
Abstract
Abstract
Induced pluripotent stem cells (iPSCs) can be artificially reprogrammed from somatic cells by overexpression of exogenous transcription factors. The pig has increasingly become an important large animal model for preclinical tests and studies of human diseases; thus, the generation of porcine iPSCs will facilitate research into the efficacy and safety of stem cell therapy. A current major problem facing the generation of porcine iPSCs is the failure to silence exogenous transgenes. We hypothesized that this problem can be resolved by reducing the number of transcriptional factors used for porcine iPSCs induction. Here, we report the successful generation of porcine iPSCs using the porcine factors Oct4 and Klf4 in combination with specific small molecules. In comparison with high oxygen conditions (20%), the efficiency of porcine iPSC generation was higher under low oxygen conditions (5%). Porcine iPSCs exhibited a normal karyotype and morphology, like mouse embryonic stem cells (ESCs), and could proliferate in the absence of basic fibroblast growth factor (bFGF) and in the presence of human leukemia inhibitory factor (hLIF) and mouse embryonic fibroblast feeder cells. These iPSCs also expressed ESC-like markers (Oct4, Nanog, Klf4, c-Myc, Bmp4, bFgf). Importantly, the porcine iPSCs showed pluripotency, as evidenced by differentiation into three germ layers in vitro following embryoid body formation, as well as by efficiently forming teratomas containing three germ layers in immunodeficient mice. Thus, pluripotent porcine iPSCs can be generated from somatic stem cells by using only two porcine transcription factors in combination with small molecules. These attempts represent the first step toward generating truly pluripotent porcine iPSCs with fewer exogenous genes and less integration.
Introduction
The pig is a useful model for evaluating both human diseases and safety of iPSCs, because pigs are more physiologically similar to humans than are mice, and pigs have an organ size conducive to studies of organ transplantation (Brandl et al., 2007; Kolber-Simonds et al., 2004; Prather et al., 2003). Despite this, no stable pluripotent porcine ESC lines from the inner cell mass (ICM) have been established (Brevini et al., 2007; Hall, 2008; Vackova et al., 2007), although the reasons for this are unclear. Rather, the majority of porcine iPSCs have been generated using the four human OSKM factors with the addition of basic fibroblast growth factor (bFGF) (Esteban et al., 2009; Ezashi et al., 2009; Telugu et al., 2010; West et al., 2010; Wu et al., 2009). However, the ectopic expression of c-Myc causes tumorigenicity, and retroviruses themselves can cause genome mutagenesis (Meyer and Penn, 2008).
The major problem currently facing the development of porcine iPSCs is the failure to silence exogenous transcription genes (Wu et al., 2009), which may limit the activation of endogenous genes (Takahashi et al., 2007). Moreover, the integration of exogenous genes can cause extensive genetic mutations in the genome (Yu et al., 2007), and safety issues limit the potential applications of iPSCs (Carey et al., 2009; Okita et al., 2007; Sommer et al., 2009). This could partially be addressed through the introduction of fewer exogenous factors, but decreasing the number of transcription factors used to induce iPSCs reduces the efficiency of this process (Li et al., 2011; Zhu et al., 2010). Notably, low oxygen conditions and the introduction of small molecules have been reported to improve the generation of iPSCs (Huangfu et al., 2008; Lin et al., 2009; Yoshida et al., 2009).
We attempted to generate porcine iPSCs under low oxygen conditions and in the presence of a combination of five inhibitors (5i) to enhance reprogramming efficiency and avoid the need for Sox2 and c-Myc. The inhibitors used included sodium butyrate (NaB, 0.25 mM), a histone deacetylase inhibitor; SB431542 (SB, 2 μM), an activin receptor-like kinase (ALK) inhibitor; PD0325901 (PD, 1 μM), an inhibitor of the mitogen-activated protein kinases/extracellular signal-regulated kinases (MAPK/ERK) pathways; forskolin (FK, 10 μM), a cyclic adenosine monophosphate (cAMP) stimulator; and CHIR99021 (CH, 3 μM), a GSK3 inhibitor (Hanna et al., 2010; Lin et al., 2009; Ying et al., 2008; Zhu et al., 2010). We report here that porcine iPSCs can be reprogrammed from somatic stem cells under low oxygen conditions by using a reduced number of porcine factors in combination with small molecules.
