
Abstract
Abstract
Bovine embryonic stem cells (ESCs) are a powerful tool for agricultural and biomedical applications. The purpose of this study was to introduce a new method for generating bovine ESCs. Mechanically isolated bovine inner cell masses (ICMs) from in vitro–produced blastocysts were cultured individually on a 10-μL mouse embryonic fibroblast (MEF) feeder cell drop covered with oil. From 126 blastocysts classified by their developmental stage and ICM size, 21 primary bovine ESC-like colonies were formed (16.7%) and established six JNU (Jeju National University)-ibES cell lines (28.6%, 6/21; hatched blastocyst×4, hatching blastocyst×1, and expanded blastocyst×1). These cells exhibited typical ESC morphology, and pluripotency markers were detected through immunocytochemistry, RT-PCR, and real-time RT-PCR, including Oct4, stage-specific embryonic antigen-1 (SSEA-1), Nanog, Tumor rejection antigen-1-81, Rex1, and alkaline phosphatase. Through RT-PCR analysis of spontaneous differentiation, gene expression of all three embryonic germ layers was detected: ectodermal (Pax6 and DBH), mesodermal (CMP and Enolase), and endodermal [alpha fetoprotein (α-FP) and albumin]. In addition, JNU-ibES cell lines were directed differentiated into neuronal (Map2 and Tuj1) and glial (GFAP) cells. Bovine ESC lines had a normal karyotype, with a chromosome count of 58+XY (JNU-ibES-05). This is the first trial investigating a minimized microdrop culture method for the generation of bovine ESCs. These results demonstrated that the minimized MEF feeder cell drop can support the establishment of bovine ESC lines.
Introduction
However, ESC lines from domestic animals have been relatively less studied than those of murine ESC due to several challenges, such as longer generation time and suboptimal culture condition (Mann and Stewart, 1991; Wilmut et al., 1991). The derivation and maintenance of ESC lines from bovine blastocysts is perhaps more difficult and challenging than that of all other mammalian blastocysts. To date, there were some reports of preventing spontaneous differentiation of bovine ESCs, such as ICM isolation or subculture technique, feeder cell type, ESC culture medium, and growth factors (Cao et al., 2009; Gong et al., 2010; Mitalipova et al., 2001; Verma et al., 2007; Wang et al., 2005). Also, there have been conflicting or variable results for pluripotent marker expression in bovine ESCs (Cao et al., 2009; Gong et al., 2010; Mitalipova et al., 2001; Wang et al., 2005). In addition, although there have been several reports on the derivation of bovine ESC-like cells, stable bovine ESC lines are not yet available (Keefer et al., 2007; Talbot and Blomberg, 2008).
In this study, we developed a novel strategy—a minimized feeder cell drop—to enhance the generation rate and extend the maintenance term of bovine ESCs developed from in vitro-produced 9-day-old bovine blastocysts that had been classified by developmental stage and ICM size. We then attempted to determine typical bovine ESC morphology, ESC marker expression, and pluripotency characteristics under spontaneous or directed neuronal cell differentiation conditions in vitro.
Materials and Methods
Unless stated otherwise, all chemicals were purchased from Sigma Chemical Company (St Louis, MO, USA).
