
Abstract
Abstract
Although telomeres are elongated during morula-to-blastocyst transition in cloned embryos, it is still unknown whether donor cell types have any effect on this elongation. In the present study, we examined the changes of telomere length during morula-to-blastocyst transition in cloned porcine embryos using different types of donor cells. Porcine embryonic stem–like cells (pESLCs), porcine cumulus cells (PCs), and porcine embryonic fibroblasts at passages 7 and 10 (PEF7s and PEF10s, respectively) were used as donor cells. Telomere lengths of pESLCs (35.8±1.5 kb), PCs (24.4±0.5 kb), PEF7s (18.7±0.6 kb), and PEF10s (17.2±0.1 kb) were significantly different. In contrast, telomere length in morulae derived from pESLCs (18.2±0.3 kb), PC (17.8±0.7 kb), PEF7 (18.5±0.3 kb), and PEF10 (18.4±0.4 kb) did not differ significantly. Likewise, telomeres in blastocysts derived from pESLCs (22.3±1.5 kb), PCs (23.5±2.6 kb), PEF7s (20.2±1.0 kb), and PEF10s (20.9±1.0 kb) had similar lengths. However, telomeres in blastocysts were significant longer (p<0.05) compared with morulae in each group. Relative telomerase activities of morulae derived from pESLCs (4.2±0.4), PCs (4.0±0.5), PEF7s (5.1±0.4), and PEF10s (4.9±0.4) were significantly lower (p<0.01) than those of blastocysts derived from pESLCs (8.2±1.1), PCs (8.6±0.6), PEF7s (12.5±2.9), and PEF10s (8.3±1.1). In conclusion, the telomere elongation in cloned pig embryos that occurred during morula-to-blastocyst transition may be related to the rise of telomerase activity. The telomere elongation may also be independent of the type and telomere length of the donor cell.
Introduction
Telomere lengths in cloned animals have been reported to be shorter (Alexander et al., 2007; Betts et al., 2005; Clark et al., 2003; Imsoonthornruksa et al., 2012; Konishi et al., 2011; Shiels et al., 1999), comparable (Betts et al., 2001; Imsoonthornruksa et al., 2012; Jiang et al., 2004; Kurome et al., 2008; Tian et al., 2000), or longer (Imsoonthornruksa et al., 2012; Jeon et al., 2005; Lanza et al., 2000; Wakayama et al., 2000) compared with age-matched controls and/or donor cells. Telomere length rescue in cloned animals is suggested to depend on the donor cell type (Miyashita et al., 2002). In a recent study, Schaetzlein et al. (2004) found that telomeres are elongated during morula-to-blastocyst transition in cloned bovine embryos using fibroblasts as donor cells. The elongation is likely a telomerase-dependent process (Schaetzlein et al., 2004) and represents a general mechanism rather than relating to the cloning process itself, because this observation has also been found in embryos derived from in vivo and in vitro fertilization (IVF) (Schaetzlein et al., 2004; Treff et al., 2011). However, it is uncertain whether different types of donor cells would have different patterns of telomere elongation during morula-to-blastocyst transition.
In the present study, we examined the telomere length during morula-to-blastocyst transition in cloned porcine embryos using different donor cell types. Porcine embryonic stem–like cells (pESLCs), porcine cumulus cells (PCs), and porcine embryonic fibroblasts (PEFs) at different passages were used as donor cells for nuclear transfer (NT).
Materials and Methods
Oocyte collection, in vitro maturation
Porcine ovaries were collected from prepubertal cross-bred gilts (Landrace×Large White) at a local abattoir and transported to the laboratory in Dulbecco's phosphate-buffered saline (D-PBS; Nissui Pharmaceutical Co. Ltd., Tokyo, Japan) at 35°C within 1 h. Cumulus–oocyte complexes (COCs) were aspirated from 3- to 6-mm follicles, and cultured in groups of 40–50 in 500 μL of modified North Carolina State University (NCSU)-37 medium (Petters and Wells, 1993) according to Kikuchi et al. (2002) in four-well dishes (Nunclon Multidishes, Nalge Nunc International, Roskilde, Denmark) for 22 h. The in vitro maturation (IVM) medium was modified by adding 10% (vol/vol) porcine follicular fluid, 0.6 mM cysteine (Sigma, St. Louis, MO, USA), 50 mM β-mercaptoethanol (β-ME; Axon Medchem, Groningen, The Netherlands), 1 mM dibutyryl cyclic adenosine monophosphate (dbcAMP; Sigma), 10 IU/mL equine chorionic gonadotropin (eCG; Serotropin, ASKA Pharmaceutical Co. Ltd., Tokyo, Japan), and 10 IU/mL human chorionic gonadotropin (hCG; Puberogen, Novartis Animal Health, Tokyo, Japan). The COCs were then transferred to IVM medium without dbcAMP and hormones and cultured for 22 h. IVM was performed in 5% CO2 and 20% O2 at 38.5°C.