Materials and Methods
Cell culture
Porcine mesenchymal stem cells (MSCs) derived from 35- to 45-day-old embryos by using a standard procedure were cultured in high glucose Dulbecco's modified Eagle medium (DMEM) culture medium (Invitrogen) containing 1% penicillin-streptomycin, 0.1 mM nonessential amino acids (Invitrogen) and 10% fetal bovine serum (FBS; Hyclone) (Peister et al., 2004). The porcine iPSCs were maintained on B6D2F1 mouse embryonic fibroblasts (MEFs) treated with mitomycin C in DMEM/F12 medium supplemented with 20% KnockOut Serum Replacement (KSR) or FBS (Hyclone), 1% penicillin-streptomycin, 0.1 mM nonessential amino acids, 1 mM
Retrovirus production and reprogramming of MSCs
Retroviruses were produced and harvested following the protocol of the George Q. Daley lab (Park et al., 2008). Before infection, 1×105 MSCs per well were plated in a six-well dish. After 2 days of recovery, MSCs infected with retrovirus-transduced pMXs porcine factors Oct4 and Klf4 were induced under four different conditions (either high or low oxygen, in the presence or absence of 5i), and the MSC medium was changed to porcine ES medium. Ten days after infection, 5×104 cells were passaged onto MEF feeder cells, after which the medium was changed every day. ESC-like colonies were picked at days 23–25 following a standard protocol.
Alkaline phosphatase staining and immunofluorescence microscopy
Alkaline phosphatase (AP) staining was performed using an Alkaline Phosphatase Substrate Kit iii (Vector, sk-5300) according to the instruction manual. For immunostaining, iPSCs were washed in phosphate-buffered saline (PBS), and then fixed in 3.7% paraformaldehyde for 30 min. Cells were permeabilized with 0.1% Triton X-100 in PBS and blocked with blocking solution (3% goat serum in PBS) for 2 h before being incubated overnight at 4°C in Oct4 (sc9081, Santa Cruz), Nanog (Abcam, ab21603), SSEA-1 (MAB4301), SSEA-4 (MAB4304), or TRA-1-81 (MAB4381) primary antibodies diluted 1:200. After three PBS washes, cells were incubated with a secondary antibody [1:200, Alexa Fluor 568 goat anti-rabbit immunoglobulin G (IgG) or Alexa Fluor 488 goat anti-mouse; Molecular Probes, Invitrogen] for 2 h at room temperature. Samples were washed three times, and nuclei were visualized with Vectashield (Vector) after being incubated with 10 μg/mL Hoechst 33342 (Sigma) for 5 min. Fluorescence images were captured using a Nikon inverted fluorescence microscope.
Teratoma formation
Porcine iPSCs (1×106) or an equivalent number of MSCs as controls were injected subcutaneously into immunodeficient mice (SCID). After 7–10 weeks, the mice were sacrificed to assess teratoma formation. Teratomas were fixed, sectioned, and processed for Hematoxylin & Eosin (H&E) staining.
Karyotype analysis
Porcine MSCs and iPSCs were arrested in 0.3 μg/mL nocodazole for 3 h to enrich cells at metaphase. Cells were collected and treated with 75 mM KCl for 30 min. After centrifugation at 300×g for 8 min, cell pellets were fixed three times in 1:3 cold acetic acid:methanol for 30 min. For chromosome analysis, fixed cells were dropped onto precooled slides and dried overnight. More than 30 chromosome spreads were counted, and the normal karyotype percentage was recorded. A percentage of above 70% was considered normal. For metaphase analysis, fixed cells were dried at room temperature for 2 days, and then digested in 0.05% trypsin at room temperature and stained with 10% Giemsa (Invitrogen).
RNA extraction and quantitative real-time PCR
Total RNA was isolated from porcine testis or cells by using the RNeasy Mini Kit (Qiagen) and reverse transcribed using M-MLV Reverse Transcriptase (Invitrogen). cDNA was used as a template for reverse transcriptase polymerase chain reaction (RT-PCR). Quantitative real-time PCR (qRT-PCR) reactions were performed with the FastStart Universal SYBR Green Master (Roche) using an iCycler iQ5 2.0 Standard Edition Optical System (Bio-Rad). Primers were either as previously described or designed using the IDT DNA website, as follows: endo-Oct4, forward (F) (CTT CAC CAC CCT GTA CTC CTC) and reverse (R) (GCT TCT CTC CCT AGC TCA CC); ex-Oct4, F (GAC GGC ATC GCA GCT TGG ATA CAC) and R (GAG AAG GCG AAG TCG GAA G); endo-Klf4, F (GCA TTT TCA AGG TCG GAC CAC) and R (CCT CAC CAG GGA GAC AGT GTG); ex- Klf4, F (GAC GGC ATC GCA GCT TGG ATA CAC) and R (GTC TTT GCT TCA TGT GGG); Nanog, F (AGG ACA GCC CTG ATT CTT CCA CAA) and R (TCT TCT GCT TCT TGA CTG GGA CCT); Sox2, F (AGG ACA GCC CTG ATT CTT CCA CAA) and R (TCT TCT GCT TCT TGA CTG GGA CCT); Myc, F (CAA GAG GGC TAA GTT GGA CAG TGG) and R (GCT GCG TTT CAG CTC GTT TCT C); Lin28, F (TCT TTG TGC ACC AGA GTA AGC) and R (ATT CCA AGC CCT TAG CAG AC); β-actin: F (TGC GGC ATC CAC GAA ACT AC) and R (TTC TGC ATC CTG TCG GCG AT); bFgf, F (GCG ACC CTC ACA TCA AAC TAC) and R (AGC CAG TAA TCT TCC ATC TTC C); P53, F (AGT GGC AAC TTG CTG GGA CG) and R (TGG TGG GCA GTG CTC GCT TA); Lif, F (CAC TGG AAA CAC GGG GCA) and R (AGG GCG GGA AGT TGG TCA); Bmp4, F (CTC GCT CTA TGT GGA CTT CAG) and R (GAT GGC ATG ATT GGT TGA GTT G); alphaAT, F (AGC CTT CTC CAT CAA CTT CAG) and R (AGT TCA CCA GAG CAA ATA CGG); enolase, F (AGA ACA ATG AAG CCC TGG AG) and R (GTC GAG ATC ATA CTT CCC GTT G); and NeuroD, F (AAA TCA TAC AGC GAG AGT GGG) and R (AGT CCT CTT CCG CAT TCA TG). The relative expression level of the target genes was normalized by using β-actin as an internal control and calculated using the Ct method. (A fit sample was chosen as the relative one and the relative expression among different genes was calculated by the same threshold.) The specificity of the PCR products was confirmed by melting curve analysis and reverse-transcription PCR.