Production of bovine in vitro–matured, in vitro–fertilized, and in vitro–cultured embryos
Bovine ovaries were collected from a slaughterhouse and transported to the laboratory within 2 h in 0.9% saline at 35°C. Cumulus–oocyte complexes (COCs) were aspirated from visible follicles (2–6 mm in size) with an 18-gauge needle attached to a 10-mL disposable syringe. The medium used for COC collection was HEPES-buffered Tyrode's medium (TL-HEPES). Sets of 10 COCs were matured in vitro in a 50-μL drop of tissue culture medium-199 (TCM-199; Gibco, USA) supplemented with 10% fetal bovine serum (FBS), 0.2 mM sodium pyruvate, 1 μg/mL follicle-stimulating hormone (Folltropin™, Bioniche Animal Health, Canada), 1 μg/mL estradiol-17β, and 1 mM epidermal growth factor (EGF) in mineral oil at 38.8°C in an incubator (5% CO2, 5% O2, and 90% N2). After incubation for 22–24 h in the in vitro maturation (IVM) medium, the COCs were used for in vitro fertilization (IVF) using highly motile sperm recovered from frozen–thawed bull semen, which had been separated on a discontinuous Percoll (GE Healthcare, Bio-Sciences AB, Uppsala) column. Matured COCs were transferred to 44-μL drops (10 COCs per drop) of fertilization medium containing 6 mg/mL fatty acid-free bovine serum albumin (FAF-BSA), 0.2 mM pyruvate, and 25 μg/mL gentamicin. Sperm were counted using a hemocytometer and diluted at a concentration of 1×106 cells/mL. Two microliters each of sperm, heparin (2 μg/mL), and phenylalanine (PHE) containing 18.2 M penicillamine, 9.1 M hypotaurine, and 1.8 M epinephrine were added to the 44-μL IVF drop. Fertilization was assessed by the cleavage rate (≥two-cell) after 44±2 h of incubation. For in vitro culture (IVC), cleaved embryos were incubated in CR1aa medium containing 3 mg/mL of fatty acid-free bovine serum albumin (FAF-BSA) until embryonic day 4 and then they were transferred into CR1aa medium containing 10% FBS until embryonic day 9.
Preparation of mouse embryonic fibroblastic feeder cells
For mouse embryonic fibroblast (MEF) cell preparation, uteri isolated from 13.5-day pregnant CDF1 mice were washed with phosphate-buffered saline (PBS). The head and visceral tissues were removed from isolated embryos, and the bodies were washed with fresh PBS, minced using a pair of scissors, transferred into a solution of 0.25 mM trypsin/1 mM EDTA, and incubated at 37°C in a shaking incubator for 30 min. After trypsinization, the cells were dissociated by pipetting up and down, and were then collected by centrifugation at 100×g (1000 rpm) for 1 min. The pelleted cells were washed twice in culture medium, and the resuspended cells were cultured on 100-mm dishes (≥1×106) in a 5% CO2 incubator at 37.0°C in Dulbecco's modified Eagle medium (DMEM; Gibco) containing 10% defined FBS (Hyclone, USA), 50 units/mL of penicillin, and 50 μg/mL of streptomycin. The third to the fifth passages of cultured and 10 μg/mL mitomycin C–treated MEF cells were used as the feeder cells, and they were seeded in a two-well dish with several 10-μL drops at a concentration of 1.6×105 cells/mL, and then covered with mineral oil.
Derivation and culture of IVF bovine ESCs
The ICMs of bovine IVM/IVF/IVC blastocysts were isolated mechanically using 28-gauge needles attached to 0.5-mL syringes (BD, USA). Isolated ICMs were classified by embryo development stage and size (small, medium, and large) as a parameter for the experiment of IVF-bovine ESCs (ibESCs) establishment. ICM cells were placed onto each MEF feeder cell drop and cultured in DMEM (Gibco) containing 20% FBS, 0.55 mM β-mercaptoethanol (Gibco), 1% nonessential amino acids (Sigma), 4 ng/mL basic fibroblast growth factor (bFGF; KOMA Biotech. Inc., Korea), 1 ng/mL insulin-like growth factor (IGF; Bio-Research Product Inc., USA), 10 ng/mL EGF, and 500 ng/mL human noggin protein (PeproTech Inc., USA). The medium was changed every 2 days. After 5–8 days, the ICM-derived outgrowths were separated by mechanical dissociation and replated on a new mitomycin C–treated MEF feeder cell drop. To passage the putative ESCs, individual colonies with a uniform and undifferentiated morphology (exhibiting a small cytoplasmic/nuclear volume ratio and forming tight, rounded, and multilayered clumps) were selected using a micropipette, mechanically dissociated into two to three clumps, and then placed on an MEF feeder cell drop. The established ibESCs were passaged every 7–10 days after replating.
