
Abstract
Abstract
Kunming (KM) mice are the most widely used strain in China. However, authentic embryonic stem cells (ESCs) from KM mice have never been available, and this hampers the genetic manipulation of this valuable mice strain. In this study, we show that KM ESCs can be efficiently derived and maintained in chemically defined N2B27 medium with the presence of two small molecules PD0325901 and CHIR99021 (2i medium). These KM ESCs exhibit all features of ESCs, including long-term self-renewal ability, expression of key molecular markers (Oct4, Nanog, and Sox2), the ability to form teratomas, and the capacity to incorporate into the developing embryo and then transmit through the germ line.
Introduction
In this study, our results showed that ESCs from KM mice could be derived and maintained in serum-free, feeder-free 2i medium. The established KM mice ESCs showed long-term self-renewal ability in vitro. In addition, these ESCs expressed pluripotency markers and retained the capacity to differentiate into derivatives of all three germ layers. Most importantly, the ESCs maintained the ability for generating functional gametes in chimeras.
Materials and Methods
Derivation and propagation of KM ESCs
Mice were purchased from Shanghai Experimental Animal Center, Chinese Academy of Sciences. All animal experiments were carried out in adherence with the National Institutes of Health Guidelines on the Use of Laboratory Animals and approved by the Second Military Medical University Committee on Animal Care (EC11-055). X-ray–inactivated mouse embryonic fibroblasts were prepared as feeder. The KM mice embryos were gently flushed out from the uterine horn of KM mice at embryonic (E) day 3.5 with phosphate-buffered saline (PBS), washed three times with N2B27 medium, individually placed on a 96-well plate (one embryo per well), and cultured in 2i medium. The culture medium was changed by half every day. After 3 days, the adherent primary explants were separately trypsinized into small clumps and replanted onto a fresh gelatin-coated 96-well plate. The KM ESCs were propagated in serum-free, feeder-free 2i medium at 37°C, 5% CO2, and 100% humidity. The chemically defined N2B27 medium was prepared as previously described (Ying et al., 2003b). The 2i medium was prepared by the addition of 1 μM PD0325901 (Stemgent, San Diego, CA, USA) and 3 μM CHIR99021 (Stemgent, San Diego, CA, USA) to N2B27 medium (Nichols et al., 2009b). Other culture medium and growth factors were purchased from Invitrogen (Carlsbad, CA, USA).
In vitro differentiation of KM ESCs
Differentiation of KM ESCs was induced by the formation of embryoid bodies (EBs). The cells were dissociated by 0.25% trypsin and cultured in ultra-low-attachment 24-well plates in Dulbecco's modified Eagle medium (DMEM) medium plus 10% fetal bovine serum (FBS). The cells grew in suspension and formed EBs. The medium was changed every other day. After 7 days, the EBs were plated onto a Matrigel-coated dish in DMEM plus 10% FBS. Three to 7 days later, the cells were fixed and the expression of markers for the three germ layers was examined.
Alkaline phosphatase staining, karyotype analysis, and immunoassaying
Alkaline phosphatase (AP) staining was performed according to the manufacturer's instructions using the AP detection kit (Millipore, Bedford, MA, USA). Karyotype analysis was performed according to Yasumura et al. (1966). Immunoassaying was performed according to the standard protocols. Briefly, cells were fixed in 4% paraformaldehyde (PFA) for 15 min at room temperature (RT), washed three times with PBS, and then incubated in blocking buffer (BB) (0.3% Triton X-100 and 1% bovine serum albumin) in PBS for 30 min at RT. Afterward, the cells were incubated with primary antibody for 12 h at 4°C in BB and washed three times with PBS. After that, the cells were incubated with secondary antibody at a dilution of 1:500 for 30 min at RT and washed three times with PBS. The cells were then incubated with 0.5 μg/mL 4′,6-diamidino-2-phenylindole (DAPI) for 5 min for staining of nuclei. Primary antibodies used in this study include the following: Mouse anti-Oct4 (1:200 dilution; Santa Cruz Biotechnology, Santa Cruz, CA, USA), rabbit anti-Nanog (1:500 dilution; Abcam, Cambridge, MA, USA), mouse anti-Sox2 (1:500 dilution; Millipore), goat anti-GATA4 (1:200 dilution; Santa Cruz), mouse anti-tyrosine hydroxylase (TH) (1:200 dilution; Abcam), mouse anti-β-III tubulin (1:200 dilution; Abcam), goat anti-α-fetoprotein (AFP) (1:200 dilution; Santa Cruz).