Embryoid body formation and differentiation
Porcine iPSCs were harvested by trypsinization and transferred to low-adhesive 35-mm noncoated plates before being cultured in MSC medium. After 4–7 days, aggregated cells were selected, transferred onto gelatin-coated tissue culture dishes, and incubated for another 5–7 days before the differentiated cells were stained with α-smooth muscle actin (SMA; Abcam, ab5694-100), βIII-tubulin (Abcam, CBL412), and alpha-fetoprotein (AFP; DAKO, DAK-N150130).
Flow cytometry
Fluorescence-activated cell sorting (FACS) analysis of the MSC and porcine iPSCs were carried out using a BD LSR analyzer (BD Biosciences), and data were analyzed by CELLQuest Pro. Antibodies used for this study were the same as those used for immunofluorescence above. Immunostaining without primary antibodies served as negative controls.
Results
Reprogramming porcine MSCs into ESC-like colonies by using porcine Oct4 and Klf4
Germ-line stem cells from neonatal mouse testis are pluripotent and have an ability to differentiate similar to that of ESCs (Kanatsu-Shinohara et al., 2004). Therefore, pluripotent factors were initially cloned from porcine testis using RT-PCR, and their expression levels were compared to those in porcine bone marrow–derived MSCs. qRT-PCR was performed and analyzed by the cycle threshold (CT) value. Like testis, MSCs also expressed a few pluripotent genes, including Klf4, c-Myc, Bmp4, and Nanog, and reduced levels of Oct4, Sox2, and Lin28 (Fig. 1A).

Porcine iPSCs generated from MSCs using Oct4 and Klf4 (OK) porcine factors. Low oxygen and treatment with small molecules are shown to have enhanced the reprogramming of the porcine MSCs. (
When MSCs expressing exogenous Oct4 and Klf4 were induced under four different conditions (high oxygen with 5i, high oxygen without 5i, low oxygen with 5i, and low oxygen without 5i) and passaged onto feeder cells on day 10, they formed ESC-like colonies by days 16–20. Colonies were subsequently picked on day 25. The reprogramming scheme and cell morphologies observed on days 0, 10, 20, and at passage 5 are shown in Figure 1, B and C. The porcine iPSCs lines maintained in 20% KSR medium supplemented with 1000 U/mL hLIF exhibited a compact morphology, clear boundaries, and a high nuclei-to-cytoplasm ratio (Fig. 1C).
When an attempt was made to induce iPSCs generation using only Oct4 and 5i, it was found that although donor cells exhibited some morphological changes, they were unable to form colonies. The detailed groups and morphologies on day 20 are presented in Figure 1D. Furthermore, AP staining as an initial marker of ESCs was negative (Fig. 1F, control).
Early stem cell properties and reprogramming efficiency under different conditions
Low oxygen was more effective at producing AP-positive cells by day 30, because the number of AP-positive colonies formed in the low oxygen condition was nearly three times greater than under the high oxygen condition, regardless of the inclusion of 5i (Fig. 1E,F). Interestingly, porcine iPSCs showed different morphologies and efficiencies for the establishment of cell lines, depending on the conditions in which they were grown. Colonies grown in low oxygen (5%) were generally larger than those grown in high oxygen (atmosphere at 20%), and colonies grown in the presence of 5i were more compact than those that were not, regardless of the oxygen concentration (Fig. 1F). Therefore, we grew colonies under low oxygen conditions in the presence of 5i in the majority of subsequent experiments. Furthermore, while 40 of the 48 picked colonies that had been grown in the presence of 5i survived the initial trypsin-EDTA passage, only 4 of the 24 picked colonies that had not been treated with 5i survived. After initial trypsin-EDTA passage, we subsequently digested the iPSCs with collagenase IV and passaged them onto feeder cells.