Alkaline phosphatase staining and immunocytochemistry
To detect alkaline phosphatase (AP) activity, ibESC colonies were fixed in 4% paraformaldehyde (PFA) in 0.1 M PBS for 2 min and then treated with Fast Red Violet/Naphthol AS-BI phosphate mixed solution for 15 min (Millipore, USA). For immunocytochemistry, ibESC colonies or in vitro–differentiated cells were fixed in 4% PFA for 20 min at 4°C and then permeabilized with 0.2% Triton X-100 for 10 min at room temperature. To block any nonspecific binding, the cells were incubated with 10% normal goat serum (VECTOR, USA) for 1 h and then incubated with primary antibody at 4.0°C overnight. The primary antibodies used were anti-stage-specific embryonic antigen 1 (SSEA-1, Santa Cruz, USA, 1:20), anti-SSEA-4 (Santa Cruz, 1:20), anti-tumor rejection antigen (TRA) 1-81 (Santa Cruz, 1:20), anti-Nanog (Santa Cruz, 1:20), anti-Oct-4 (Santa Cruz, 1:250), anti-Map2 (Chemicon, 1:1000), anti-Tuj1 (Chemicon, 1:1000), and anti-glial fibrillary acidic protein (GFAP; Chemicon, 1:1000). The secondary antibodies used were Alexa Fluor 488 conjugated goat anti-mouse immunoglobulin G (IgG) or IgM (for SSEA-1, SSEA-4, TRA1-81, and Oct-4), Alexa Fluor 594–conjugated goat anti-rabbit IgG (for Nanog), goat anti-mouse tetramethylrhodamine isothiocyanate (TRITC; Jackson Laboratories, USA; for Tuj1), or goat anti-rabbit TRITC (for Map2 and GFAP) at a dilution of 1:200. Nuclei were stained with 5 μg/ml of 4′-6-diamidino-2-phenylindole (DAPI). Cells were observed and photographed with an inverted Olympus IX-71 (Japan) microscope equipped for epifluorescence.
mRNA Extraction and RT-PCR
For the reverse transcription PCR analysis, six ibESC lines were examined for bovine ESC marker expression and two ibESC lines were selected for in vitro differentiation ability. Total RNA was extracted from several colonies using Dynabeads mRNA DIRECT Micro Kit (Dynabeads mRNA purification kit; Dynal, Norway) according to the manufacturer's instructions. Briefly, ESC colonies and differentiation cells were suspended in 100 μL of lysis/binding buffer [100 mM Tris-HCl, pH 7.5, 500 mM LiCl, 10 mM EDTA with pH 8.0, 0.1% lithium dodecyl sulfate (LiDS), 5 mM dithiothreitol (DTT)] and vortexed at room temperature for 5 min to lyse the cells. A 50-μL aliquot of an oligo(dT)25 magnetic bead suspension was added to the samples, and the samples were incubated at room temperature for 5 min. The hybridized mRNA and oligo(dT) beads were washed twice with washing buffer A (10 mM Tris-HCl, pH 7.5, 0.15 M LiCl, 1 mM EDTA, 1% LiDS) and once with washing buffer B (10 mM Tris-HCl, pH 7.5, 0.15 M LiCl, 1 mM EDTA). mRNA samples were eluted from beads in 15 μL of double-distilled diethyl pyrocarbonate (DEPC)-treated water. cDNA was synthesized from about 1 μg total RNA using SuperScript II reverse transcriptase (Invitrogen). cDNA samples were subjected to PCR amplification with selective primers described in Table 1. The PCR products were size-fractionated by 1% agarose gel electrophoresis and visualized by ethidium bromide staining. The final analysis was conducted in an image analyzer (BioRad).
ESC, embryonic stem cell.
Real-time RT-PCR quantification
Comparative real-time PCR was performed using a Chromo 4 (Bio-Rad) and DyNAmo HS SYBR Green qPCR kit (FINNZYMES, Finland), according to the manufacturers' instructions, with the primers described in Table 1. mRNA for real-time RT-PCR was prepared from six Jeju National University (JNU)-ibESCs using magnetic beads (Dynabeads mRNA purification kit). Gene expression was quantified by the 2−ddCt method (Livak et al., 2001).
