
Abstract
Abstract
Previous studies have shown that the time interval between fusion and activation (FA interval) play an important role in nuclear remodeling and in vitro development of somatic cell nuclear transfer (SCNT) embryos. However, the effects of FA interval on the epigenetic reprogramming and in vivo developmental competence of SCNT embryos remain unknown. In the present study, the effects of different FA intervals (0 h, 2 h, and 4 h) on the epigenetic reprogramming and developmental competence of bovine SCNT embryos were assessed. The results demonstrated that H3 lysine 9 (H3K9ac) levels decreased rapidly after fusion in all three groups. H3K9ac was practically undetectable 2 h after fusion in the 2-h and 4-h FA interval groups. However, H3K9ac was still evidently detectable in the 0-h FA interval group. The H3K9ac levels increased 10 h after fusion in all three groups, but were higher in the 2-h and 4-h FA interval groups than that in the 0-h FA interval group. The methylation levels of the satellite I region in day-7 blastocysts derived from the 2-h or 4-h FA interval groups was similar to that of in vitro fertilization blastocysts and is significantly lower than that of the 0-h FA interval group. SCNT embryos derived from 2-h FA interval group showed higher developmental competence than those from the 0-h and 4-h FA interval groups in terms of cleavage rate, blastocyst formation rate, apoptosis index, and pregnancy and calving rates. Hence, the FA interval is an important factor influencing the epigenetic reprogramming and developmental competence of bovine SCNT embryos.
Introduction
For the success of SCNT, epigenetic modifications of highly differentiated somatic cells should be completely erased, and new epigenetic modifications similar to that of embryonic cells should be reestablished at the appropriate time (Rideout et al., 2001). This process, known as nuclear reprogramming, mainly involves various epigenetic modifications, such as DNA methylation and histone acetylation (Hochedlinger and Jaenisch, 2006). Numerous experiments have shown that epigenetic reprogramming after SCNT is incomplete and improper. Aberrant DNA methylation and histone acetylation at the global genome level and at some developmentally important single-gene sites have been observed in most SCNT embryos at different developmental stages (Dean et al., 2001; Nolen et al., 2005; Rybouchkin et al., 2006). Aberrant epigenetic modifications have been considered to be the main cause of developmental abnormalities observed in cloned embryos or offspring.
Previous studies have indicated that the time interval between fusion and activation (FA interval) has great impact on the nuclear remodeling and developmental potential of SCNT embryos (Aston et al., 2006; Yin et al., 2003). However, the conclusions from different research groups vary. Several studies have indicated that prolonged exposure of the donor nuclei to the inactivated metaphase II (MII) oocytes containing high activity of maturation-promoting factor (MPF), and mitogen-activated protein kinase is beneficial for nuclear remodeling, as evidenced by increased proportions of nuclear envelope breakdown (NEBD) and premature chromosome condensation (PCC) of somatic donor nuclei in reconstructed oocytes (Lee and Campbell, 2006). The accessibility of reprogramming factors is perceived to increase if the donor nuclei have undergone NEBD and PCC (Campbell and Alberio 2003). Several studies have suggested that the occurrence of PCC is not essential for the effective reprogramming of the incoming nuclei in the reconstructed oocytes (Sung et al., 2007). Moreover, other studies have shown that prolonged exposure of donor nuclei to inactivated recipient ooplasts impairs the developmental potential of SCNT embryos by inducing chromosome scattering and nuclear aneuploidy (You et al., 2010). Thus, the impact of the FA interval on the developmental competence of SCNT embryo remains controversial. The examination of additional parameters on the effects of the FA interval on the reprogramming and developmental competence of SCNT embryos may contribute to the understanding of this process.
Accordingly, this study investigated the effects of the FA interval on epigenetic reprogramming and developmental competence of SCNT embryos. The influence of the FA interval on the epigenetic reprogramming of SCNT embryo was assessed in the examination of the DNA methylation level of satellite I at the blastocyst stage and the dynamic change in the histone acetylation level of H3 lysine 9 (H3K9ac) at various developmental stages of SCNT embryos. The developmental competence of SCNT embryos was comprehensively evaluated in terms of cleavage rate, blastocyst formation rate, apoptosis index of day-7 blastocysts, and pregnancy and calving rates.
Materials and Methods
Unless otherwise noted, all chemicals and reagents were purchased from the Sigma-Aldrich Company (St. Louis, MO, USA). Plasticware was purchased from Nunclon (Roskilde, Denmark). The entire experimental procedure was approved by the Animal Care and Use Committee of Northwest A&F University and performed in accordance with the animal welfare and ethics guidelines.
Preparation of nuclear donor cells
Primary neonatal fibroblasts were isolated from the ear skin of a newborn Holstein heifer and cultivated in Dulbecco's modified Eagle medium (DMEM; Gibco, Grand Island, NY, USA) containing 15% fetal bovine serum (FBS; Gibco, Grand Island, NY, USA), 1 mM sodium pyruvate, 100 IU/mL penicillin, and 100 mg/mL streptomycin. The cells were trypsinized when 80% confluence was reached and seeded into three new 60-mm dishes for further passaging. Donor cells for SCNT were derived from generations 2–5 and induced to the G0/G1 stage of the cell cycle by serum starvation for 2–3 days. A single-cell suspension was prepared by trypsinization of cultured cells and resuspension in phosphate-buffered saline (PBS) containing 10% FBS and 7.5 μg/mL cytochalasin B (CB) before nuclear transfer.