We analyzed the expression of exogenous Oct4 and Klf4 in the porcine iPSCs. After five passages, cells were collected, and qRT-PCR was used to calculate the mRNA copy number. Although exogenous gene expression remained high in the iPSCs, after several passages we found that the low oxygen plus 5i condition produced cells that were best able to maintain mouse ESC-like morphology and had lower exogene expression than was detected under low oxygen only or high oxygen plus 5i conditions (Fig. 2A).
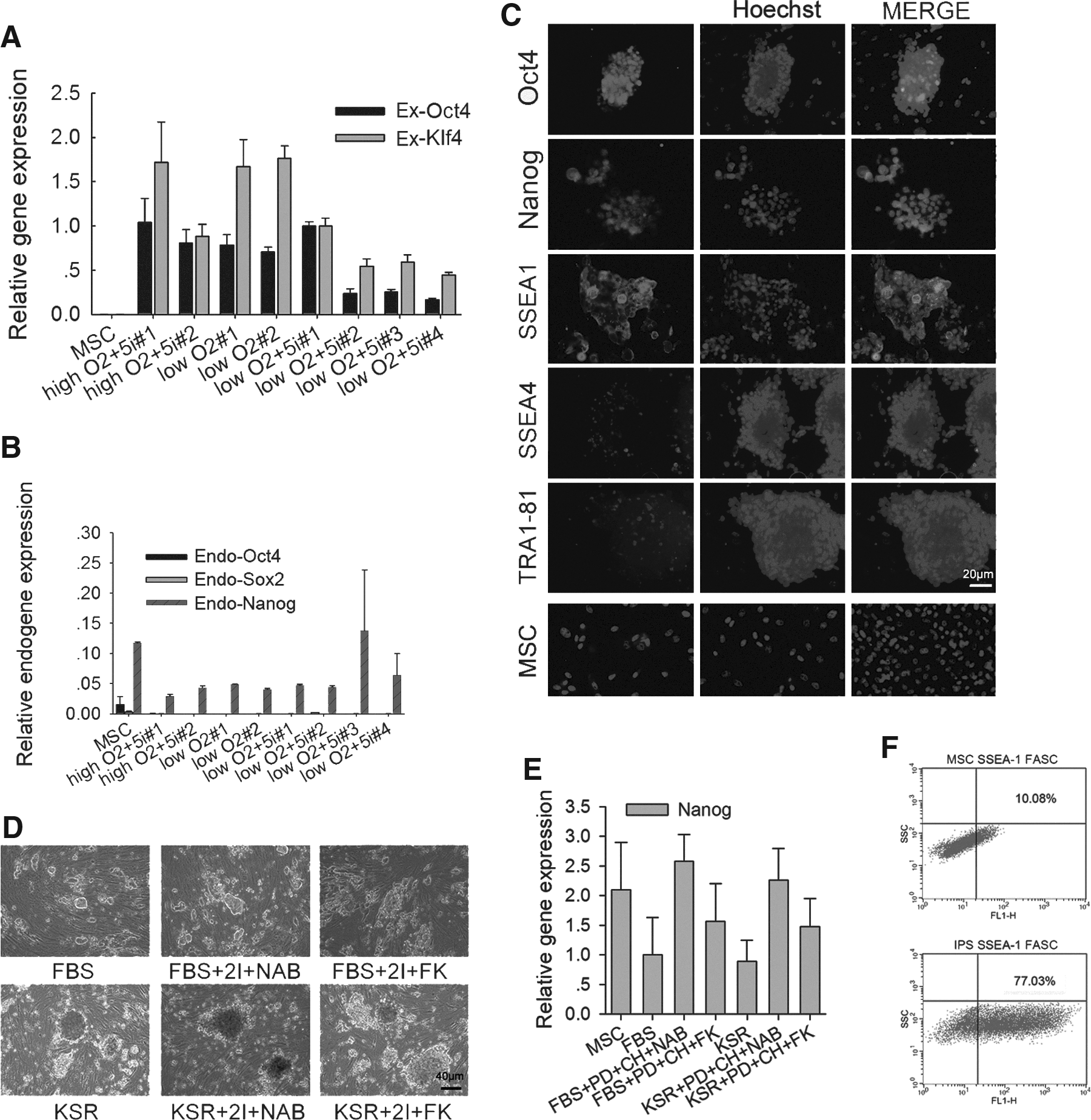
Characterization of porcine iPSCs. (
Expression of endogenous pluripotent genes in porcine iPSCs
Like those of MSCs, endogenous Oct4, Sox2, and Nanog were not distinctly reactivated in the iPSCs colonies examined by quantitative PCR analysis (Fig. 2B). However, immunofluorescence staining showed that the majority of porcine iPSCs expressed Nanog. These iPSCs also expressed Oct4, as well as the specific mouse ESC-like pluripotent surface marker stage-specific embryonic antigen-1 (SSEA-1). Few of the colony cells expressed very little amounts of human ESC-like pluripotent surface markers SSEA-4 and TRA-1-81. In contrast to porcine iPSCs, the progenitors MSCs expressed only minimal or very low levels of Nanog and SSEA-1. Immunostaining without the primary antibodies served as negative controls showed only background staining (Fig. 2C).