In vitro differentiation of ibESCs
For spontaneous differentiation, ibESC colonies recovered from a clump state were suspension-cultured in feeder-free conditions in ESC culture medium without bFGF and Noggin for 2 weeks. Embryoid bodies (EBs) were then plated on 0.1% gelatin-coated culture dishes and cultured in the same medium for an additional 7 days. During differentiation, the medium was replaced every 2 days. For directed neuronal cell differentiation, day-7 EBs were dissociated with TrypLE (Invitrogen, USA) solution, and single cells or clumps were cultured onto 0.1% gelatin-coated, two-well plates in N2 medium that consisted of DMEM/F12 supplemented with 0.01% BSA, 20 nM progesterone, 100 μM putrescine, 25 μg/mL insulin, 50 μg/mL transferrin, and 30 nM sodium selenite. Immunocytochemistry was performed after an additional 8 days of culture.
Karyotyping of ibESCs
After treatment in 5% Colcemid (Gibco) for 4 h, ibESCs were dissociated using TrypLE solution, hydrated in hypotonic KCl for 15 min, and fixed in acetic acid (BDH, UK) at a ratio of 3:1. The fixed cells were dropped onto clean slides at room temperature. The slides were stained with a freshly made staining solution containing 3 mL of Leishman stain in 17 mL of Gurr buffer (Invitrogen) for 8 min. A coverslip was mounted on the slides with Histomount (National Diagnostics, USA), and slides were viewed using a light microscope under oil immersion optics (Nicon C1) at 1000×magnification.
Statistical analysis
Differences in the pluripotency marker gene expression levels among JNU-ibES cell lines were examined. Differences were evaluated by analyses of variance (ANOVA) with the general linear model (PROC-GLM) in the SAS software program. Differences of p<0.05 were considered significant.
Results
Establishment of ibESC line
To establish bovine ESCs, 126 blastocysts, including hatched (n=43), hatching (n=41), or fully expanded (n=42) blastocysts, were produced at day 9 post-IVF (Fig. 1A). ICM cells were recovered mechanically. The isolated ICM cells were classified by size as small (40–90 μm, n=15), medium (90–140 μm, n=56), or large (140–190 μm, n=55) (Fig. 1B–D) using an eyepiece micrometer. Individual ICM cells were plated separately on a 10-μL-sized and mitomycin C–treated MEF feeder cell drop. Primary ESC-like colonies appeared at 4–7 days after plating (Fig. 1E). The bovine ESCs had a high nucleus-to-cytoplasm ratio and formed domed colonies with a clear borderline. At about 7–10 days, densely packed bovine ESC-like colonies were subcultured onto a new MEF feeder cell drop after they were mechanically dissected into several small clumps using needles (Fig. 1F). As shown in Table 2, the rates of bovine ESC-like colonies derived from expanded, hatching, and hatched blastocysts were 11.9% (5/42), 14.6% (6/41), and 23.3% (10/43), respectively. Also, the rates of deriving ESC-like colonies from ICMs of the size categories small, medium, and large were 13.3% (2/15), 19.6% (11/56), and 14.5% (8/55), respectively. In total, 21 bovine ESC-like colonies were obtained. From these ESC-like colonies, six bovine ESC lines were established (in total, 6/21, or 28.6%; from expanded blastocysts, 1/5, or 20.0%; from hatching blastocysts, 1/6, or 16.7%; from hatched blastocysts, 4/10, or 40.0%). These bovine ESC lines were maintained for more than 12 passages (>90 days) and termed JNU-ibES-01∼06.

Micrographs documenting the course of establishment of bovine ESCs lines from IVF embryos: morphology of the cells and colonies. (
ESC, embryonic stem cell; ICM, inner cell mass.
Characterization of bovine ESC lines
In characterizing the bovine ESC colonies, we confirmed that the pluripotency markers AP (Fig. 2A), Oct4 (Fig. 2B), SSEA-1 (Fig. 2C), Nanog (Fig. 2D), and TRA-1-81 (Fig. 2F) were all highly expressed, whereas SSEA-4 (Fig. 2E) was not. Also, RT-PCR (Fig. 3A) analysis revealed that four stem cell marker genes (Oct4, SSEA-1, ALP, and Rex1) were expressed in all JNU-ibES cell lines. However, there were differences in the relative mRNA expression levels of these four stem cell marker genes among the different JNU-ibES cell lines, as shown in Figure 3B (p<0.05). For Oct4 and ALP expression, the levels of JNU-ibES-02–06 were significantly higher than those of JNU-ibES-01; for Rex1 expression, the levels of JNU-ibES-03–06 were higher than those of JNU-ibES-01–02; and, finally, for SSEA 1 expression, the levels of JNU-ibES-03, -05, and -06 were higher than those of JNU-ibES-01, -02, and -04.
