, GATA4, cTnT, hANP, Mlc-2a, Mlc-2v, α-MHC, β-MHC, HCN4, and Kir2.1, were expressed in Nanog-positive iHAMs (Fig. 5A). Immunofluorescent staining revealed that Nanog-positive iHAMs were stained with Nkx2.5 (Fig. 5C, a–d), cTnT (Fig. 5C, e–h), HCN4 (Fig. 5C, i–l), Kir2.1 (Fig. 5C, m–p), and connexin43 (Fig. 5C, q–t).
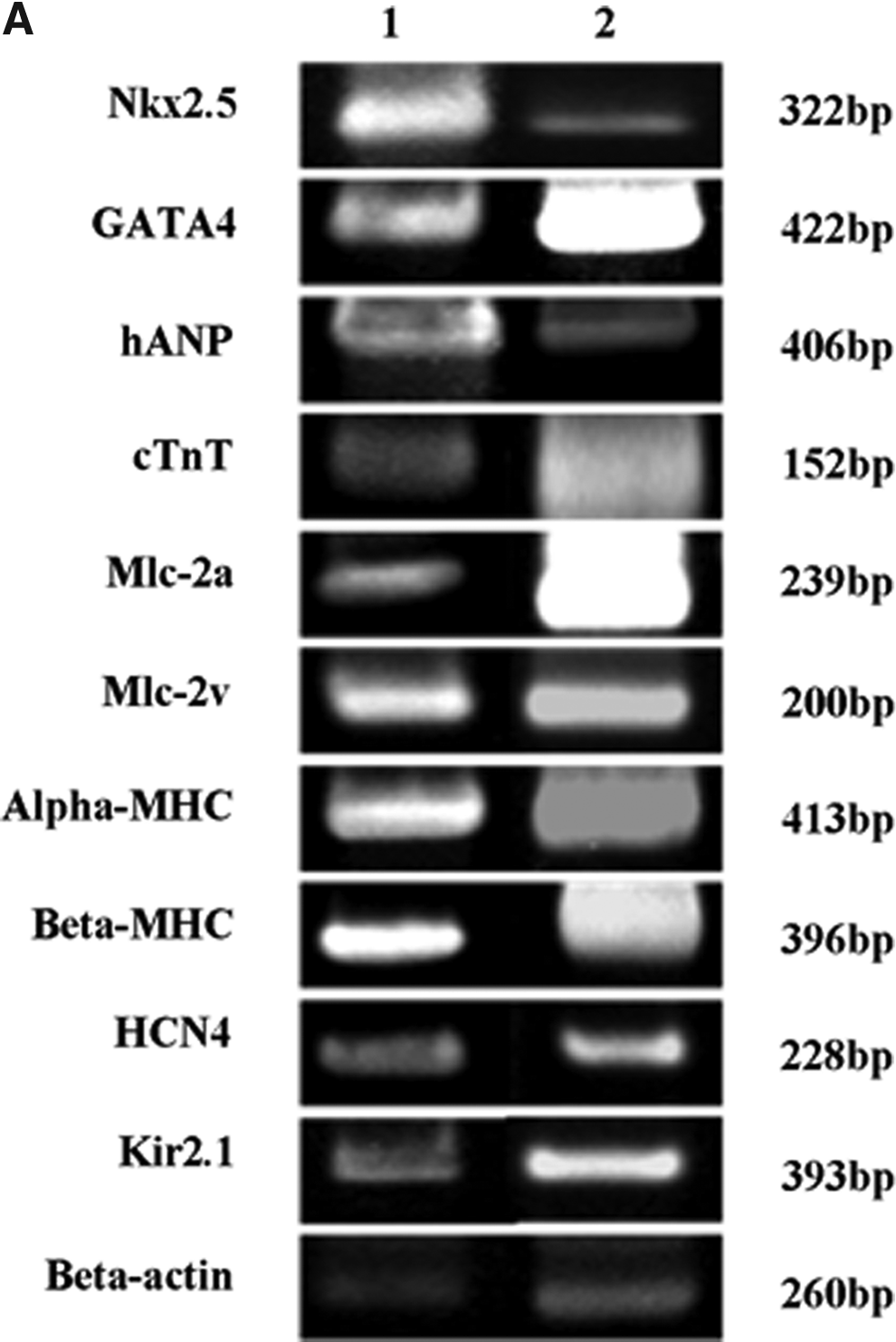
Cardiomyogenic differentiation after co-cultivation with murine fetal cardiomyocytes. (
Discussion
In this study, we demonstrated that Nanog-positive iHAMs were isolated by using the Cre/loxP system. These results suggested that Nanog-positive iHAMs were considered to be a useful cell source for cardiac regeneration therapy.