To achieve active expression of endogenous genes, colonies were subcultured in different culture media from passage 5 to passage 10. Images of cell lines and Nanog expression at passage 10 are shown in Figure 2, D and E. We found that incubation in media supplemented with KSR resulted in the formation of colonies that were both larger and less dense than those formed in media supplemented with FBS. Furthermore, treatment with the small molecules PD and CH (2i) plus NaB or FK did not have any detectable positive effect with regard to Nanog activation (Fig. 2E), and the expression of exogenes remained high (Fig. 3A).
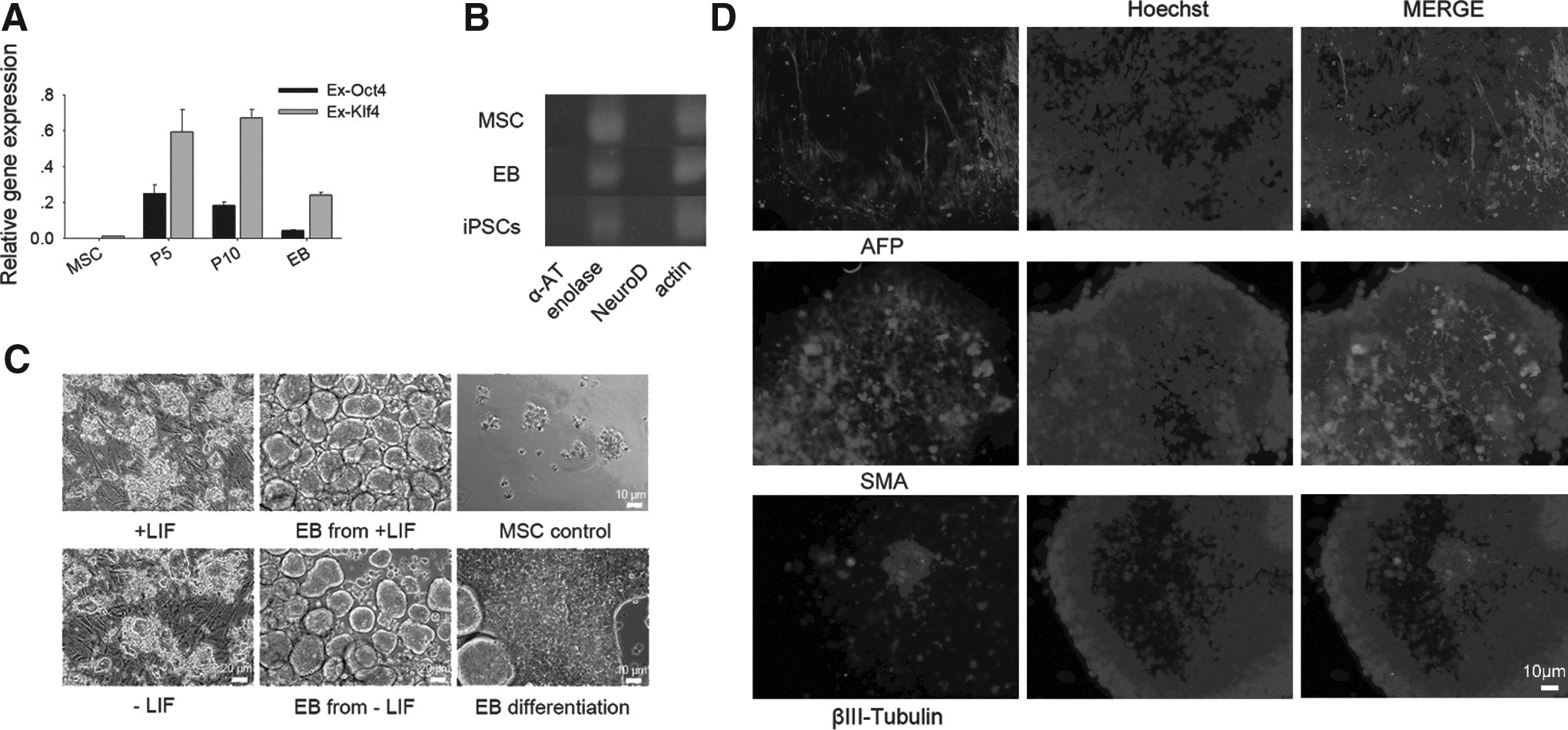
In vitro differentiation of porcine iPSCs. (
Interestingly, SSEA-1+ mesenchymal cells have been identified (Anjos-Afonso and Bonnet, 2007). We performed FACS analysis and also found that 10% of MSCs (P10) were positive for the mouse ESC pluripotent surface marker SSEA-1; in contrast, 77% of iPSCs were positive for SSEA-1 (Fig. 2F). The porcine iPSCs with mouse ESC-like morphology express high levels of SSEA-1, but low levels of SSEA-4 and TRA-1-81. This is different from other porcine iPSCs reported previously that express SSEA-1 with human ESC-like morphology or SSEA-1/4 with mouse ESC-like morphology (Ezashi et al., 2009; Telugu et al., 2010). However, the porcine iPSCs generated by the two factors expressed pluripotency markers Nanog and SSEA1 by both immunostaining and flow cytometry, apparently different from their progenitors MSCs.