Oocyte recovery and in vitro maturation
Cow ovaries obtained from a local slaughterhouse were transported to the laboratory within 3–4 h at 15–20°C in sterile physiological saline containing 100 IU/mL penicillin and 100 mg/mL streptomycin (Wang et al., 2011c). The ovaries were washed three times in sterile physiological saline. Cumulus–oocyte complexes (COCs) were obtained by aspirating 2-mm to 10-mm follicles with a 12-gauge needle attached to a 10-mL syringe, and then recovered in PBS containing 3% (vol/vol) FBS. Only COCs with a minimum of three compact cumulus cell layers and homogeneous cytoplasm were selected and washed in PBS (vol/vol) supplemented with 3% FBS. Groups of up to 50 COCs were placed in 2 mL of maturation medium in four-well dishes and cultured for 18–20 h at 38.5°C in 5% CO2 humidified air atmosphere. The maturation medium used was the bicarbonate-buffered tissue culture medium-199 (TCM-199, Gibco, BRL, Grand Island, NY, USA) supplemented with 10% (vol/vol) FBS, 50 ng/mL epidermal growth factor, 1 μg/mL 17-estradiol, and 0.075 IU/mL human menopausal gonadotropin.
Nuclear transfer and activation
SCNT was performed as previously described (Wang et al., 2011c). Briefly, at 18–20 h after the onset of oocyte maturation, cumulus cells were completely removed from COCs by repeated pipetting in PBS containing 0.2% bovine testicular hyaluronidase. Denuded oocytes with an extruding first polar body and evenly granulated ooplasm were selected and cultured in PBS supplemented with Hoechst 33342 (10 μg/mL) for 15 min. After incubation, the oocytes were transferred into a manipulation medium (PBS containing 10% FBS and 7.5 μg/mL CB) in microdrops overlaid with mineral oil. The first polar body and a small amount of adjacent cytoplasm (about 1/4 cytoplasm) were aspirated with a glass pipette (inner diameter, 20 μm) under an inverted microscope equipped with epifluorescence. The expelled cytoplasm was examined under ultraviolet radiation to confirm the complete removal of the nuclear material, and oocytes harboring residual DNA were discarded. After enucleation, a single trypsinized donor cell was inserted into the perivitelline space of the enucleated oocytes. The oocyte–cell couplet was inserted between a pair of platinum electrodes connected to the micromanipulator in microdrops of 280 mM mannitol containing 0.001 mM CaCl2 and 0.05 mM MgCl2, and a double electrical pulse of 34 V for 10 μsec was applied for oocyte–cell fusion (Liu et al., 2007). The successful fusion of oocyte–cell couplets were examined after 30 min, and this time point was defined as 0-h FA interval. Based on the experimental design, successfully reconstructed embryos were randomly assigned to the 0-h, 2-h, and 4-h FA interval groups, and then activated by exposure to 5 mM ionomycin for 4 min, followed by 5 h of incubation in modified synthetic oviduct fluid (mSOF) containing 10 mg/mL cycloheximide and 5 μg/mL CB.
Culture of SCNT embryos
SCNT embryos were cultured in G1.2/G2.2 sequential media as previously described (Wang et al., 2011b). Briefly, embryos were first cultured in droplets (10 embryos/50 μL) of G1.2 covered with mineral oil in a humidified atmosphere with 5% CO2 in air at 38.5°C for 72 h. Embryos were washed three times in G2.2 and cultured for another 96 h in 50 μL of G2.2. Subsequent in vitro development in the two-cell and blastocyst stages was monitored at 24, 48, 144, and 168 h of culture (0 h indicates the time at which embryos were transferred into G1.2). G1.2/G2.2 sequential media were purchased from Vitrolife AB (Gothenburg, Sweden), both containing 5 mg/mL of human serum albumin (Gothenburg, Sweden).
Immunofluorescence Staining of H3K9ac
For immunodetection of H3K9ac, SCNT embryos from different FA interval groups were washed three times (5 min per wash) in PBS containing 0.2% polyvinyl alcohol (PVA), and fixed at room temperature in 4% (wt/vol) paraformaldehyde in PBS at different time points after fusion (2, 6, 10, and 30 h). The embryos were then permeabilized by 0.5% Triton X-100 for 8 min. After washing three times in PBS, the embryos were blocked in the Immunol Staining Blocking Solution (Beyotime, P0102) overnight at 4°C. Afterward, the embryos were exposed to primary antibodies (rabbit polyclonal antibody against H3K9ac, Abcam, Hong Kong, diluted to 1:500) overnight at 4°C. After washing three times, the embryos were incubated for 2 h at room temperature in the presence of 1:500 diluted secondary antibodies (goat anti-rabbit IgG, Beyotime, Nantong, China). The embryos were rinsed three times for 5 min each in 0.2% PVA-PBS and stained with 4′,6-diamidino-2-phenylindole (DAPI) (Beyotime, C1005) for 3 min. Finally, the embryos were mounted on glass slides with a drop of antifade mounting medium (Beyotime, P0126) and analyzed using a Nikon Eclipse Ti-S microscope equipped with a 198 Nikon DS-Ri1 digital camera (Nikon, Tokyo, Japan). The mean values of the fluorescence intensities were measured on the H3K9 and DNA channels with Image J Software (NIH, Bethesda, MD, USA) by selecting the area of the individual nuclei in SCNT embryos for every time point and FA interval group. The average profiles, represented as the ratio of H3K9ac to DNA signals, were used to compare between different groups.