It has never been reported that hAMCs can differentiate into mature, functional cardiomyocytes. Zhao et al. reported that freshly isolated amnion-derived cells expressed cardiac genes, such as Nkx2.5, GATA4, β-MHC, and connexin43 (Zhao et al., 2005). They failed to show clear evidence of cardiomyogenic differentiation in vivo, although they used a colonial cloning technique to isolate stem cells from hAMCs. We did not get sufficient results because our previous studies had used the bulk cells from hAMCs for cardiomyogenic differentiation. These results suggest that it is necessary to purify stem cells from hAMCs with another new method to differentiate them into mature cardiomyocytes.
Nanog, Oct3/4, and Sox2 are known to be homeobox-containing transcription factors and main factors for maintaining the pluripotent embryonic stem cell phenotype (Kuroda et al., 2005; Rodda et al., 2005; Yates and Chambers, 2005). The expression of Nanog is promoted when Oct3/4 and Sox2 are sufficiently expressed and bind the Oct3/4-Sox2 enhancer region, which lies at upstream of Nanog (Kuroda et al., 2005; Rodda et al., 2005; Yates and Chambers, 2005). In addition, Nanog contributes to maintain stemness in an expression level–dependent manner. Many hAMCs expressed Oct3/4 and Sox2, but few expressed Nanog (Miki and Strom, 2006). Thus, we hypothesized that Nanog-positive hAMCs would express Oct3/4 and Sox2 sufficiently, and focused on the purification of Nanog-positive hAMCs using the new system.
In our study, we used the Cre/loxP system to isolate Nanog-positive iHAMs. This system is a site-specific recombination system that involves the targeting of a specific sequence of DNA and splicing it (Ray et al., 2000; Yoshimura et al., 2001). We expected that Cre/loxP system would allow us to purify Nanog-positive iHAMs. In fact, Nanog-positive iHAMs could be visualized under the fluorescence microscope and isolated from iHAMs after treatment with puromycin (Fig. 2). Nanog-positive iHAMs existed as 2–2.5×105 cells in initial 1×106 cells. 1–3×107 hAMCs are able to be obtained from an amnion. Thus, it will be expected that 2–2.5×106 Nanog-positive hAMCs are able to be obtained at one parturition.
Yamanaka et al. established iPSCs from mouse somatic cells by introducing four factors, such as Oct3/4, Sox2, Klf4, and c-myc (Takahashi et al., 2007). They also reported that Nanog-iPSCs, which were selected by Nanog expression, showed morphology and proliferation similar to those of ESCs and expressed ESC markers, such as Nanog, Fbx15, ERas, and ESG1, higher than their “original” iPSCs (Okita et al., 2007). They confirmed that Nanog, Oct3/4, and Sox2 play an essential role in maintaining undifferentiated state and pluripotency. Our present studies for the characteristics of Nanog-positive iHAMs showed that the expression levels of Nanog, Oct3/4, Sox2, and Klf4 were significantly higher in Nanog-positive iHAMs than in Nanog-negative iHAMs (Fig. 3A). In addition, immunofluorescence analysis showed that Nanog-positive iHAMs expressed Nanog and Oct3/4 stronger at nucleus than cytoplasm (Fig. 3B). In some previous studies with ESCs, Nanog and Oct3/4 expression was detected, especially in nucleus (Hatano et al., 2005; Yamaguchi et al., 2005). These results suggest that Nanog-positive iHAMs are a group of undifferentiated stem cells and would have pluripotency.