Porcine iPSCs are able to differentiate into three germ layers in vitro
We chose two of the iPSCs lines, low oxygen+5i #3 (LI3) and low oxygen+5i #4 (LI4), to test embryoid body (EB) formation. After 4–7 days, the EBs were moved into tissue culture dishes, where half of them were observed to attach and initiate differentiation. Regardless of LIF, porcine iPSCs maintained similar colony formation and morphology and robust EB formation (Fig. 3C), suggesting that hLIF is dispensable for porcine stem cell proliferation and differentiation. When iPSCs formed EBs, exogenes of iPSCs decreased nearly half of initiation (Fig. 3A). After another 7 days, immunostaining detected cells positive for alpha smooth muscle actin (SMA, mesoderm marker), alpha-fetoprotein (AFP, endoderm marker), and punctate βIII-tubulin (ectoderm marker; Fig. 3D). However α-1 antiproteinase (α-AT, endoderm marker) and NeuroD (ectoderm marker) were not detected by RT-PCR in MSCs, EBs, and iPSCs, whereas all of them expressed enolase (mesoderm marker) (Fig. 3B).
In vivo differentiation and analysis
Severe combined immunodeficient (SCID) mice injected with iPSCs (LI3 and LI4) that possessed more than 70% normal 38 chromosomes by karyotype formed teratomas with clear blood vessels, whereas control mice did not (Fig. 4B,C). Porcine iPSCs had been subcultured more than 10 passages (over approximately 50 days), and the normality of karyotypes was further confirmed by G-banding chromosomal analysis (normal 38 XY) (Fig. 4A). Histological examination showed that the teratomas included various tissues from the three germ layers—the neuronal epithelium (ectoderm), gland-like cells with blood vessel and cancer-like cell (endoderm), and muscle and adipose tissue (mesoderm; Fig. 4D). These results indicate that the iPSCs thus generated have the capacity to differentiate into three germ layers.
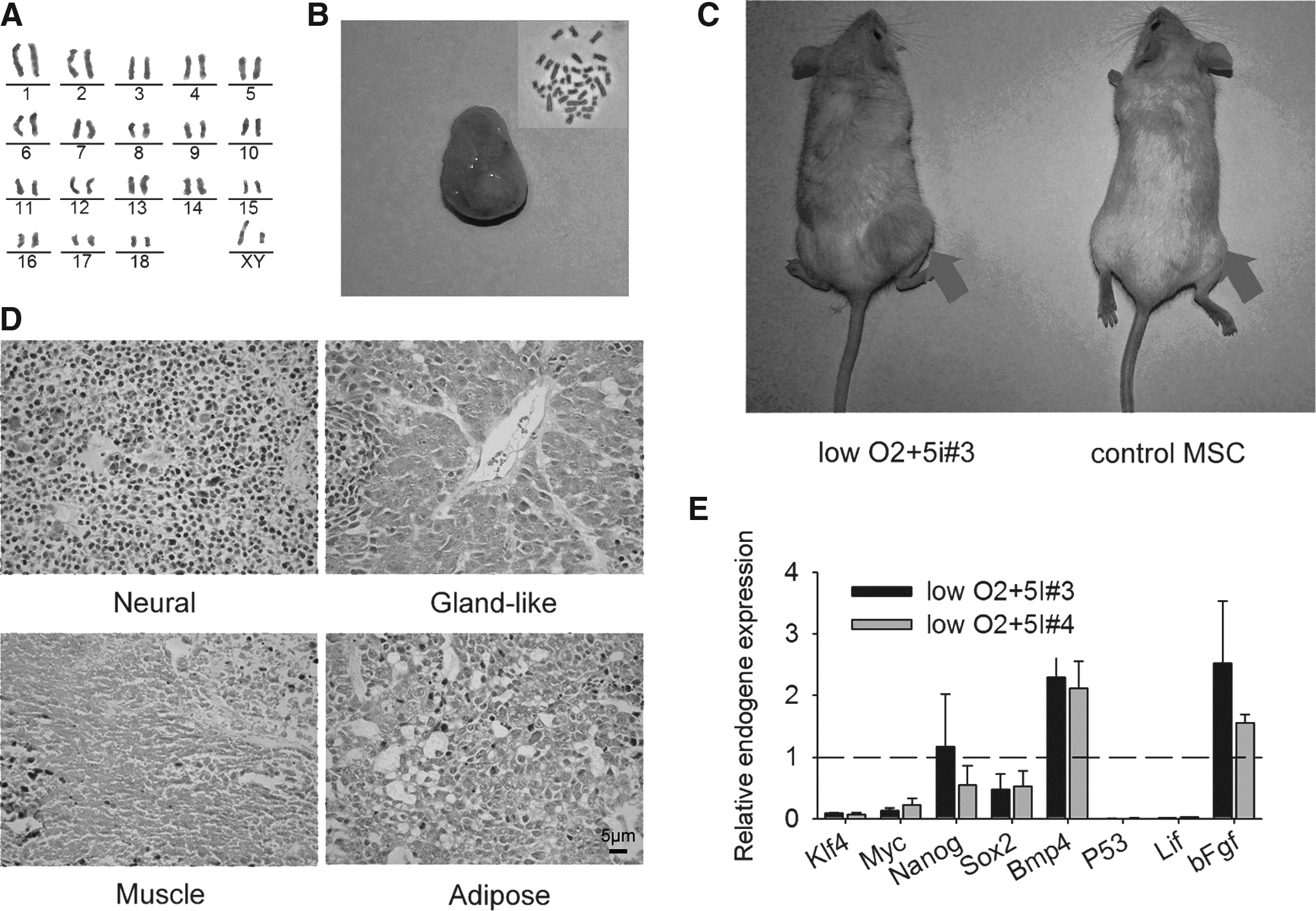
In vivo differentiation of porcine iPSCs. (
When the teratoma-forming cell lines were further analyzed, the expression levels of endo-Klf4, c-Myc, and Sox2 decreased in iPSCs when compared to their expression in MSCs (dashed line), whereas Nanog expression was stable. Furthermore, although P53 and Lif expression was nearly undetectable, the expression levels of Bmp4 and bFgf in the iPSCs were nearly twice as high as those in the MSCs (Fig. 4E).
Discussion