Bisulfite sequencing analysis
A single high-quality day-7 blastocyst (having an evident blastocoel cavity) was used per sample, and 5, 9, and 6 embryo samples were used for the 0-h, 2-h, and 4-h FA intervals, respectively. Six embryo samples were used for the IVF groups. Genomic DNA from the embryo was extracted and subjected to sodium bisulfite treatment using the EZ DNA Methylation-Direct™ Kit (Zymo Research, USA) in accordance with the instruction manual with minor modifications. Briefly, each blastocyst was rinsed twice with PBS-PVA and transferred to 20 μL of digestion mixture containing 10 μL of M-digestion Buffer (2×), 9 μL of dH2O, and 1 μL of Proteinase K. After incubation at 50°C for 3 h, the sample was added to 130 μL of CT conversion reagent in a polymerase chain reaction (PCR) tube, denatured at 98°C for 8 min, and underwent bisulfite conversion at 64°C for 3.5 h. DNA was then desalted, purified, and eluted with 15 μL of elution buffer. Bisulfite sequencing PCR (BS-PCR) was immediately performed using 2 μL of modified DNA per PCR run. The reaction mixture (50 μL) contained 25 μL of Zymo Taq™ premix, 2 μL of modified DNA, 21 μL of dH2O, and 1 μL of both forward and reverse primers (Table 1). The PCRs were performed with the following program: 95°C for 4 min, followed by 40 cycles of denaturation at 95°C for 30 sec, annealing at 46°C for 30 sec, extension at 72°C for 20 sec, and final extension at 72°C for 7 min. To confirm the specific amplification, the PCR products were resolved on 2.5% agarose gel and gel-purified using the TIANgel Midi Purification Kit (Tiangen, China). Purified fragments were subcloned into pMD18-T vectors (TaKaRa, Japan) and selected for DNA sequencing (BGI, China). Three independent amplification experiments were performed for each sample. Three to four clones from each independent set of amplification and cloning were sequenced, such that a total of 9–12 clones are available in each sample. Bisulfite sequencing data and C-T conversion rates were analyzed by BIQ Analyzer software (Bock et al. 2005). To ensure high data quality, sequences that had C-T conversion rates below 95% were excluded. Methylation data from bisulfite sequencing was analyzed by computing the percentage of methylated CpGs from the total number of CpGs.
Annealing temperature.
Forward primer.
Reverse primer.
Embryo transfer and pregnancy diagnosis
Beef-breed Angus cows between 2–5 years of age was used as recipient animals. They were certified free from all major infectious cattle diseases by repeated serological testing and randomly assigned to receive embryos. Estrous cycles were synchronized using a progestagen implant for 9 days combined with a prostaglandin analog injection (2 mL of Estrumate) 2 days before implant removal. After estrus detection, day-7 recipients carrying palpable corpus luteum (>0.8 cm) were selected for embryo transfer. Two high-quality blastocysts were loaded into 0.25-mL straws in HEPES-buffered TCM-199 supplemented with 10% FBS, and transported at 37°C from the laboratory to the experimental farm within 30 min. The embryo was nonsurgically transferred from the uterine horn ipsilateral to the corpus luteum. Pregnancy was detected by rectal palpation on the 90th day of gestation (day 0 being the day embryos were transferred into recipients).
Statistical analysis
All statistical analyses were performed with SPSS version 13.0 software (SPSS, Inc., Chicago, IL). Data on the development of SCNT embryos and H3K9ac fluorescence intensity ratios were analyzed with a chi-squared test. Differences were considered significant when p<0.05. Data on the mean number of cells in the blastocysts and the DNA methylation level of satellite I were analyzed by one-way ANOVA and compared using Tukey–Kramer test. Differences were considered significant when p<0.05.
Results
Effects of the FA interval on dynamic reprogramming of histone acetylation levels in SCNT embryos
Representative acetylated H3K9 immunofluorescence images of SCNT embryos derived from different FA interval groups are illustrated in Fig. 1. The mean relative levels of histone acetylation signal intensities in the groups are shown in Table 2. Low histone acetylation levels were found in all three groups 2 h after fusion and were practically undetectable in the 2-h and 4-h FA interval groups. However, 6 h after fusion, the histone acetylation level increased in all three groups, but the acetylation levels of H3K9 in the 2-h and 4-h FA interval groups were higher than that in the 0-h FA interval group. A similar phenomenon was also observed 10 h after fusion. At the two-cell developmental stage (30 h after fusion), no significant difference was observed in the H3K9 acetylation levels among the three FA interval groups.
