
Abstract
Abstract
The shortage of human organs and tissues for transplant has led to significant interest in xenotransplantation of pig tissues for human patients. However, transplantation of pig organs results in an acute immune rejection, leading to death of the organ within minutes. The α-1,3-galactosyltransferase (GALT) gene has been knocked out in pigs to reduce rejection, yet additional genes need to be modified to ultimately make pig tissue immunocompatible with humans. The development of pig induced pluripotent stem cells (piPSCs) from GALT knockout (GALT-KO) tissue would provide an excellent cell source for complex genetic manipulations (e.g., gene targeting) that often require highly robust and proliferative cells. In this report, we generated GALT-KO piPSCs by the overexpression of POU5F1, SOX2, NANOG, LIN28, KLF-4, and C-MYC reprogramming genes. piPSCs showed classical stem cell morphology and characteristics, expressing integrated reprogramming genes in addition to the pluripotent markers AP, SSEA1, and SSEA4. GALT-KO piPSCs were highly proliferative and possessed doubling times and telomerase activity similar to human embryonic stem cells. These results demonstrated successful reprogramming of GALT-KO fibroblasts into GALT-KO piPSCs. GALT-KO piPSCs are potentially an excellent immortal cell source for the generation of pigs with complex genetic modifications for xenotransplantation, somatic cell nuclear transfer, or chimera formation.
Introduction
Pig xenografts in nonhuman primates are rapidly rejected by hyperacute rejection (HAR) and acute humoral xenograft rejection (AHXR) responses (Hisashi et al., 2008; McGregor et al., 2012; Yamada et al., 2005). These responses are triggered by antibodies against the galactose α-1,3-galactose (Gal) epitope on porcine vascular endothelial cells present in the tissue. The subsequent immune response results in vascular damage leading to hemorrhaging, edema, and necrosis in the graft. To prevent this response, the α-1,3-galactosyltransferase (GALT) gene was knocked out in pigs using a gene targeting and somatic cell nuclear transfer (SCNT) approach, resulting in a complete loss of the Gal epitope in tissue (Lai et al., 2002; Phelps et al., 2003; Sharma et al., 2003). However, knockout (KO) of this gene has proven to be insufficient to completely attenuate immune rejection. Additional genes have now been identified that could be knocked out such as CD86, which activate human T and natural killer (NK) cells, or genes could be introduced such as human complement regulatory proteins CD55 and CD59, which both block HAR (Byrne et al., 1997; Costa et al., 2004; McGregor et al., 2012). The identification of other genes that are capable of reducing the immune response has led to interest in creating genetically engineered pigs that carry multiple gene deletions and additions. However, the current state of the art in transgenic pig engineering has been a significant hurdle in creating the needed animals.
One of the major challenges of producing genetically engineered pigs for xenotransplantion is the lack of highly proliferative, immortal pig cells similar to pluripotent stem cells. Pig pluripotent stem cells would allow for multiple rounds of genetic manipulation and screening to produce cells with the necessary mutations for generating immune-tolerant cells. A recent breakthrough in the production of pluripotent stem cells using iPSC technology has now made it possible to create highly proliferative stem cells from pigs (Takahashi and Yamanaka, 2006; West et al., 2010b; West et al., 2011). A number of groups, including our own, have recently shown that iPSC technology can be adapted for use in pigs (Esteban et al., 2009; Ezashi et al., 2009; West et al., 2010b; West et al., 2011; Wu et al., 2009). piPSCs are highly proliferative and express high levels of telomerase, key characteristics needed for multiple rounds of gene targeting and selection. Recently, it was demonstrated that piPSCs introduced into developing embryos can successfully incorporate and form chimeric animals and transmit genes through the germ line (West et al., 2010b; West et al., 2011). The ability to produce chimeric animals offers an alternative to create transgenic pigs without the use of SCNT. These cells present a unique opportunity to create pigs for xenotransplantion. However, it has never been shown that piPSCs can be produced from a complex genetically manipulated background such as GALT-KO.
In this study, we demonstrated that GALT-KO pig fibroblast cells can be successfully reprogrammed into piPSCs. GALT-KO piPSCs showed morphological features and expressed a number of key pluripotent genes and proteins indicative of a pluripotent stem cell state. piPSCs were highly proliferative, maintained a normal karyotype, and have been expanded for more than 50 passages. These results demonstrate that GALT-KO piPSCs can be successfully generated and have qualities necessary to serve as a robust cell source for the development of genetically engineered pigs for xenotransplantation.