In this study, we examined which chemical agents were most effective for differentiating iHAMs into cardiomyocytes. 5-aza was the most popular chemical agent to induce cardiomyogenic differentiation. Many research groups have reported that 5-aza induced ESCs, iPSCs, and mesenchymal cells to differentiate into cardiomyocytes (Kehat et al., 2001; Li et al., 2008; Mummery et al., 2003; Shim et al., 2004; Tanaka et al., 2009; Zwi et al., 2009). Hosseinkhani et al. reported that TA promotes the differentiation of cynomolgus monkey ESCs into cardiomyocytes (Hosseinkhani et al., 2007). Burridge et al. showed that AA increased beating embryoid bodies 13-fold relative to mass culture (Burridge et al., 2007). Another chemical agent, BMP-4, was reported by Sachinidis et al. Their results showed that BMP-4 induced the expression of Nkx2.5 in cardiac progenitor cells (Sachinidis et al., 2006; Takei et al., 2009). Our results showed that all of these chemical agents promoted cardiomyogenic differentiation of Nanog-positive iHAMs at varying degrees of efficiency. However, Nanog-positive iHAMs treated with 5-aza expressed the largest number of cardiac specific genes and cardiac proteins (Fig. 4B and D). 5-aza was the most effective chemical agent for cardiomyogenic induction of Nanog-positive iHAMs in this study.
Nanog-positive iHAMs differentiated into cardiomyocyte-like cells after treatment with 5-aza. RT-PCR results showed that the expression of cardiac marker genes, such as Nkx2.5, GATA4, hANP, Mlc-2a, Mlc-2v, cTnT, β-MHC, and cardiac ion channel genes HCN4 and Kir2.1 were observed in Nanog-positive iHAMs (Fig. 4A).
Immunofluorescent staining showed that Nkx2.5, cTnT, HCN4, Kir2.1, and connexin43 were detected in Nanog-positive iHAMs (Fig. 4C). On the other hand, Nanog-negative iHAMs treated with 5-aza expressed Nkx2.5, cTnT, and connexin43, but they did not express other cardiac genes and proteins. These results suggested that Nanog-positive iHAMs have much more potential for differentiation into cardiomyocytes than Nanog-negative iHAMs. Expression of Nkx2.5 was also observed in Nanog-negative iHAMs treated with 5-aza, which indirectly attests to the differentiation potential of hAMCs.
In the present study, Nanog-positive iHAMs treated with 5-aza could differentiate into cardiomyocyte-like cells, but no beating cells were observed. One reason for this situation might be attributed to the fact that the concentration of 5-aza was not adequate or there is a variety of additional factors necessary to promote the differentiation of iHAMs to mature cardiomyocytes. Makino et al. first reported that mouse bone marrow MSCs differentiated into beating cardiomyocytes after treatment with 3 μM 5-aza (Makino et al., 1999). Some authors reported that the best concentrations for cardiac differentiation of human MSCs were between 5 and 10 μM (Cao et al., 2004; Xu et al., 2004). However, for cardiac differentiation of human ESCs, Yoon et al. reported that the concentration 0.1 μM was best (Yoon et al., 2006). Therefore, it appears that the concentration of 5-aza was critical in the differentiation into cardiomyocytes. Another reason might be that we used immortalized cells. Cardiomyogenic potential of hAMCs may be limited due to the transfection with viral oncogenes E6 and E7, and hTERT.
According to the results of Nishiyama et al., human cord blood MSCs could differentiate into cardiomyocytes when co-cultured with murine fetal cardiomyocytes (Nishiyama et al., 2007). They suggested that there might be some factors supplied by the cardiac microenvironment. After co-culturing with the murine fetal cardiomyocytes, we showed that Nanog-positive iHAMs looked like they were beating in a synchronized manner, though the expression of Nkx2.5, GATA4, hANP, cTnT, Mlc-2a, Mlc-2v, α-MHC, β-MHC, HCN4, and Kir2.1 (Fig. 5A and C). Our results suggested that cardiac microenvironment might be essential for obtaining beating functional cardiomyocytes because chemical agents alone might not be enough to differentiate Nanog-positive iHAMs into beating cardiomyocytes. Subsequent research will be required to differentiate iHAMs into beating cardiomyocytes.
In conclusion, we could isolate Nanog-positive hAMCs by using the Cre/loxP system. This isolation method would be useful because of its simplicity and certainty. Nanog-positive iHAMs had characteristics of stem cells and expressed many cardiac markers after treatment with 10 μM 5-aza, and few markers after treatment with TA, AA, or BMP-4. Beating cardiomyocytes were observed after co-cultivation with murine fetal cardiomyocytes. Nanog-positive iHAMs were considered to have the potential to be a useful cell source for cardiac regeneration therapy. Further studies will be necessary to obtain functional cardiomyocytes derived from Nanog-positive hAMCs.
Footnotes
Acknowledgments
We would like to thank Mrs. Furuichi for providing expert technical assistance with this study.
Author Disclosure Statement
The authors declare that they have no conflicts of interest.