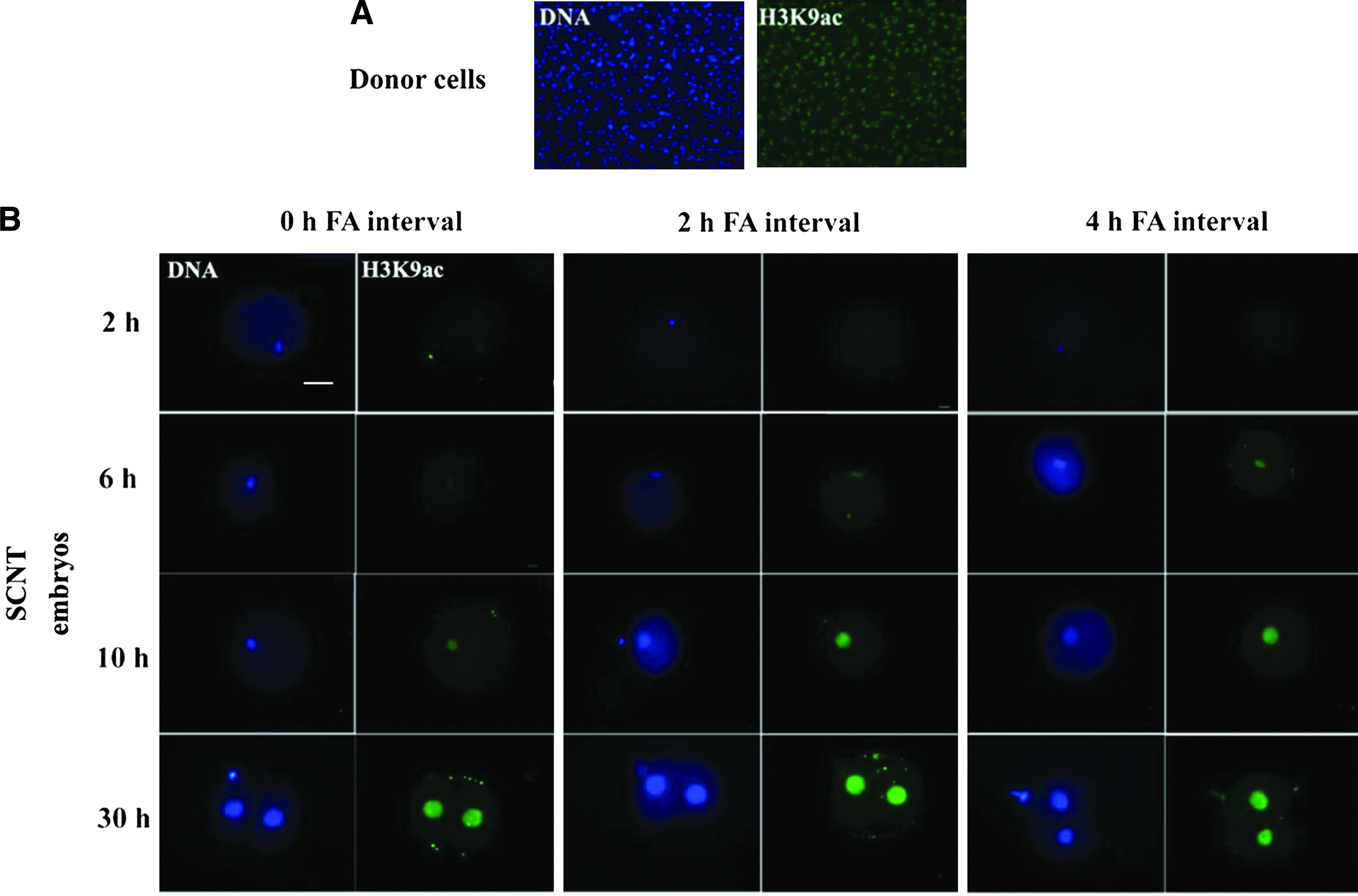
Global acetylation levels of histone H3 lysine 9 (H3K9ac). (
0* is acetylation level of H3K9 (mean±standard error of the mean [SEM]) in nuclear donor somatic cells prior to SCNT
Values with different superscripts (a and b) differ significantly (p<0.05).
FA, interval time between fusion and activation; SCNT, somatic cell nuclear transfer.
Effects of FA interval on DNA methylation status of the satellite I sequence
The effects of FA interval on the DNA methylation status of the satellite I sequence of day-7 blastocysts were analyzed by bisulfite sequencing. As shown in Figure 2, the DNA methylation level of the satellite I sequence was significantly higher in the day-7 blastocysts derived from the 0-h FA interval group than those from the 2-h and 4-h FA interval group as well as that from the control (IVF) (54.1±5.9 vs. 31.4±3.6, 31.7±5.6, or 13.5±5.9, respectively; p<0.05).