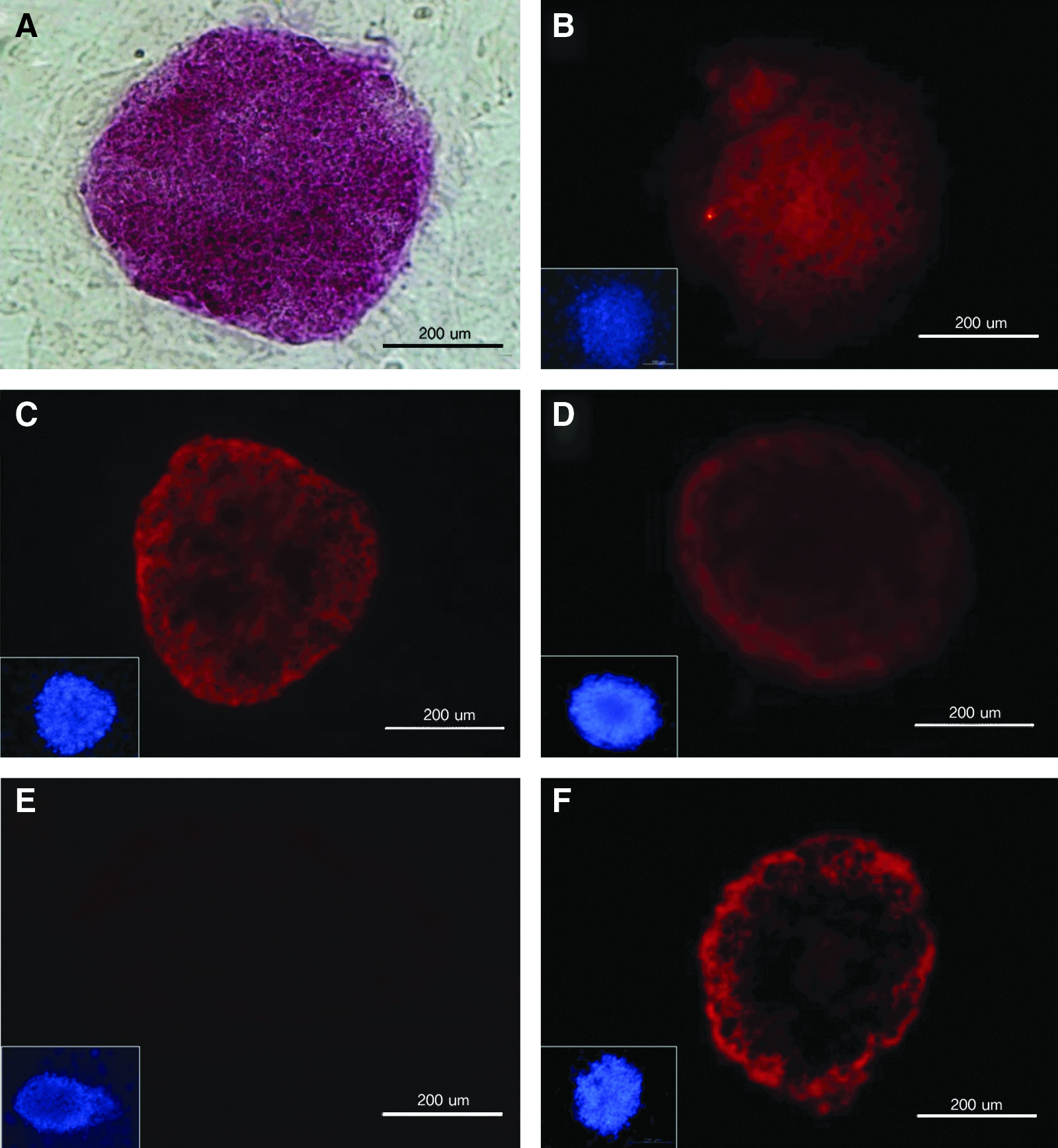
Characterization of pluripotency marker expression in bovine ESC colonies. Alkaline phosphatase
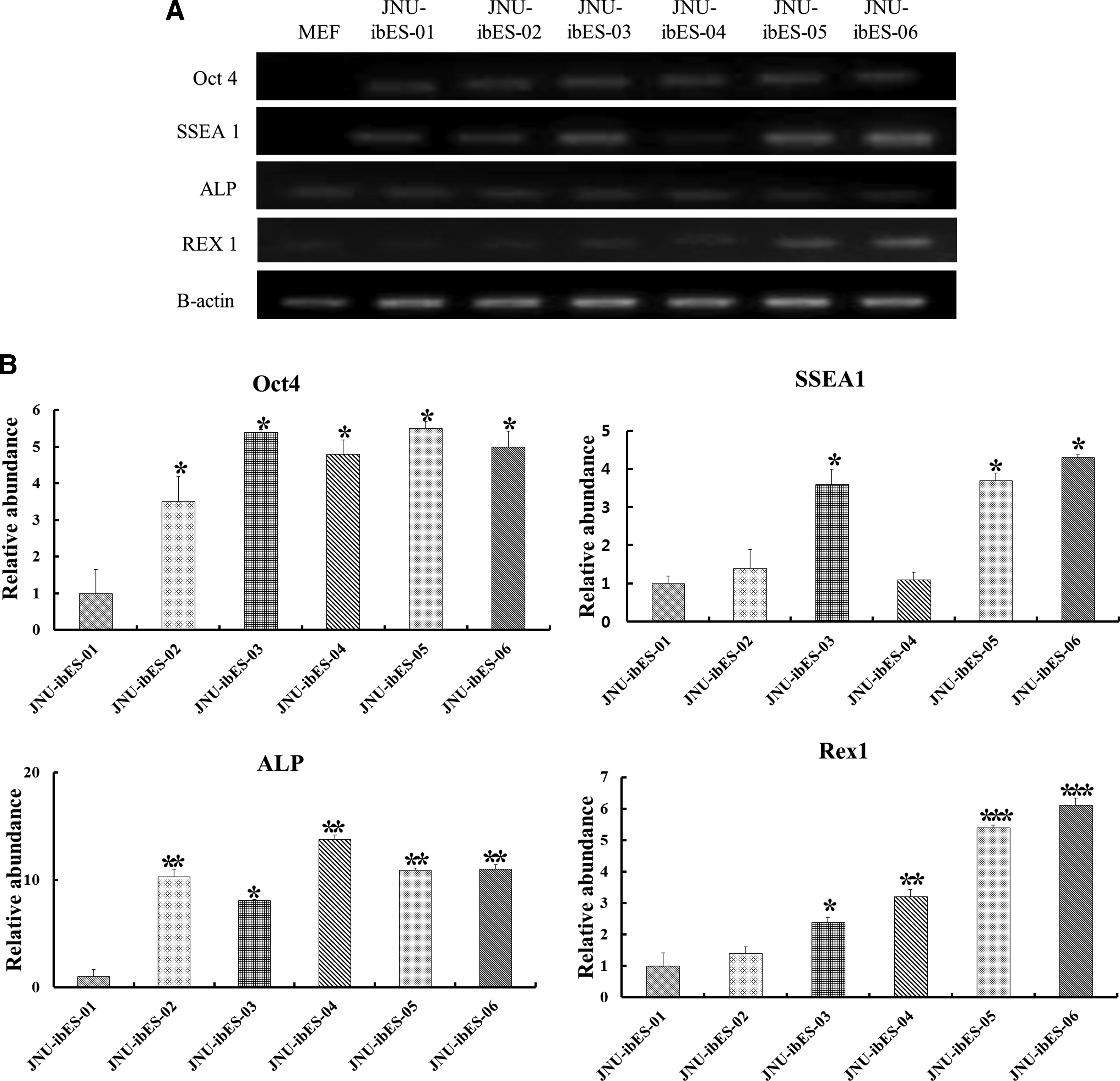
Gene expression analysis of bovine pluripotent ESC markers (Oct 4, SSEA 1, ALP, and Rex1) for six JNU-ibES cell lines by RT-PCR
In vitro differentiation of the bovine ESC lines
To determine pluripotency characteristics, spontaneous and directed in vitro differentiation of bovine ESCs were examined. In spontaneous differentiation, EBs (Fig. 4A) cultured in 0.1% gelatin-coated culture plates for 2 weeks were differentiated into various types of cells (Fig. 4B). RT-PCR analysis of spontaneously differentiated JNU-ibES-05 and JNU-ibES-06 cells confirmed expression of marker genes for all three germ layers (ectoderm, Pax6 and DβH; mesoderm, CMP and Enolase; endoderm, α-FP and albumin), whereas none of these genes were expressed in the undifferentiated ibES cells (Fig. 4C). Also, when the directed differentiation ability of bovine ESCs (JNU-ibES-05) into neuronal cells (Fig. 4D) was examined, microtubule-associated protein (MAP) 2 (Fig. 4E), neuron-specific class III β-tubulin (Tuj1) (Fig. 4F), and GFAP (Fig. 4G) expression was detected by immunocytochemistry.

In vitro differentiation characteristics of bovine ESCs. (
Karyotyping of bovine ESCs lines