Culture and preparation of donor cells for NT
The pESLC line was established from in vitro–produced blastocysts according to Haraguchi et al. (2012). The cells were subcultured at 37°C in a 5% CO2 atmosphere on mouse embryonic fibroblasts as feeder cells in ES medium consisting of Knock-Out Dulbecco's Modified Eagle Medium (KO-DMEM, Gibco, Grand Island, NY, USA) supplemented with 2 mM GlutaMax, 1% modified Eagle's medium (MEM) nonessential amino acids, 20% KnockOut Serum Replacement (KSR), 1% antibiotic-antimycotic liquid (all from Invitrogen, Carlsbad, CA, USA), 20 ng/mL human recombinant leukemia inhibitory factor (hLIF) (made in-house according to Haraguchi et al. 2012), and 0.1 mM β-ME, 6 μM CHIR99021, PD184352 (both from Axon Medchem), and 10 μM Y-27632 (Wako, Osaka, Japan). On the day of passage, colonies of pESLCs were detached from the culture plate by CTK solution consisting of 0.1% collagenase IV (Sigma), 0.25% trypsin (Invitrogen), 20% KSR, and 1 mM CaCl2 in PBS. This treatment helps eliminate the feeder cells because CTK solution does not have an effect on fibroblast cells. The clusters of pESLCs were then separated into single cells for NT by gentle pipetting.
After 44 h of IVM, cumulus cells were removed from oocytes by gentle pipetting after treated with 0.1% (wt/vol) hyaluronidase (Sigma). Cumulus cells were then collected, washed with PBS, and used for NT on the same day.
PEFs collected from a 46-day fetus (given by the Transgenic Pig Research Unit, National Institute of Agrobiological Sciences, Tsukuba, Japan) were cultured and prepared for NT according to Akagi et al. (2003). Briefly, PEFs were cultured in DMEM (Sigma) with 10% fetal bovine serum (FBS; Tissue Culture Biologicals, Tulare, CA, USA) and 1% antibiotic-antimycotic liquid at 37°C in a 5% CO2 atmosphere. The cultured cells were then frozen and stored at −80°C until use. Before NT, PEFs were serum-starved in DMEM supplemented with 0.5% FBS for 5 days after reaching approximately 80% confluence.
NT and in vitro culture
NT was performed using a Nikon Diaphot 300 inverted microscope (Nikon, Tokyo, Japan) equipped with Nikon Narishige hydraulic micromanipulator (Nikon) according to Akagi et al. (2008) with modifications. Briefly, after 44 h of maturation, oocytes were freed of cumulus by gently pipetting in the presence of 0.1% (wt/vol) hyaluronidase. Oocytes with an intact plasma membrane were selected and kept in tissue culture medium 199A (TCM-199A) supplemented with 20% (vol/vol) FBS until use. Oocytes with a first polar body were enucleated in TCM-199A supplemented with 20% FBS and 5 μg/mL Cytochalasin D (Sigma). A slit in the zona pellucida was made with the aid of a beveled needle. The first polar body and the surrounding cytoplasm were removed by squeezing the needle against the oocyte. Successful enucleation was confirmed by 20 μg/mL Hoechst 33342 (Sigma) staining under a fluorescence microscope (Nikon Diaphot 200, Nikon). Enucleated oocytes were kept in TCM-199A supplemented with 20% FBS until NT. Donor cells were transferred to the perivitelline space of enucleated oocytes. The oocyte–cell complex was sandwiched with a pair of electrodes in fusion medium consisted of 0.3 M mannitol, 1 mM CaCl2, 0.1 mM MgCl2, and 0.5 mM HEPES, and a single direct current pulse of 30 V for 10 μsec was applied for oocyte–cell fusion. Fused embryos were activated with a DC pulse of 1.5 kV/cm for 100 μsec using an Electro Cell Fusion (LF-100, Life Tec Co., Tokyo, Japan). Activated oocytes were transferred into porcine zygote medium (PZM)-5 (Yoshioka et al, 2008) and cultured for 6 days at 38.5°C under 5% CO2, 5% O2, and 90% N2.