Obstacles facing the development of porcine iPSCs include lack of silencing of exogenes, failure in reactivation of endogenes, destruction of genomic integrity through genetic alterations caused by induction methods, and inability to maintain porcine iPSCs or ES cells in vitro. For instance, when exogenes are not successfully silenced, cells are prevented from undergoing differentiation, and donor cells are likely to form tumors (Miura et al., 2009; Takahashi et al., 2007). Also, porcine iPSCs have not gained the true pluripotency that can be achieved in mouse iPSCs when one or more pivotal pluripotent genes fail to reactivate, likely because the true pluripotent pathway in pig is currently unclear (Brevini et al., 2007; Hall, 2008). Finally, the optimal medium in which porcine pluripotent cells should be maintained remains to be determined. Despite these challenges, we successfully induced more than 10 stable porcine iPSCs lines, suggesting that low oxygen conditions and treatment with a small molecule cocktail can enhance reprogramming. The fact that low oxygen levels enhanced the reprogramming efficiency of iPSCs in our experiment is consistent with other studies (Yoshida et al., 2009). The low oxygen condition improves porcine MSC proliferation (Bosch et al., 2006). We split the MSC cells transfected with retroviruses on day 10 and subjected those cells to high and low oxygen culture conditions. After more than 10 days, low oxygen still generated more colonies than high oxygen did (data not shown).
The current major problem facing the generation of porcine iPSCs is the failure to silence exogenous transgenes. We hypothesized that this problem can be resolved by reducing the number of transcriptional factors used for porcine iPSCs induction. The endogenous pluripotency genes are not robustly activated in the two-factor pig iPSCs. Notably, endogenous Nanog and SSEA1 were highly expressed in the iPSCs than in MSCs by both immunostaining and flow cytometry, indicating that reprogramming does take place during porcine iPSCs induction. Compared with four or more factor-induced porcine iPSCs, the two-factor pig iPSCs also show differentiation ability because these cells efficiently form EBs and teratomas. We also employed traditional protocols (four factors) to induce iPSCs and achieved some porcine iPSCs colonies. Some of colonies show high reactivation of endogenous pluripotency genes including Nanog, Tert, and Terc; however, they failed to form EBs and teratomas. These data suggest that the current pluripotency-related genes commonly used for characterization of mouse and human pluripotent stem cells might not apply to mark pig pluripotent stem cells. We do not show these data in this report; they are included in another report in preparation.