Materials and Methods
Cell culture and transduction
GALT-KO fibroblasts were cultured in fibroblast medium [Dulbeco's modified Eagle medium (DMEM) high glucose (Hyclone) with 10% fetal bovine serum (FBS; Hyclone), 4 mM
Alkaline phosphatase, immunocytochemistry, and flow cytometry
Alkaline phosphatase (AP) staining was carried out with the VECTOR Red Alkaline Phosphatase Substrate Kit (Vector Laboratories) according to manufacturer's instructions. Immunocytochemistry and flow cytometry protocols followed were previously reported (West et al., 2010a). Primary antibodies used were POU5F1 (R&D Systems, 1:200), SOX2 (R&D Systems, 1:200), NANOG (Millipore, 1:200), βIII-TUB (Neuromics, 1:200), αSMA (Santa Cruz, 1:100), vimentin (R&D Systems, 1:200), SSEA1 (Developmental Studies Hybridoma Bank, 1:200), SSEA4 (Developmental Studies Hybridoma Bank, 1:200), TRA-1-60 (Millipore, 1:200), and TRA-1-81 (Millipore, 1:200). Primary antibodies were detected using fluorescently conjugated secondary antibodies Alexa Fluor 488 (Invitrogen, 1:500) and 594 (Invitrogen, 1:500). Galα-1,3-Gal epitope expression was analyzed with fluorescein isothiocyanate (FITC)-conjugated BS-I-B4 lectin (Sigma).
Proliferation and telomerase activity
The proliferation assay was performed by manual counts (n=3) every 12 h for 48 h. Population doubling time was determined using exponential regression curve fitting. Telomerase activity was determined using the TRAPeze XL Telomerase Detection Kit (Millipore) following the manufacturer's instructions. Statistical analysis was done using analysis of variance (ANOVA) and Tukey pair-wise comparisons between each population, with a p value<0.05 being considered significant.
Embryoid body formation and differentiation
Embryoid bodies (EBs) were formed by plating 2.0×106 iPSCs in mTeSR1 medium in an AggreWell plate (Stemcell Technologies). After 24 h, aggregates were harvested and maintained in mTeSR1 medium for 8 days. Differentiation was assessed by RT-PCR using the primers in Table 1. To assess differentiation by immunocytochemistry, EBs were plated and maintained in 20% KSR media without bFGF for an additional 2 days.
RNA isolation, DNA isolation, and PCR
RNA was isolated using the RNeasy QIAprep Spin miniprep Kit (Qiagen) per the manufacturer's instructions. mRNA was reverse transcribed using the iScript cDNA Synthesis Kit (Bio-Rad Laboratories). PCR amplification was performed using GoTaq Green master mix (Promega). Primers used in RT-PCR are listed in Table 1. PCR reactions conditions were: Denaturing cDNA at 95°C for 3 min, 30 cycles of denaturing at 95°C for 30 sec, annealing at 60°C for 30 sec, extension at 72°C for 30 sec, with a final 10-min extension at 72°C.
For genomic DNA analysis, DNA was extracted per manufacturer's instructions, and 500 ng of DNA was used for PCR. Fragments were amplified by using a LongRange PCR Kit (Qiagen) with three GALT-KO primer sets (see Table 1). The cycle conditions were 94°C for 2 min, followed by 94°C for 15 sec, 65°C for 30 sec, and 68°C for 10 min, and 20 sec per cycle for 30 cycles and a final extension step of 68°C for 7 min.
Results
GALT-KO piPSCs display morphological characteristics and immunoreactivity consistent with a pluripotent fate
Porcine GALT-KO fibroblasts (Fig. 1A) were transduced with six human pluripotency genes—hPOU5F1, hNANOG, hSOX2, hLIN28, hC-MYC, and hKLF4—driven by the EF1-α promoter. After 24 h, cells were replated on feeder cells in stem cell expansion medium. Putative GALT-KO piPSCs were observed around 8 days after transduction and grew as compact colonies. Colonies were selected mechanically and initially replated on feeder cells in stem cell expansion medium. Morphologically, GALT-KO piPSCs at the single cell level showed clear cell borders, high nuclear-to-cytoplasm ratios, and prominent nucleoli (Fig. 1B). GALT-KO piPSCs were then transferred to feeder-free conditions in mTeSR1 medium and on Matrigel (Fig. 1C). GALT-KO piPSCs were strongly positive for AP (Fig. 1D). Immunocytochemistry revealed that iPSCs were strongly positive for the introduced factors POU5F1 (Fig. 1E–H), SOX2, and NANOG, whereas pig fibroblasts were negative (Fig. 1I, J). To determine if cells were positive for nonintroduced pluripotency markers, they were stained for SSEA1, SSEA4, TRA-1-81, and TRA-1-60. GALT-KO piPSCs were highly positive for SSEA1 (Fig. 2A, C) and SSEA4, whereas only a limited number of cells were positive for TRA-1-60 (Fig. 2E, G) and TRA-1-81. To quantify expression of nontransduced pluripotency markers, flow cytometry was performed and confirmed that SSEA1 (∼95%) and SSEA4 (∼40%) were highly expressed in piPSCs (Fig. 2B, D), whereas TRA-1-60 and TRA-1-81 were expressed in a smaller subset of cells (Fig. 2F, H). These results indicate that GALT-KO fibroblasts were successfully reprogrammed.