Hoechst 33342 staining
To evaluate the total cell number in blastocysts, blastocysts were washed in PBS and treated for 5 min with 99.5% ethanol supplemented with 20 μg/mL Hoechst 33342. The blastocysts were then washed in glycerol, mounted on a glass slide, and covered with a cover slip. The blastocysts were flattened and the cells appeared on a plane. Total cell numbers were counted under an epifluorescent microscope (Nikon Diaphot 200, Nikon).
Extraction of total DNA and telomere length assays
Total DNA was extracted from pESLCs, PCs, PEF7s, PEF10s, morulae, and blastocysts by using the DNeasy Blood & Tissue Kit (QIAGEN, Hilden, Germany) following manufacturer's protocols.
Telomere length was measured by the monochrome multiplex quantitative polymerase chain reaction (MMQPCR) method according to Cawthon (2009) with modifications. PCR reactions were set up by aliquoting 18 μL of master mix into each reaction well of a 96-well plate compatible with LightCycler 480 Real-Time PCR System, followed by 2 μL of each experimental DNA sample, containing approximately 16 ng of DNA diluted in PCR-graded water, for a final volume of 20 μL per reaction. Five concentrations of a reference DNA sample (with the highest concentration being 6 ng/μL) spanning an 81-fold range of DNA concentration were prepared by serial dilution. These reactions provided the data for the generation of the standard curves used for relative quantitation.
The final concentrations of reagents in the PCR were 1×LightCycler 480 SYBR Green I Master (Roche Applied Science, Penzberg, Germany), 0.7 μM primers for telomeres (telg and telc), and 0.2 μM primers for single-copy gene albumin (albugcr1 and albdgcr1). The primer sequences for telomere used in this study were reported by Cawthon (2009). The forward and reverse primers for albumin (accession number, NM_001005208) were designed using the open software Primer3 ver0.4.0 (http://frodo.wi.mit.edu/primer3/). The primer sequences for albumin were: forward, CGGCGGCGGGCGGCGCGGGCTGGGCGGAGAACTGCTGGCTGGAACAT, and reverse, GCCCGGCCCGCCGCGCCCGTCCCGCCGGCCGATACACAGAGCACAGA. It should be noted that two GC-rich tagging sequences were attached to both primers for albumin to increase the primer melting temperature according to Cawthon et al. (2009). The thermal cycling profile was: Stage 1, 15 min at 95°C; Stage 2, 2 cycles of 15 sec at 94°C, 60 sec at 49°C; Stage 3, 4 cycles of 15 sec at 94°C, 30 sec at 59°C; Stage 4, 20 cycles of 15 sec at 85°C, 30 sec at 59°C with single acquisition; and Stage 5, 27 cycles of 15 sec at 94°C, 10 sec at 84°C, 15 sec at 85°C with single acquisition. The 59°C reads in Stage 4 provided the Ct values for the amplification of the telomere template (in early cycles when the scg signal is still at baseline); the 85°C reads in Stage 5 provided the Ct values for the amplification of the albumin template (at this temperature there is no signal from the telomere PCR product, because it is fully melted). Double melting peaks of telomere and albumin amplicons following 20 cycles of amplification reaction for telomere and 27 cycles of amplification reaction for albumin are shown in Supplementary Data (Fig. S1). (Supplementary Data are available at www.liebertonline/cell/.)
After thermal cycling and raw data collection were complete, the LightCycler 480 software (Roche Applied Science) was used to generate two standard curves for each plate, one for the telomere signal and one for the albumin signal. The T/S ratio for an experimental DNA sample is T, the quantitation of the standard DNA that matches the experimental sample for copy number of the telomere template, divided by S, the quantitation of the standard DNA that matches the experimental sample for copy number of single copy gene albumin. As three biological replications were performed, and each experimental DNA samples were assayed in triplicate, nine T/S quantitation values were obtained for each sample. The final reported result for a sample in a given run is the average of the nine T/S values. The telomere lengths were calculated using Cawthon's formula (2009) based on T/S values.