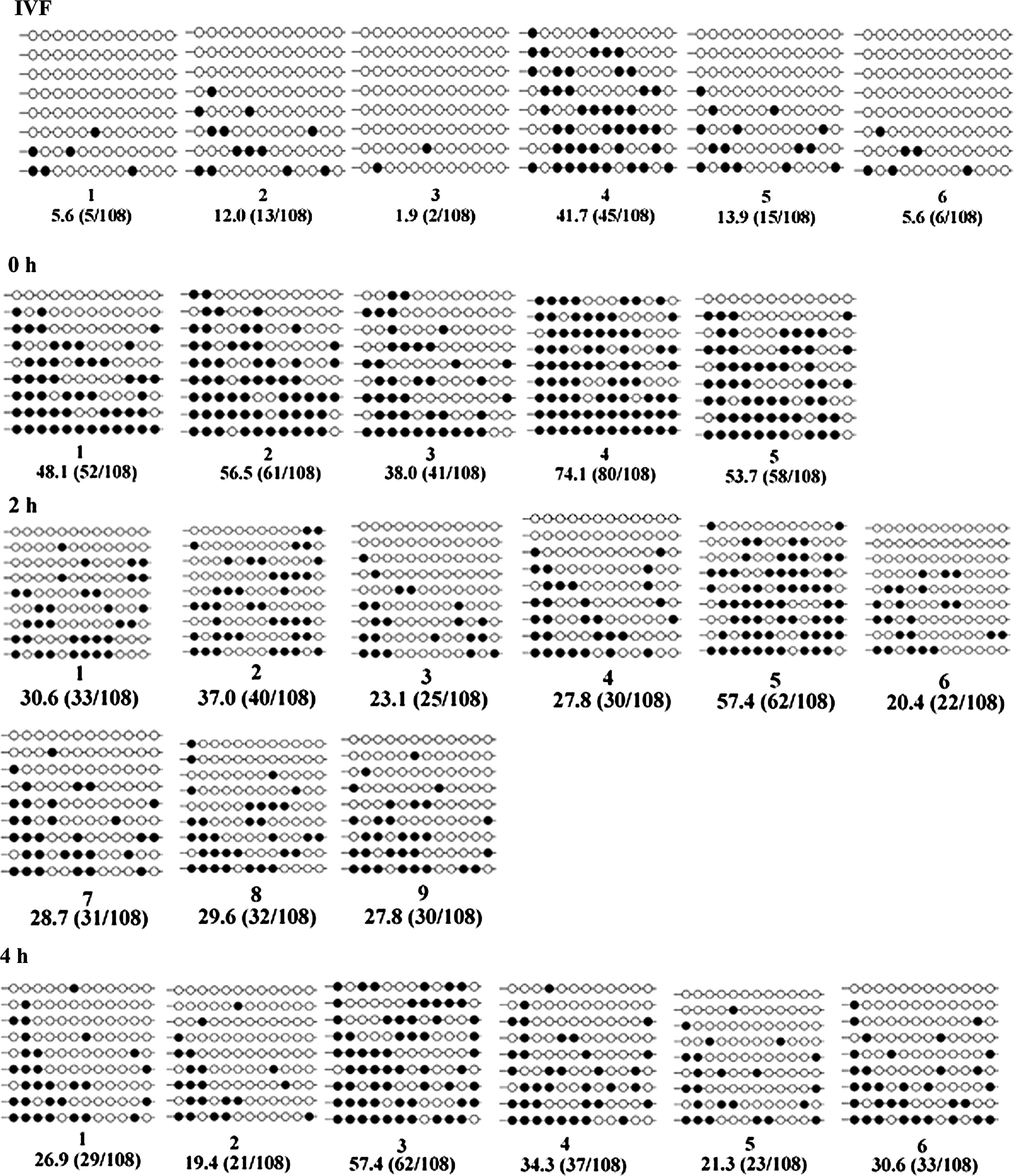
Methylation profiles of 12 CpGs in the satellite I region analyzed by bisulfite sequencing. Unfilled (white) and filled (black) circles represent unmethylated and methylated CpGs, respectively. Horizontal lines of circles represent one separate clone that was sequenced (5–9 for each sample). For each sample, the methylation data were analyzed by computing the percentage of methylated CpGs from the total number of CpGs.
Effect of the FA interval on the in vitro developmental competence of bovine SCNT embryos
Although no statistical difference was observed in the cleavage rate among the three FA interval groups after 48 h of culture (75.5%, 79.4%, and 83.7% for 0-h, 2-h, and 4-h FA interval groups, respectively), the cleavage rate after 24 h of culture was significantly higher in the 2-h and 4-h FA interval groups than that in the 0-h FA interval group (60.0% or 62.7% vs. 42.3%, p<0.05), as seen in Table 3. Blastocyst formation rate was significantly higher in the 2-h FA interval group compared with the 0-h and 4-h FA interval groups on both days 6 and 7 of culture (day 6, 30.3% vs. 16.4% and 18.0%; day 7, 42.4% vs. 28.3% and 27.1%, respectively).