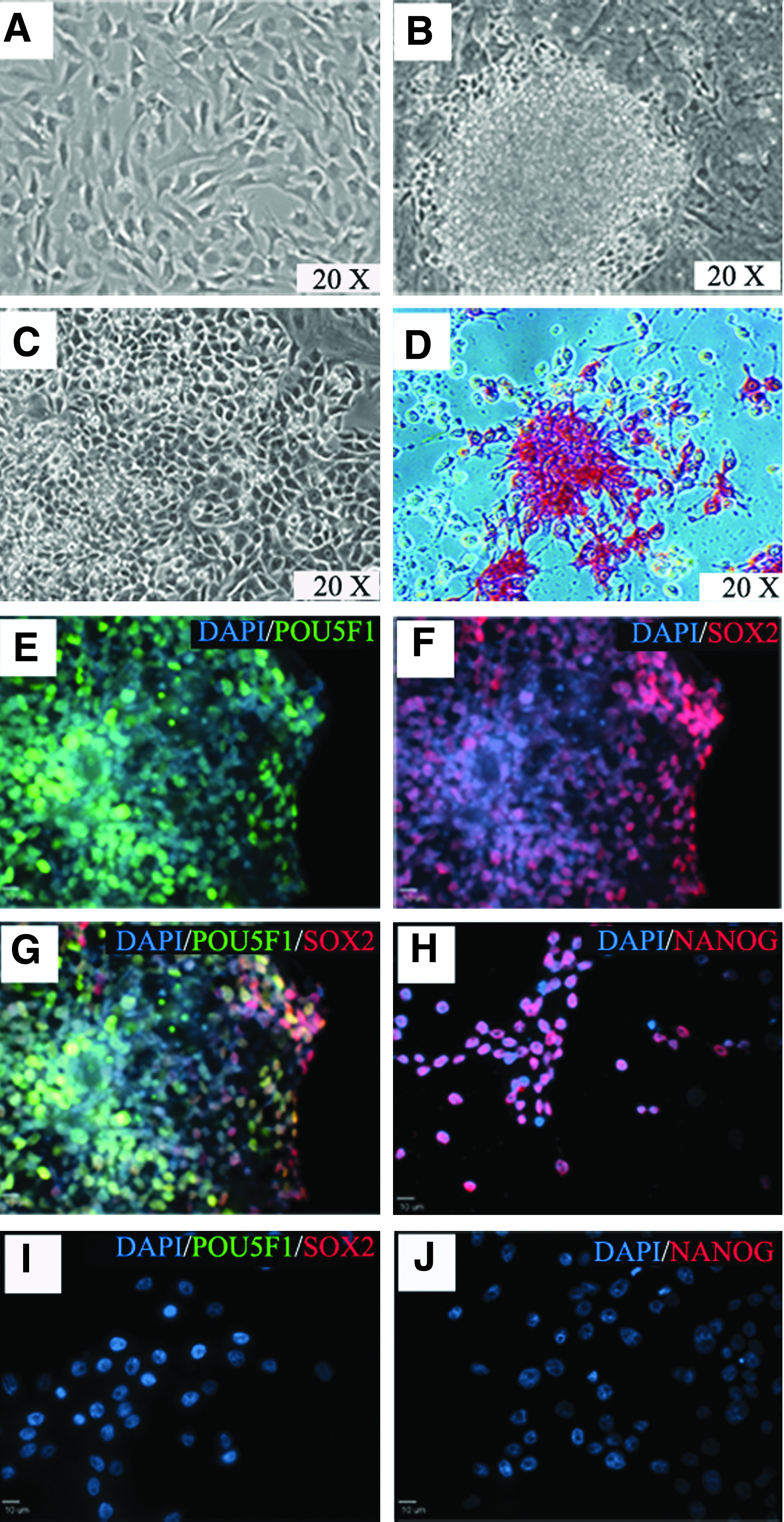
Generation of GALT-KO piPSCs from fibroblasts. GALT-KO fibroblasts (
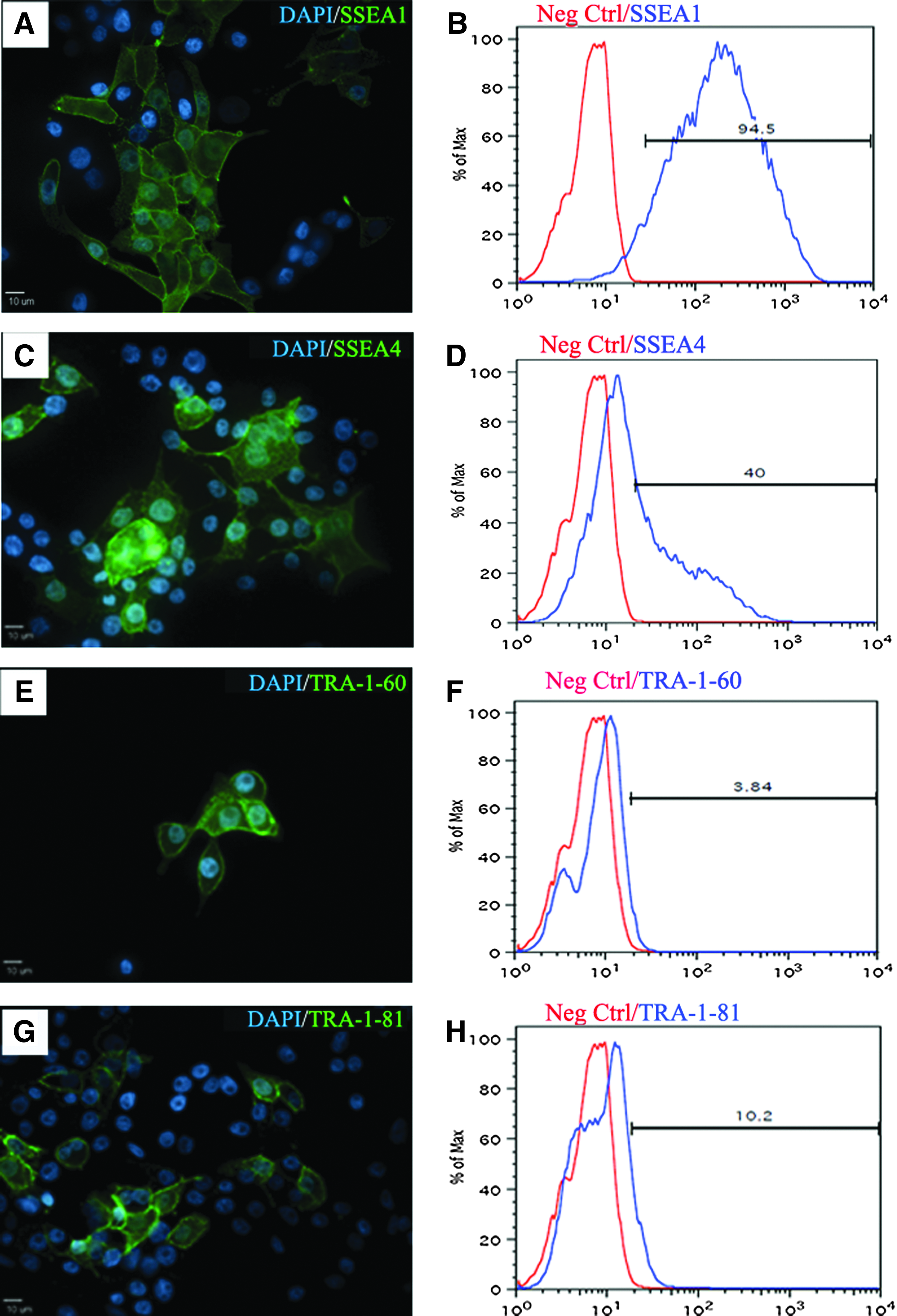
Gal-KO iPSCs express endogenous pluripotent markers. Immunostaining showed expression of SSEA1 (
Exogenous and endogenous pluripotent genes are expressed in GALT-KO piPSCs
PCR and RT-PCR using human specific primers were performed to determine if all six human reprogramming factors were integrated and expressed in GALT-KO piPSCs. PCR results showed that all six factors were integrated into the genome of piPSCs, similar to human iPSCs (hiPSCs) positive control cells (Fig. 3A). GALT-KO fibroblast parent cells (GALT-KO F) were negative for all six human genes. RT-PCR results showed that the human reprogramming genes were expressed in the piPSCs and positive control human (h) ESCs, whereas reprogramming gene expression was absent in GALT-KO fibroblasts (Fig. 3B). In previous studies, overexpression of exogenous pluripotency genes resulted in the reactivation of endogenous genes (Esteban et al., 2009; West et al., 2010b; Yu et al., 2007). RT-PCR results showed that the porcine POU5F1 and SOX2 genes were reactivated by the overexpression of exogenous genes (Fig. 3C). In addition, other pluripotency network genes were expressed including TERT, CDH1, TDGF1, Dax1, and Dnmt3b in the piPSC population, yet were not expressed in GALT-KO fibroblasts (Fig. 3D).