The telomere lengths of donor cells were confirmed by Terminal Restriction Fragment (TRF) assay using TeloTAGGG kit (Roche Applied Science) by following the manufacture's protocol.
Telomerase activity assays
Telomerase activities in morulae and blastocysts were evaluated by using the TRAPEZE Telomerase Detection Kit (Milipore, Billerica, MA, USA) according to the manufacturer's protocol. Briefly, 10 morulae or blastocysts were resuspended in 50 μL of 1×CHAPS lysis buffer. The suspensions were incubated on ice for 30 min. The samples were centrifuged at 12,000×g for 20 min at 4°C. The extracts were then quickly frozen on dry ice and stored at −80°C until use. Telomerase activities in morulae and blastocysts were determined using SYBR Green real-time quantitative telomeric repeat amplification protocol (RQ-TRAP) assay according to Wege et al. (2003) with modifications. The SYBR Green RQ-TRAP assay was conducted with 5-μL extracts (equivalent to one blastocyst/morula), 0.1 μg of telomerase primer TS, and 0.05 μg of anchored return primer ACX, in a 25-μL reaction using LightCycler 480 SYBR Green 1 Master (Roche Applied Biosystems). Primer sequences were as described by Kim and Wu (1997). Samples were incubated for 20 min at 25°C and amplified in 35 PCR cycles with 30 sec at 95°C and 90 sec at 60°C (two-step PCR). All PCRs were performed in triplicate on each plate on the LightCycler 480 Instrument (Roche Applied Science).
Five concentrations of a reference sample (positive telomerase extract control supplied in TRAPEZE Telomerase Detection Kit) spanning a 16-fold range of DNA concentration were prepared by serial dilution. These reactions provided the data for the generation of the standard curves used for relative quantitation. Because three biological replications were performed, and each experimental sample was assayed in triplicate, nine relative quantitation values were obtained for each sample. Relative quantitation of telomerase activity was expressed relative to reference sample by the standard curve method. Inactivated (heat-treated) samples and lysis buffer were also assayed on every plate for negative control for each sample and primer-dimer/PCR contamination control, respectively.
Experimental design
Telomere length in different donor cell types
The telomere length of pESLCs, PCs, PEF7s, and PEF10s were measured according to MMQPCR method, and the results were confirmed by TRF assays.
Telomere length in cloned morula and blastocysts
The telomere lengths in morulae/blastocysts derived from pESLCs, PCs, PEF7s, and PEF10s were measured. Morulae were collected on day 5 whereas blastocysts were collected on day 6. Morulae were defined as embryos with more than eight cells. Blastocysts were defined as embryos with clear blastocoels and without any degenerated part.
Telomerase activity in cloned morula and blastocysts
Telomerase activities in cloned morulae/blastocysts derived from pESLCs, PCs, PEF7s, and PEF10s were determined. Morulae were collected on day 5 whereas blastocysts were collected on day 6. Morulae were defined as embryos with more than eight cells. Blastocysts were defined as embryos with clear blastocoels and without any degenerated part.
Statistical analysis
The data of blastocyst rate, total cell numbers in blastocysts, telomere lengths, and telomerase activities were analyzed by one-way analysis of variance (ANOVA) followed by a Tukey test by using general linear models in Statview 5 software package (SAS Institute Inc., Cary, NC) to determine differences. Data regarding the percentage of blastocysts were arcsine transformed before analysis. Random distribution of NT embryos was made in each experimental group. NT experiments were replicated seven times. For telomere length and telomerase activity assays, three pooled samples were collected for each group, and each sample was assayed in triplicate. Data were expressed as mean±standard error of the mean (SEM). A probability of p<0.05 was considered to be statistically significant.
Results
Telomere length in different donor cell types
Using MMQPCR, we found that telomere length in pESLCs (35.8±1.5 kb), PCs (24.4±0.5 kb), PEF7s (18.7±0.6 kb), and PEF10s (17.2±0.1 kb) were significantly different from each other. The telomere length of donor cells were confirmed by TRF assays using TeloTAGGG Telomere Length Assay kit (Fig. 1). Telomere length in pESLCs (35.1±1.7 kb), PCs (22.9±1.1 kb), PEF7s (19.7±0.4 kb), and PEF10s (17.7±0.6 kb) obtained by using TRF assays were similar to those by MMQPCR.