JNU-ibES-05 cells were confirmed as having a normal karyotype, with a chromosome count of 58+XY (Fig. 5).

Karyotyping analysis of JNU-ibES-05 cells showed a normal XY line (58+XY).
Discussion
This study demonstrates that bovine ESC lines can be generated efficiently using a novel strategy that includes minimized MEF feeder cell drop and oil cover, from in vitro–produced day-9 hatched blastocysts. These ibESCs present typical ESC morphology, ESC marker expression, and in vitro pluripotency characteristics. The established six JNU-ibES cells showed clear formation of colonies with distinct boundaries and were maintained with more proliferative potential (>3 months) in a 10-μL MEF cell drop culture than those in other studies using large-scale culture methods (Gong et al., 2010; Pashaiasl et al., 2010; Sritanaudomchai et al., 2007). This is the first trial of a minimized microdrop culture method for the generation of bovine ESCs.
Until now, there has been only one report of successful production of unlimited expanding cultures of bovine ESCs (Mitalipova et al., 2001). The major reason for the limited proliferative potential of bovine ESCs may be the absence of an optimized feeder cell environment. In general, MEF or STO cells have been commonly used as feeder cells. MEF and STO cells have been known to function in the establishment and maintenance of mammalian pluripotent cells, including mouse ESCs, human embryonic germ cells, and hESCs (Park et al., 2004; Shi et al., 2006; Turnpenny et al., 2003). These feeder cells express various cytokines [leukemia inhibitory factor (LIF), interleukin (IL), tumor necrosis factor-α (TNF-α)], growth factors [insulin-like growth factor binding protein (IGFBP), FGF, vascular endothelial growth factor (VEGF), hepatocyte growth factor (HGF)], and extracellular matrix (ECM) materials (laminin, collagen, and fibronectin) (Park et al., 2003; Talbot et al., 2012). Most normal cells require adhesion to the ECM for survival, migration, and growth, and the high concentration of ECM materials is very helpful for ESC growth (colony type). However, those two mouse feeder cell lines may be unsuitable for the bovine ESC culture environment, considering the difference in body temperature between the two species (37.0°C vs. 39.0°C). Overly warm (≥2°C) bovine cell culture conditions may negatively affect the bioactivity of the mouse feeder cells. Therefore, the development of a more appropriate bovine feeder cell line is needed for the stable culture of ibESCs. Until now, many of the advances in bovine ESC cell generation were accomplished using mouse feeder cells, and the efficiency was very low (0.5–8.0%) (Cao et al., 2009; Mitalipova et al., 2001; Wang et al., 2005).