The cleavage and blastocyst rates are based on the number of cultured embryos.
Values with different superscripts differ significantly (p<0.05).
FA, interval time between fusion and activation.
Effect of the FA interval on the frequency of apoptotic nuclei in SCNT blastocysts
The apoptosis index of day-7 blastocysts was analyzed to determine the impact of the FA interval on the quality of bovine SCNT embryos. As shown in Table 4 and Figure 3, the incidence of apoptosis in day-7 SCNT blastocysts was significantly higher in the 4-h interval group than those in the 0-h and 2-h FA interval groups (15.9±2.4 vs. 5.8±1.6 and 5.6±2.1, P<0.05). In addition, the total cell number in day-7 blastocysts was significantly higher in the 2-h FA interval group than those the in 0-h and 4-h FA interval groups (100.2±7.1 vs. 83.6±8.2, and 85.4±10.8, p<0.05).
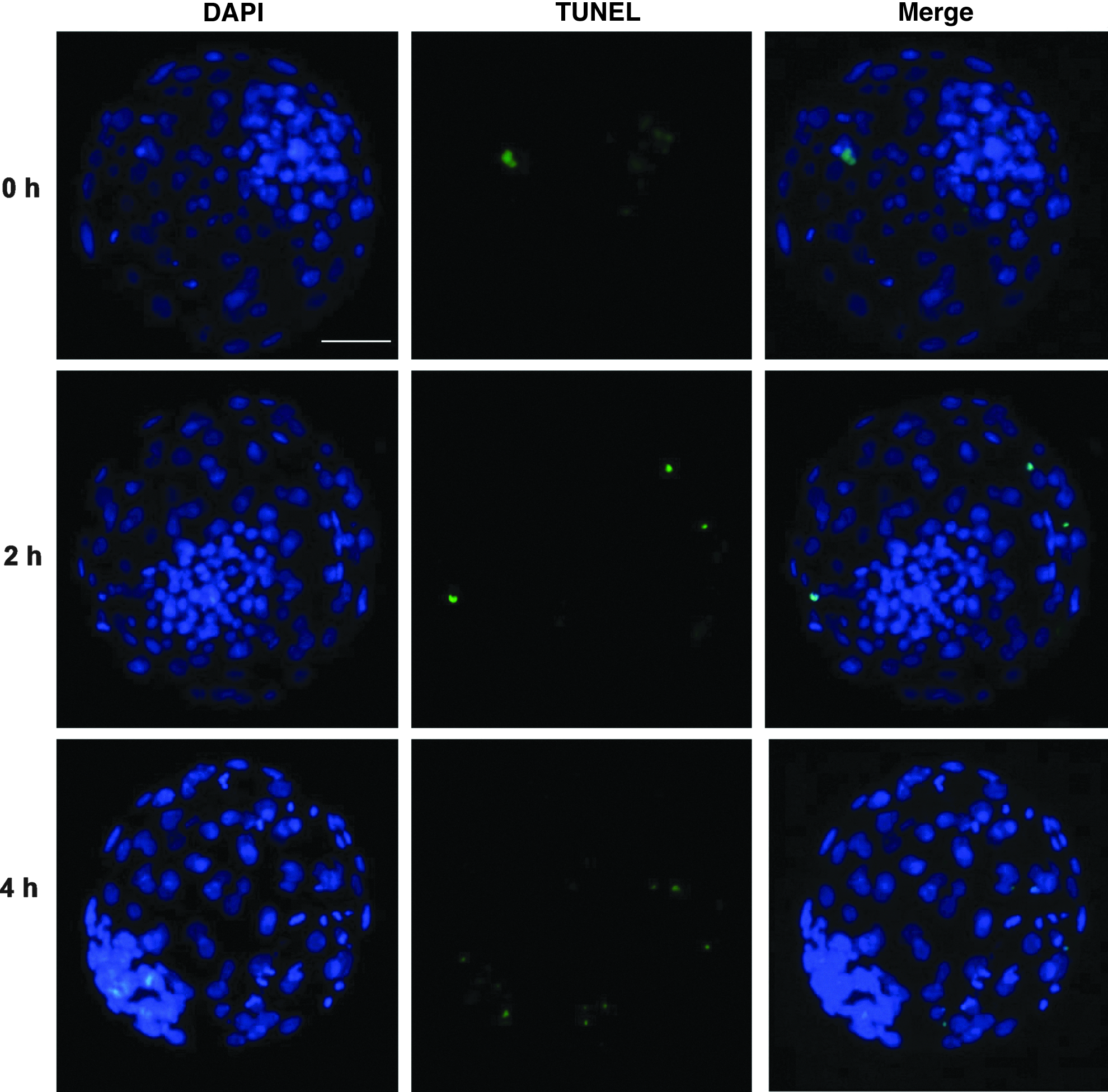
Detection of apoptotic nuclei in day 7 blastocysts derived from 0-h, 2-h, and 4-h FA interval groups. Blastocysts were fixed, permeabilized, and treated using terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL). The cell nuclei were stained with DAP1 whereas the apoptotic cells were labeled by TUNEL. Original magnification, 100×. Scale bar, 50 μm. Color images available online at www.liebertpub.com/cell
Within the same column, values with different superscripts differ significantly (p<0.05).