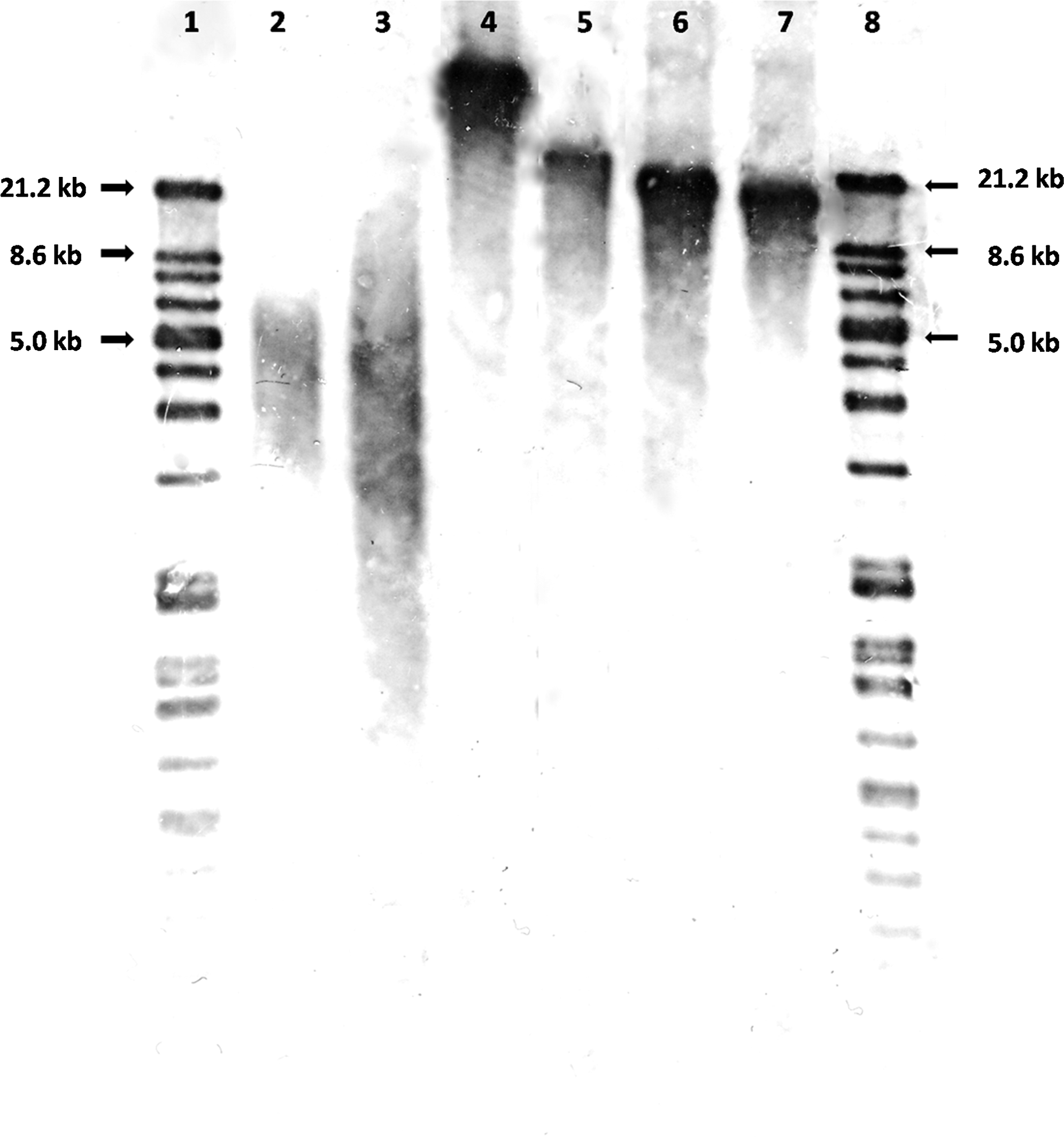
Telomere length in different donor cell types. Telomere length was determined by TRF assay using the TeloTAGGG Telomere Length Assay kit (Roche Applied Science) to confirm the results on telomere length measured by MMQPCR. The telomere length in pESLCs (35.1±1.7 kb), PCs (22.9±1.1 kb), PEF7s (19.7±0.4 kb), and PEF10s (17.7±0.6 kb) were different. The results corresponded to those obtained by using MMQPCR method. Lanes 1 and 8, DIG marker; lane 2, K562 cells; lane 3, DNA control (purified genomic DNA from immortal cell lines provided by the kit); lane 4, pESLCs; lane 5, PCs; lane 6, PEF7s; lane 7, PEF10s.