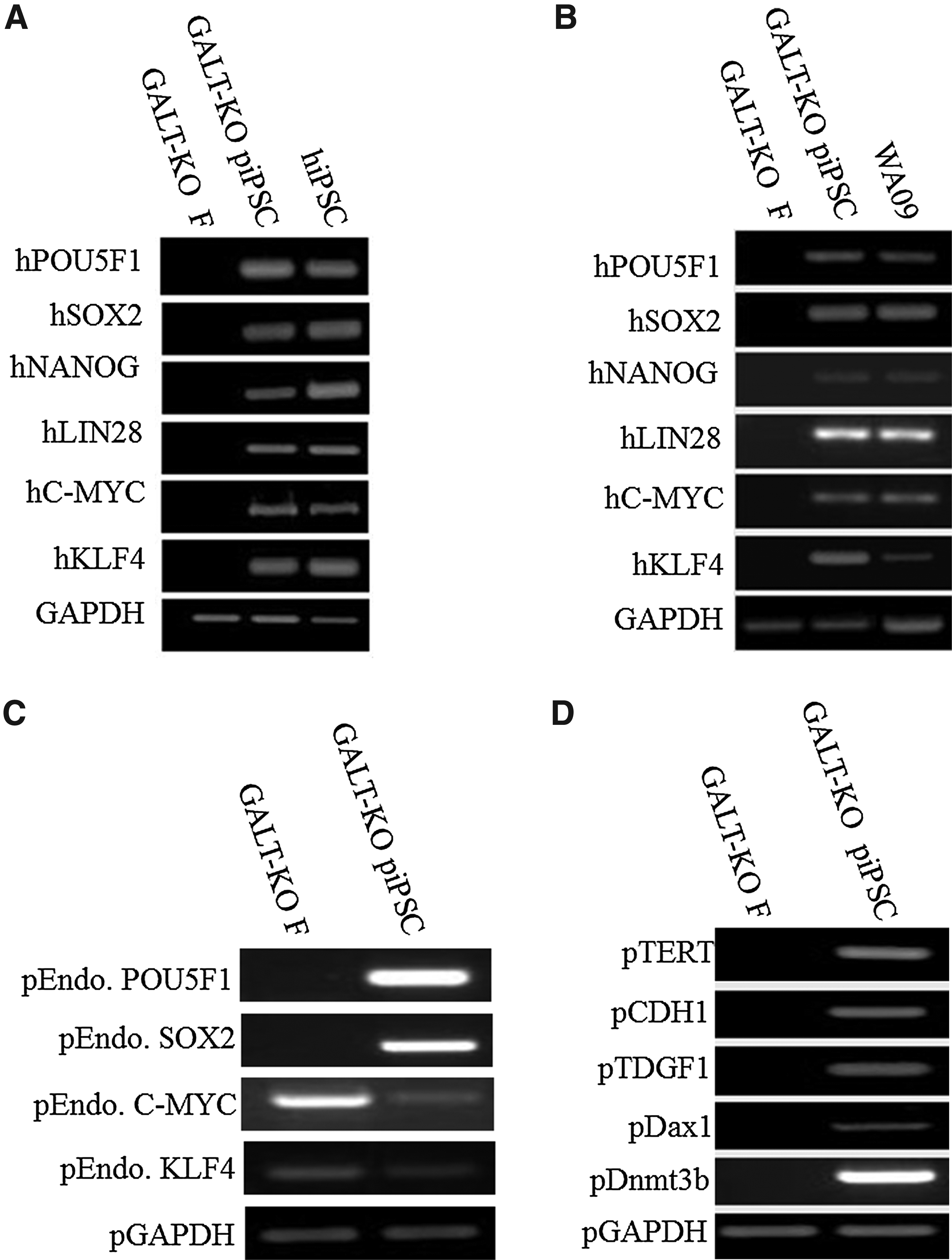
Activation of endogenous pig pluripotency network in GALT-KO piPSCs. piPSCs showed stable integration of the six reprogramming genes (
GALT-KO piPSCs are highly proliferative and differentiate into all three germ layers
High levels of telomerase activity and rapid proliferation are hallmarks of pluripotent stem cells. Telomerase activity revealed a significant (p<0.01) increase of six-fold from 42.5 total product generated (TPG) in GALT-KO fibroblasts to 249.3 TPG in GALT-KO piPSCs (Fig. 4A). Telomerase activity of GALT-KO piPSCs was significantly less than the hESC positive control (356 TPG) and HeLa cell line (465 TPG), yet was closer to hESC activity levels than GALT-KO fibroblasts. To determine the doubling time of GALT-KO piPSCs, cells were plated and quantified every 12 h for 48 h. The population doubling time of GALT-KO piPSCs was 19.4 h, which was significantly (p<0.01) faster than the GALT-KO fibroblasts (33.7 h) and similar to WA09 hESC control cells (22.7 h; Fig. 4B). GALT-KO piPSCs are highly proliferative and are passaged every 3 days. Cells have been maintained for more than 50 passages without loss of pluripotent phenotype. The karyotypes of GALT-KO piPSCs were tested at passage 20 with 20 cells randomly selected for G-banding. All tested cells were found to be normal (Fig. 4C).