Control is the day-7 blastocysts derived from in vitro fertilization.
FA, interval time between fusion and activation; SCNT, somatic cell nuclear transfer; SEM, standard error of the mean.
Effect of the FA interval on in vivo development of bovine SCNT embryos
As shown in Table 5, a total of 64, 98, and 60 blastocysts derived from 0-h, 2-h, and 4-h FA interval groups, respectively, were transferred to the recipient Angus cows (2 blastocysts/recipient). The pregnancy rate was significantly higher in the 2-h FA interval group than those in the 0-h and 4-h FA interval groups (27.6% vs. 9.4% and 6.7%, p<0.05). Birth rates or cloning efficiency was higher in the 2-h FA interval group than that in the 0-h and 4-h FA interval groups. However, the difference was not statistically significant.
Day-7 blastocysts were nonsurgically transferred (2 embryos per recipient) to synchronized recipient cows.
Pregnancy rate was detected by rectal palpation on day 60 of gestation.
Cloning efficiency is the number of surviving calves at 60 days after birth/number of recipient cows.
Values with different superscripts within columns are significantly different from each other (p<0.05).
FA, interval time between fusion and activation; SCNT, somatic cell nuclear transfer.
Discussion
For the successful development of SCNT embryos, a highly differentiated somatic cell nucleus should be reprogrammed to a totipotent state within a short duration (usually prior to cleavage) by some unknown factors in the ooplasts. However, this nuclear reprogramming process is inefficient and various reprogramming abnormalities, such as inappropriate histone acetylation and DNA methylation, have been observed in most SCNT embryos, that have been considered as the primary reasons for the low efficiency of SCNT. In the present study, the effects of various FA intervals on the epigenetic reprogramming of bovine SCNT embryos were evaluated. Our observations show that the FA interval has considerable effect on the epigenetic reprogramming of bovine SCNT embryos. Holding the somatic nuclei in the inactivated ooplasm for 2 h is beneficial for the nuclear reprogramming and developmental competence of SCNT embryos in bovine.
For successful cloning, donor nuclei are routinely exposed for a certain period in the enucleated MII oocytes to allow extensive nuclear–oocyte interaction and stimulate reprogramming of donor nuclei before parthenogenetic activation (Rybouchkin et al., 2006). The mechanism for the molecular basis of nuclear–oocyte interaction and nuclear reprogramming in MII oocytes after SCNT remains unknown and appears to be highly complex. Several important reprogramming factors are believed to be specifically present in MII oocytes, and the reprogramming factors become inactive after oocyte activation (Tani et al., 2007). Therefore, the series of cellular events that occurs in the inactive MII oocytes after SCNT should be explored.
In the present study, insufficient epigenetic reprogramming of the introduced somatic cell nuclei and significantly lower developmental competence were observed in the 0-h FA interval group than in the 2-h exposure. The results evidently indicated that simultaneous fusion and activation could be insufficient for reprogramming the somatic cell nuclei. This finding agrees with the studies of Kishigami et al. (2006) and Rybouchkin et al. (2006), which reported that SCNT oocytes should be routinely incubated in enucleated MII oocytes for at least 40 min, and that 15 min of incubation before activation could be insufficient for reprogramming the introduced somatic cell nuclei (Kishigami et al., 2006; Rybouchkin et al., 2006). The donor nuclear membrane might not have broken down after the short exposure (less than 30 min) to MII oocyte. In this case, donor cell chromosomes could have not been directly exposed to the reprogramming factors in the oocyte cytoplasm.
Meanwhile, we demonstrated that the prolonged exposure (up to 4 h) of a donor nucleus to the preactivation MII oocyte was also not beneficial for the development of cloned embryos. The blastocyst developmental and pregnancy rates were significantly lower than those in 2-h FA interval group. Moreover, blastocysts derived from the 4-h FA interval group showed higher apoptosis index than those from the 0-h and 2-h groups. Several studies have shown that prolonged exposure (up to 4 h) of a donor nucleus to the preactivation MII oocyte containing high MPF activity may lead to abnormal chromatin structure, possibly causing a range of DNA damage (e.g., fragmented chromatin, joined chromatin, and chromosomal breakage) or loss of chromosomes (You et al., 2010). In addition, anomalous pronuclei may form, resulting in numeric chromosome errors, such as polyploidy and mixoploidy (Choi et al., 2004). These phenomena could elucidate the inferior in vitro development of NT embryos after prolonged exposure of the donor nucleus to the preactivation MII oocyte. However, Wells et al. reported a comparable blastocyst developmental rate when the FA interval was prolonged from 2 h to 6 h (Wells et al., 1999). The discrepancies in the results from various studies might be explained by the differences in age of oocyte recipients, selection of somatic donor cells, protocols used, and experimental conditions (Akagi et al., 2003; Loi et al., 1998).