Telomere length in cloned morulae and blastocysts
Although telomeres of donor cells had significantly different lengths, cloned morulae derived from pESLCs, PCs, PEF7s, and PEF10s did not have any significant differences in telomere length (Fig. 2). Likewise, cloned blastocysts derived from pESLCs, PCs, PEF7s, and PEF10s had similar telomere lengths (Fig. 2). However, blastocysts had longer telomeres (p<0.05) compared with morula-stage embryos in each respective group.
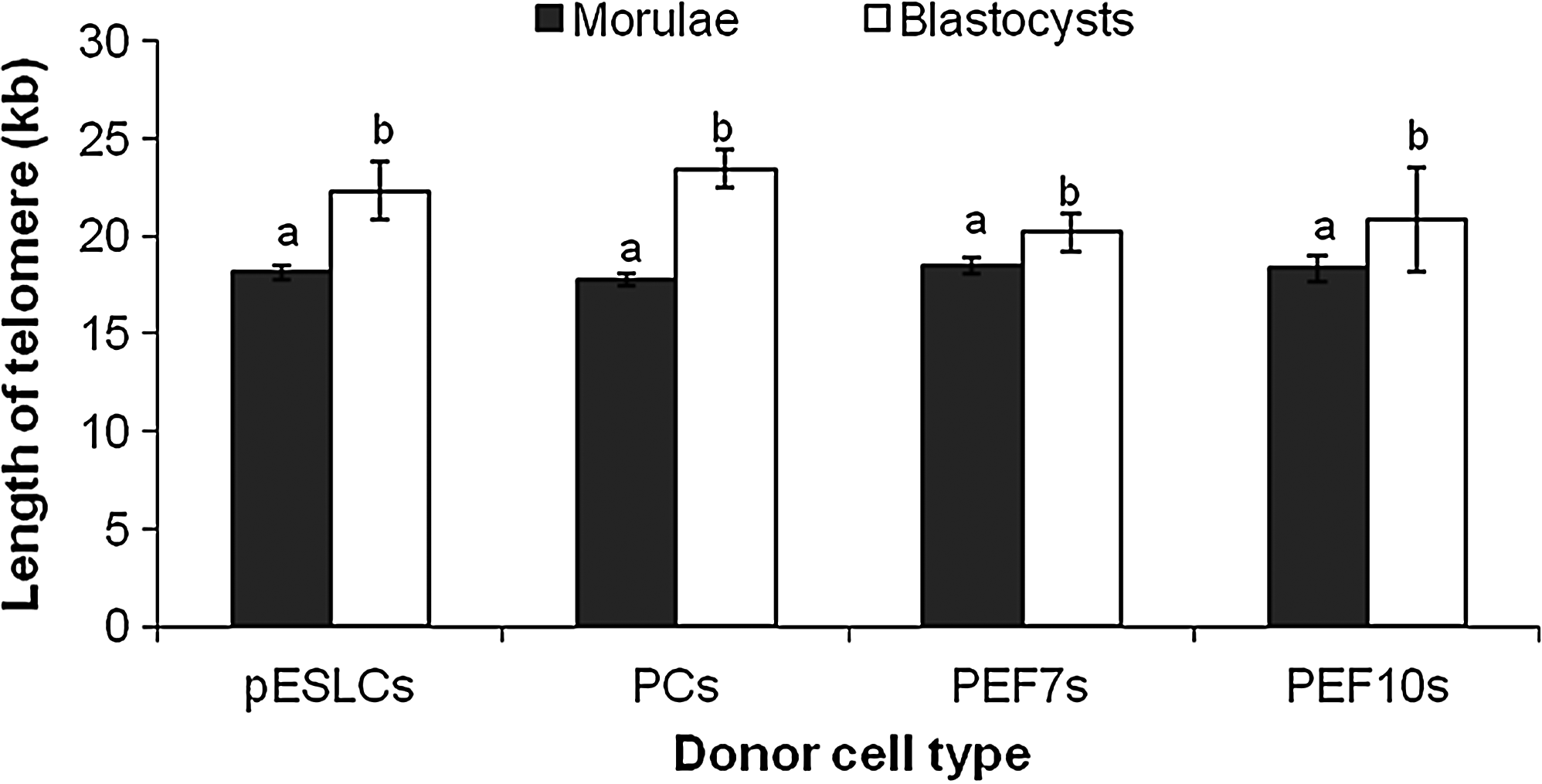
Telomere length in cloned embryos derived from different donor cell types. Telomere length was measured by MMQPCR method. Blastocysts had significantly longer telomeres compared with morula in each group. Different alphabetic letters denote significant differences (p<0.05). Three biological replications were performed.