GALT-KO piPSCs demonstrate rapid proliferation and high levels of telomerase activity. Telomerase activity in piPSCs was significantly (p<0.01) higher than GALT-KO F and was more similar to WA09 hESCs (
The potential of GALT-KO piPSCs to form ectoderm, mesoderm, and endoderm was tested by EB differentiation (Fig. 5B). RT-PCR analysis of day 8 EBs showed expression of Nestin (ectoderm), CD34 (mesoderm), and AFP (endoderm) in EBs, but not in GALT-KO piPSCs (Fig. 5A). Immunostaining of plated EBs after 10 days of differentiation showed cells positive for βIII-TUB (ectoderm, Fig. 5C), αSMA (mesoderm, Fig. 5D), and vimentin (endoderm, Fig. 5E). These results indicated that GALT-KO piPSCs could differentiate into cells from all three germ layers.
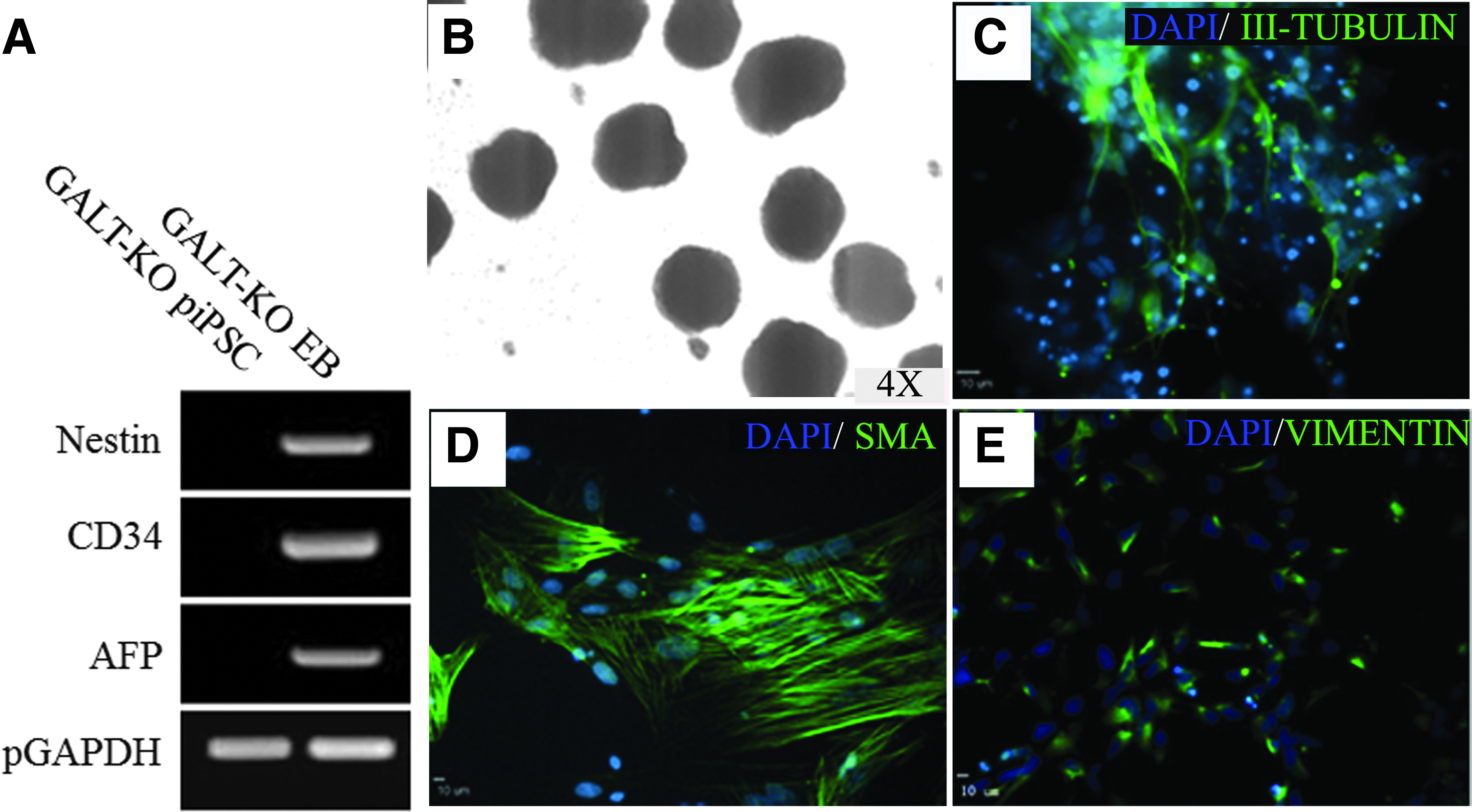
EB-mediated differentiation results in the formation of all three germ layers. piPSCs were differentiated as EBs (
GAL epitope is absent in GALT-KO piPSCs
Disruption of the GALT gene was originally achieved by insertion of the PGK-neo gene into exon 9 resulting in the loss of function (Fig. 6A) (Sharma et al. 2003). To confirm the disruption of the GALT gene, PCR analysis was performed with the 8S, 9A primer set designed to amplify exons 8 to 9. GALT−/− (lane 3) cells showed a single PCR band with increased size (9.3 kb) due to the insertion of the PGK-neo gene, whereas the GALT−/+ (lane 2) cells showed a long (9.3 kb) and short (7.6 kb) form PCR product and GALT+/+ (lane 1) cells only possessed the short (7.6 kb) form PCR product (Fig. 6B). To further confirm the PGK-neo gene interruption of the GALT gene, two additional primer sets were used, 3.3 kb (lanes 4–6) and 2.9 kb (lanes 7–9), where the forward primer binds within the PGK-neo gene and the reverse primer binds within the GALT gene. Wild type (+/+) cells were negative using both primer sets (lanes 4 and 7), whereas heterozygous (lanes 5 and 8) and homozygous (lanes 6 and 9) cells were positive. These data indicate that both alleles of the GALT gene were indeed interrupted.
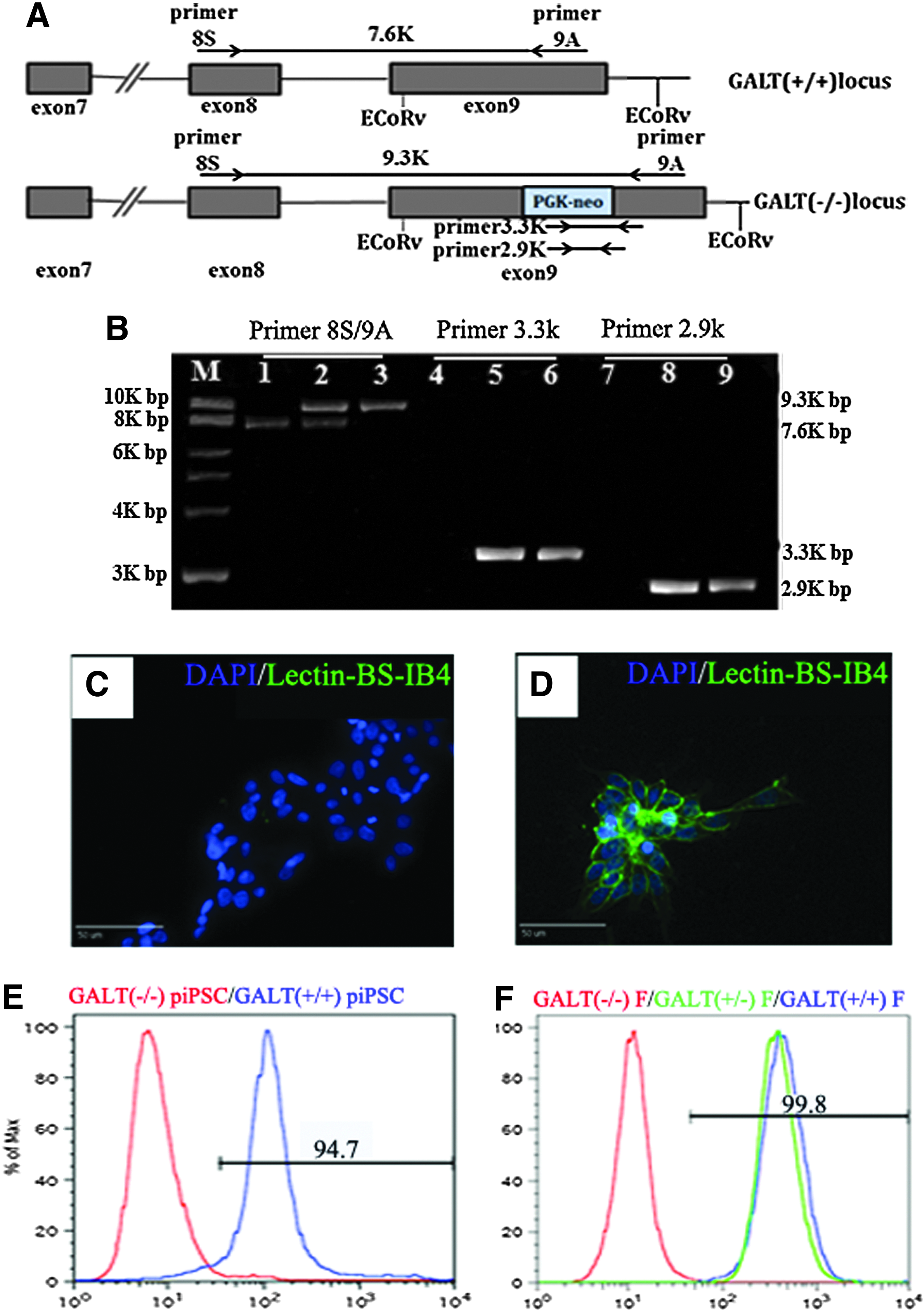