We used a cocktail of small molecules when establishing iPSCs lines in this study. PD, CH, and SB are classic inhibitors that have previously been shown to improve the establishment of such cell lines (Lin et al., 2009; Ying et al., 2008), whereas NaB is a histone deacetylase inhibitor with global effects on epigenetic activation (Zhu et al., 2010). FK cooperates with PD and CH to convert epiblast stem cell (EpiSC)-like cells to ESC-like pluripotent stem cells (Hanna et al., 2010). We initially attempted to induce iPSC formation using this 5i small molecule cocktail alone or with Oct4, and no colonies formed under any conditions. Therefore, we introduced the porcine Klf4 factor along with a combination of inhibitors, considering that porcine iPSCs may resemble EpiSC-like cells (Hanna et al., 2009; Hanna et al., 2010; Zhou et al., 2010). Treatment with 5i did indeed improve both the establishment of cell lines with decreased exogenous gene expression and colony formation. While the silencing of exogenes and reactivation of endogenes is a hallmark of fully reprogrammed iPSCs (Takahashi et al., 2007), all of the iPSCs generated in this study expressed exogenes, albeit at various degrees. Because high exogene expression can have a negative impact on iPSC differentiation, we chose two iPSC lines with relatively low exogene expression for use in differentiation assays.
Testicular cells, MSCs, and iPSCs all expressed low levels of Sox2. A recent study has shown that Oct4 and Sox2, which maintain ESC identity, also orchestrate ESC fate decisions. Contrary to Oct4, Sox2 inhibits mesodermal differentiation and promotes neural ectodermal differentiation (Thomson et al., 2011).
The expression of P53 in porcine testis was much higher than that in MSCs and iPSCs. Because decreases in P53 expression significantly increase the reprogramming efficiency of both mouse and human cells (Kawamura et al., 2009), and allow cells to escape apoptosis (Fridman and Lowe, 2003), this may partially explain how it was possible to generate iPSCs from MSCs using only two factors.
The signaling mechanisms regulating pluripotency in porcine ESCs are currently obscure. Because bFGF is necessary for the maintenance of human ESCs and porcine iPSCs (Esteban et al., 2009; Ezashi et al., 2009; Telugu et al., 2010; Vallier et al., 2005), the high expression of bFgf in our iPSCs may account for their ability to grow without supplementation with bFGF, as well as their ability to maintain ESC-like morphology despite low Sox2 and Nanog expression. In the porcine embryo, low levels of Lif and wavy Bmp4 mRNA have been detected in the ICM, and no convincing evidence is currently available on Lif and Bmp4 expression in porcine ESCs (Blomberg et al., 2008; Hall et al., 2009). Our porcine iPSCs were found to express very low Lif levels and high levels of Bmp4.
Finally, our porcine iPSCs expressed SSEA-1, a specific surface marker similar to mouse ESCs, but low or no SSEA-4 and TRA-1-81, which are expressed by human ESCs. This is different from the porcine iPSCs described in previous reports that were induced by four human or mouse factors, processed human ESC-like colonies, and expressed SSEA-4 and TRA-1-81, but not SSEA-1 (Esteban et al., 2009; Montserrat et al., 2011; Wu et al., 2009); the porcine iPSCs induced by lentivirus-mediated transfection of four human factors with flat human ESC-like colonies that partially expressed SSEA1 and lacked SSEA4 and TRA-1-81 (Ezashi et al., 2009); or the porcine iPSCs induced by seven episomal human factors with mouse-ESC like colonies that were positive for both SSEA1 and SSEA4 (Telugu et al., 2010). Therefore, we are inspired to further optimize the factors and combinations of small molecules capable of inducing the generation of iPSCs. It is possible that a particular combination of such factors and small molecules will result in the generation of true pluripotent porcine iPSCs similar to mouse iPSCs. We anticipate that further improvements will be made to our protocol (e.g., by effectively increasing expression of Sox2 and Nanog), and that appropriate porcine factors in combination with small molecules will eventually induce the generation of authentic pluripotent porcine iPSCs capable of forming germ-line chimeras. This work represents the first important step toward the ultimate goal of reprogramming cells using only small molecules to achieve true pluripotent porcine iPSCs.
Footnotes
Acknowledgments
This work was supported by the MOST National Major Basic Research Program (2009CB941000 and 2011CBA01002). We thank Drs. Ning Li and Jiangyong Han at China Agricultural University for providing us with the pMXs-sus-Oct4 and pMXs-sus-Klf4 viral vectors. We thank Dr. Lingyi Chen for helpful discussions and Dr. Fang Wang and Xinglong Zhou for help with experiments.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.