For NT to be successful, the donor nucleus must be properly reprogrammed, wherein epigenetic marks are erased from the donor nucleus genome, resulting in the erasure of tissue-specific gene expression patterns and effectively resetting the cell to a totipotent state. Reprogramming in SCNT embryos is believed to occur at an epigenetic level, and, as one of the epigenetic modifications related to chromatin structure, the global level of histone acetylation (Yang et al. 2007). In particular, H3K9ac plays an important role in mediating nucleosome remodeling, facilitating the access of the transcriptional machinery to nucleosomal DNA throughout the embryonic stages (MacDonald and Howe, 2009).
The acetylation/deacetylation dynamics of H3K9ac have been analyzed in several studies involving SCNT embryos (Rybouchkin et al., 2006; Wang et al., 2007). These studies indicated that the dynamics of H3K9ac could be used as a good marker for the extent of nuclear reprogramming. Therefore, in the present study, we selected the H3K9ac as a marker to study the effect of FA interval on the extent of nuclear reprogramming in SCNT embryos. The results indicated that the FA interval has a great impact on nuclear reprogramming after SCNT. The dynamics of H3K9ac were very similar between the 2-h and 4-h FA interval groups, but different from that in the 0-h FA interval group. The histone H3K9ac levels rapidly decreased after fusion in all three groups. H3K9ac was practically undetectable 2 h after fusion in both 2-h and 4-h FA interval groups. Similar H3K9ac dynamics have been described by another study for murine cloned embryos activated after a 2-h FA interval (Wang et al., 2007). However, the H3K9ac/DNA ratio only showed 70% decrease in the 0-h FA interval group, which was significantly higher than those in the 2-h and 4-h FA interval groups (p<0.05). The levels of histone acetylation increased 6 h and 10 h after fusion in all three groups. However, the H3K9ac levels in the 2-h and 4-h FA interval groups were higher than that in the 0-h FA interval group. No significant difference was observed in the acetylation level of H3K9 among the three FA interval groups at the two-cell developmental stage.
Histone deacetylation was suggested to allow the silencing of the somatic gene expression program prior to the initiation of the embryonic pattern of gene expression. Therefore, a complete histone deacetylation before the beginning of the reacetylation may be necessary for the correct regulation of gene expression and development of SCNT embryos. After histone deacetylation, higher histone acetylation level is correlated with the transcriptional permissive state of the chromatin, which is believed to be necessary for the activation of embryonic genes during nucleus reprogramming. In this sense, the histone acetylation dynamics observed in the SCNT embryos derived from the 2-h and 4-h FA interval groups might indicate the occurrence of more sufficient nuclear reprogramming than that observed in 0-h FA interval group. Additionally, the greater variation of histone deacetylation in different nuclei in the three FA interval groups may indicate that nuclear reprogramming is incomplete in other portions of the cells in the SCNT embryos.
In addition to histone acetylation, DNA methylation is another key epigenetic modification that regulates the chromatin structure and plays a crucial role in sustaining genomic stability, activating or suppressing gene expression, regulating X-chromosome inactivation, maintaining genomic imprinting, and silencing repetitive elements (Dean et al., 2001; Lan et al., 2010). On the basis of previous studies, DNA demethylation and remethylation events are not always faithfully recapitulated in the SCNT embryos (Cheng et al., 2011; Chung et al., 2003; Dean et al., 2001; Kang et al., 2002). Methylation status in the satellite I region has been widely used as a marker for investigating the epigenetic reprogramming level of SCNT embryos, and aberrant hypermethylation status in the satellite I region was observed in most bovine SCNT blastocysts (Kang et al., 2001; Su et al., 2011; Wang et al., 2011a).
The results of the present study demonstrated that the DNA methylation level in satellite I was significantly higher in day-7 cloned blastocysts from the 0-h FA interval group than those from the 2-h and 4-h FA interval groups as well as that of the control (IVF) groups. DNA methylation has been shown to suppress gene expression. Thus, hypermethylation may also cause improper expression of developmentally important genes, which in turn leads to the developmental failure of the SCNT embryo. The present study indicated that the aberrant hypermethylation status in the satellite I region can be alleviated by optimizing the SCNT technique. Prolonged exposure (up to 2 h) of a donor nucleus to the preactivation MII oocyte could reduce the level of DNA methylation of the satellite I region, and the DNA methylation level was much closer to that of the IVF counterpart compared to the 0-h FA interval group.
In conclusion, the results demonstrated that the FA interval is an important factor influencing the epigenetic reprogramming and developmental competence of bovine SCNT embryos. Holding the somatic nuclei in inactivated MII oocytes for 2 h is beneficial for the nuclear reprogramming and developmental competence of bovine SCNT embryos.
Footnotes
Acknowledgments
This work was supported by grants from the National Key Project for Production of Transgenic Livestock, China (No. 2008ZX-08007-004).
The authors thank Wang You Nan for providing the cow ovaries used in this study, Dr. Shan Ling for her critical correction of the revised manuscript, Guoliang Pei for the assistance with the immunofluorescence staining and observation, and Zhao Liang for embryo transfer and care for recipient cows.
Author Disclosure Statement
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the reported research.