GAL epitope absent in GALT-KO piPSCs. Disruption of the GALT gene is achieved by insertion of PGK-neo gene into exon 9 (
The lectin BS IB4 is specific for the terminal GAL residue and has been used extensively in xenotransplantation research to test for the presence of the Gal antigen (Sharma et al., 2003). Immunostaining showed that GAL-KO piPSCs were negative for BS-IB4 (Fig. 6C), whereas piPSCs derived from a wild-type (WT) pig were positive (Fig. 6D). Flow cytometry results again showed that BS-IB4 was negative in GALT−/− piPSCs (red) and positive in GALT+/+ piPSCs (blue, piPSCs previously derived from wild type pig; Fig. 6E). GALT+/+ (blue) and +/− (green) fibroblasts were positive for BS-IB4, whereas GALT−/− parent fibroblasts were negative (red; Fig. 6F). These findings demonstrate that GALT-KO piPSCs are negative for the terminal GAL residue.
Discussion
In this report we describe for the first time the derivation and characterization of a GALT-KO piPSC line that shows robust stem cell characteristics and has significant potential in developing pigs for xenotransplantation. GALT-KO piPSCs demonstrated significant stem cell characteristics similar to previously reported piPSC lines despite the knockout of the GALT gene and the loss of the Gal epitope (Aflatoonian et al., 2009; Cheng et al., 2012; Esteban et al., 2009; Ezashi et al., 2009; Hall et al., 2012; Telugu et al., 2010; West et al., 2010b; West et al., 2011; Wu et al., 2009). GALT-KO piPSCs express the transduced pluripotency reprogramming factors POU5F1, SOX2, and NANOG and also show expression of independent pluripotency markers SSEA1 and SSEA4 similar to previous studies (Cheng et al., 2012; Telugu et al., 2010). Functionally, piPSCs were able to differentiate in vitro into cells representing all three germ layers—ectoderm, endoderm, and mesoderm—demonstrating developmental plasticity. In addition, GALT-KO piPSCs were highly proliferative and possessed high levels of telomerase activity similar to immortal pluripotent stem cell lines, while maintaining a stable karyotype (Takahashi et al., 2007; Takahashi and Yamanaka, 2006; Thomson et al., 1998).