Teratoma formation
KM ESCs (2×106 per mouse) were injected subcutaneously into the dorsal flank of nude mice. Tumors were observed and collected after 2 months. The tumors were then fixed overnight in 4% PFA, embedded in paraffin, and sectioned and stained with Hematoxylin & Eosin by standard procedures.
Flow cytometry assay
The cells were dissociated into single cells using 0.05% trypsin and fixed using BD Cytofix (BD Biosciences, Franklin Lakes, NJ, USA). After that, the cells were washed three times with PBS and then incubated in BB for 30 min to block nonspecific protein–protein interactions. The cells were then incubated with the primary antibody mouse anti-Rex1 (1:200 dilution, Abcam) for 60 min at 4°C. The secondary antibody, Alexa Fluor 488 goat anti-rabbit immunoglobulin G (IgG) was then applied for 60 min at RT (1:1000 dilution, Abcam).
Aggregation of KM ESCs with zona-free embryos
The eight-cell embryos were recovered from the oviduct of 129 females at E 2.5. The embryos were incubated with acidified Tyrode's solution until the zona pellucida had disintegrated, and then cultured in microdrops of KSOM medium supplied with10% FBS under mineral oil. Clumps of KM ESCs (10–20 cells) were collected and transferred into microdrops containing zona-free eight-cell embryos. Then the embryos were cultured overnight at 37°C, 5% CO2, and the aggregated embryos were then transferred into pseudo-pregnant recipient females. Chimeric mice were identified by coat color. Germ-line transmission was tested by mating chimeras with KM mice.
Results
Efficient derivation and maintenance of KM mice ESCs
To derive the KM ESCs, KM mice blastocysts were placed on feeder-coated plates in 2i medium (0.4 μM PD0325901 and 3 μM CHIR99021) as previously described (Ying et al., 2008). After 3 days, adherent primary explants formed from KM mice blastocysts (Fig. 1A). However, the cells showed a mixture of differentiated cells and undifferentiated colonies (data not shown). To inhibit differentiation, a higher dose of PD0325901 was used, as in Nichols et al. (2009b). Our results showed that the differentiation blocking effect of 1 μM PD0325901 was dominant. Combined with CHIR99021, the cells propagated continuously and long term as undifferentiated colonies (Fig. 1A). As a control, the outgrowth of KM mice blastocysts differentiated in N2B27 medium within 3 days (Fig. 1A).