Telomerase activity in cloned morulae and blastocysts
Relative telomerase activities of cloned morulae derived from pESLCs, PCs, PEF7s, and PEF10s were significantly lower (p<0.01) than those of cloned blastocysts derived from pESLCs, PCs, PEF7s, and PEF10s (Fig. 3).
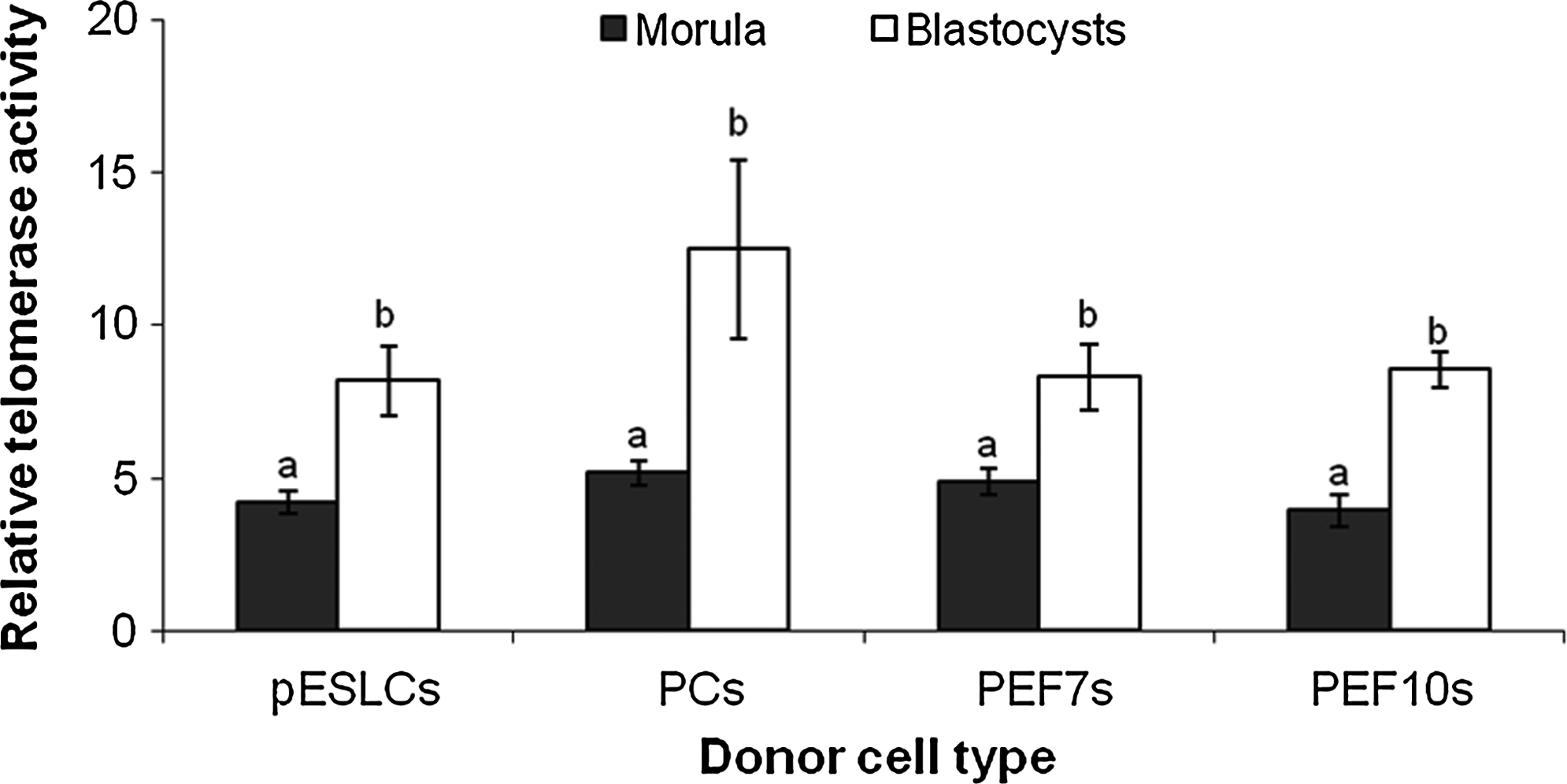
Relative telomerase activities in cloned embryos derived from different donor cell types. Telomerase activities were determined using the SYBR Green RQ-TRAP assay. Blastocysts had significantly higher telomerase activities compared with morula in each group. Different alphabetic letters denote significant differences (p<0.01). Three biological replications were performed.