The use of highly proliferative and immortal cells makes the process of gene targeting feasible, particularly when multiple genes are being manipulated, and has led to the extensive use of mouse ESCs for the generation of transgenic animals (Babinet and Cohen-Tannoudji, 2001; Capecchi, 2005; Gondo et al., 2009; Mishina and Sakimura, 2007). GALT-KO piPSCs showed rapid proliferation with a doubling time of 19 h, comparable to previously reported doubling times for piPSCs, ranging from 17 to 24 h (Ezashi et al., 2009; West et al., 2010b; Wu et al., 2009), and significantly higher levels of telomerase activity relative to fibroblast starting populations (Esteban et al., 2009; Ezashi et al., 2009; Telugu et al., 2010; Wu et al., 2009). Both proliferation and telomerase activity were comparable to hESCs, further indicating that GALT-KO piPSCs have significant expansion capabilities. GALT-KO piPSCs showed genetic stability with a normal karyotype after 20 passages. These important characteristics suggest that GALT-KO piPSCs can be used as an excellent starting population for gene targeting of other critically important xenotransplantation genes.
The shortage of organs and tissues for transplant has led to a critical need that is unmet by human donors, making pig-to-human xenotransplantation an attractive option. However, genetically modified pigs with multiple genes knocked out and knocked in are needed to prevent immune rejection. GALT piPSCs that are immortal and highly proliferative offer a suitable cell source for gene targeting that has not been previously available. These cells, once modified, can be used in standard SCNT procedures or even to generate germ line–competent chimeric animals for the production of xenotransplant pigs.
Footnotes
Acknowledgments
We would like to thank Mrs. Julie Nelson and the Center for Tropical and Emerging Global Diseases Flow Cytometry Facility, University of Georgia. We would also like to thank Mike Daniel and Amber Williams for their assistance with sample collection and Erin Jordan for her technical assistance.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.