In this study, we developed a minimized MEF feeder cell culture method that allowed the generation rates of bovine ESC-like colonies and ESC lines to increase to 16.7% and 28.6%, respectively. Hatched blastocysts were especially efficient in generating bovine ESCs (23.3% and 40.0%, respectively). Among the established ESC lines, four were made from day-9 hatched blastocysts. The expression levels of pluripotent markers in these lines were also high compared with those of ESC lines made from embryos at other developmental stages. This result might stem from the higher number of cells in the ICMs of the rapidly developing hatched blastocysts compared with those of the slowly developing expanded or hatching blastocysts. This result was in agreement with the reports of Verma et al. (2007) and Anand et al. (2009). We also employed a mechanical method for ICM cell isolation and subculture, in contrast to the ineffective method of enzyme dissolution using TrypLE or protease (Anand et al., 2009; Gong et al., 2010). Thus, in this study, efficient generation of ibESCs resulted from the use of healthy ICM cells and minimized ESC culture environment. Six JNU-ibES cell lines have been maintained for more than 90 days, subcultured for more than 12 passages, and successfully frozen (data not shown).
Unlike mESCs or hESCs (Evans and Kaufman, 1981; Thomson et al., 1998), the stemness, as measured by ESC marker expression, of ibESCs was inconsistent or conflicted in previous studies (Mitalipova et al., 2001; Saito et al., 2003; Wang et al., 2005). Mitalipova et al. (2001) and Saito et al. (2003) reported expression of both SSEA1 and SSEA4 in bovine ESCs, whereas a different study reported no expression of SSEA1 (Wang et al., 2005). Through replicated immunocytochemistry, we confirmed the expression of AP, Oct4, SSEA1, Nanog, and TRA-1-81 in ibESCs, but found no expression of SSEA4. Also, expression of SSEA1 was confirmed by the results of RT-PCR and semiquantitative RT-PCR. In addition, each of the ibESC lines presented different relative gene expression levels. The activity of Oct4 is essential for pluripotency in the founder cell population in the mammalian embryo (Nichols et al., 1998). Oct4 and ALP expression levels were higher in five of the ibESC lines (JNU-ibES-02–06) compared with JNU-ibES-01 (p<0.05). SSEA1 expression was higher in three of the ibESC lines (JNU-ibES-03, -05, and -06) (p<0.05) than in the others. Finally, Rex1, which encodes an acidic zinc finger protein, is expressed at a high level in four of the ibESC lines (JNU-ibES-03–06) (p<0.05).
Stem cells have the ability to choose between prolonged self-renewal and differentiation. This fate choice is highly regulated by intrinsic signals and the external microenvironment (Odorico et al., 2001). One study on the differentiation of ESCs, Schuldiner et al. (2000), reported that none of the growth factors direct differentiation exclusively to one cell type. In the present study, EBs derived from ibESCs were able to differentiate into three germ layers and also into specialized cells such as ectodermal cells [iris (Pax6) and adrenal (DbH)], mesodermal cells [bone (CMP) and muscle (enolase)], and endodermal cells [fetal liver (α-FP) and liver (albumin)]. In addition, bovine ESCs can be directed to differentiate into neuronal (Map2 and Tuj1) and glial (GFAP) cells in neuron cell culture medium. Moreover, the karyotype of JNU-ibES-05 was normal. In this study, JNU-ibES cells presented clear pluripotency characteristics in vitro. Together, these data will be valuable for the improvement of ibESC culture conditions.
The generation of bovine pluripotent stem cell lines may allow for complex genetic manipulations, including gene knockin and knockout technology, of potential application in industry. Further studies are being focused on redefining and improving protocols for the efficient generation of ibESCs through receptor assays or the development of homologous enriched feeder cell populations to extend culture longevity.
Footnotes
Acknowledgments
This study was supported by grants from the Bio-industry Technology Development Program (308008-5), Ministry for Food, Agriculture, Forestry and Fisheries, Next-Generation BioGreen 21 Program (PJ009075), and Cooperative Research Program for Agriculture Science & Technology Development (PJ009103), Rural Development Administration, Republic of Korea.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.