Discussion
In the present study, we demonstrated that telomere length is elongated during morula-to-blastocyst transition in porcine cloned embryos. This is consistent with a previous report in cloned bovine embryos (Schaetzlein et al., 2004). Telomere restoration has been proven to represent a general mechanism rather than only being related to the cloning process itself because telomere length restoration is also observed in in vivo and IVF embryos in humans (Treff et al., 2011), cattle, and mouse (Schaetzlein et al., 2004). The reason for telomere lengthening during morula-to-blastocyst transition is thought to link with telomere activity because a rise in telomerase activity in the blastocyst stage compared with cleavage and morula stages was observed in cloned embryos (Schaetzlein et al., 2004; Xu and Yang, 2001).
In this study, we also found an increase in telomerase activity during morula-to-blastocyst transition in cloned porcine embryos, suggesting that telomere lengthening in early embryonic development is also likely related to telomerase activities in pigs. Although it seems that there is a close correlation between the telomere elongation and the increase of telomerase activity in blastocysts compared with cleaved embryos and morulae, there are other factors that might contribute to this process, such as loss of histone modifications (Garcia-Cao et al., 2004) and DNA methylation at subtelomeric regions (Gonzalo et al., 2006) as found in somatic and embryonic stem cells in mouse. Further studies are needed to clarify the exact mechanism for this observation.
In a previous report, telomere length rescue in cloned cattle appeared to depend on the used donor cell type (Miyashita et al., 2002). This suggests the possibility that the pattern of telomere elongation in cloned embryos during morula-to-blastocyst transition might be different according to the donor cell types used for NT. Therefore, we examined the telomere lengths in cloned morulae and blastocysts derived from different donor cell types: pESLCs, PCs, and PEFs. Our first experiment has demonstrated significant differences in telomere lengths among the donor cell types. However, after NT we found that regardless of donor cell types the resultant morulae had similar telomere lengths. Furthermore, the telomere increased to a similar length in blastocysts among groups. This suggests that the telomere elongation in porcine cloned embryos during reprogramming is independent of the donor cell type. It should be noted that the developmental competence (i.e., blastocyst formation rate and total cell number in blastocysts) of cloned embryos derived from different donor cells was not significantly different in this study (see Table S1, Supplementary Data).
We revealed that although different cell types (pESLCs, PC, and PEFs) or the same cell type (PEF7 and PEF10) with different telomere lengths was used for NT, the lengthening of telomeres still occurred in all groups during the transition from morula to blastocysts. This suggests that the telomere restoration during morula-to-blastocyst transition is independent of telomere length of donor cell types in pigs. This is in agreement with a previous study in cattle (Schaetzlein et al., 2004). It should be noted that telomere lengths of pESLCs were found to be longer than those of blastocysts in our study. This is possibly caused by continuous elongation of telomeres in pluripotent cells that are locked at an epigenetic state (Marion et al., 2009).
In conclusion, telomere elongation in cloned pig embryos that occurred during morula-to-blastocyst transition might be related to the rise of telomerase activity. The telomere elongation may also be independent of the type and telomere lengths of the donor cell.
Footnotes
Acknowledgments
We would like to thank Dr. Akira Onishi and Transgenic Pig Research Unit (National Institute of Agrobiological Sciences, Tsukuba, Japan) for giving us the porcine embryonic fibroblasts, Dr. Tadashi Furusawa for critical discussion, and Ms. Ando and Ms. Suzuki for technical assistance.
Author Disclosure Statement
The authors declare that no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.