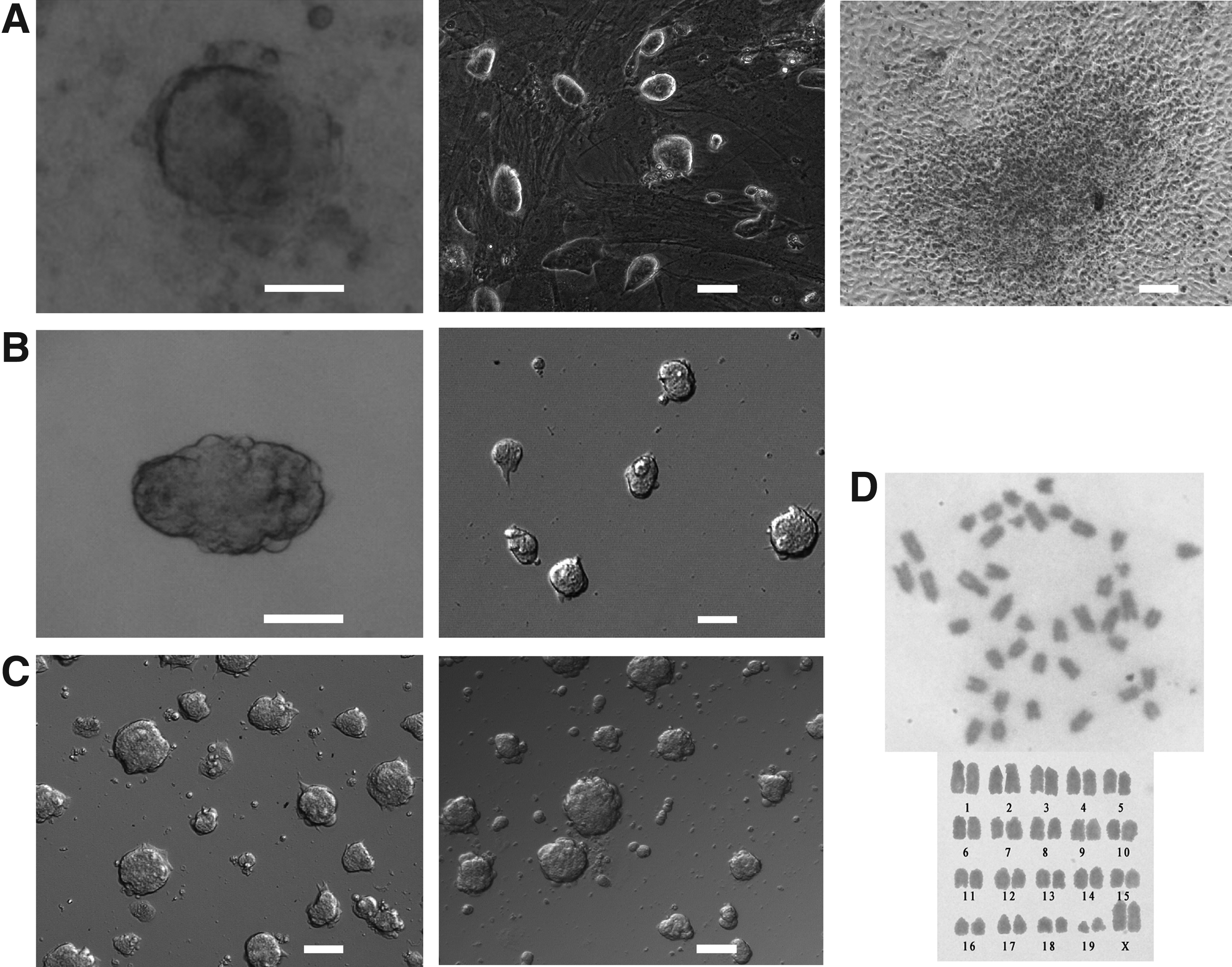
(
We then examined whether feeder was necessary for deriving and maintaining the undifferentiated state of the KM ESCs. Every blastocyst of KM mice was separately plated onto gelatin-coated plastics and cultured in 2i medium with a higher dose of PD0325901 (1 μM). After 3 days, one adherent primary explant formed (1 primary explant out of 17 blastocysts) from the KM mice blastocyst, and the primary explant was propagated in the serum-free and feeder-free 2i medium (Fig. 1B). However, the primary explant formation rates were low, and most of the blastocysts did not hatch out from the zona pellucida. We then added 10% knockout serum replacement (KSR) into the 2i medium to facilitate the survivability of KM mice blastocysts. Results showed that KSR facilitated KM mice blastocysts to survive and form adherent primary explants (13 primary explants out of 15 blastocysts). The resulting primary explants were propagated in the feeder-free and serum-free 2i medium (Fig. 1B). The three derived ESC-like cell lines were named EH-KES1, EH-KES2, and EH-KES3. Two of them (EH-KES1 and EH-KES2) have been propagated continuously (Fig. 1C). Regarding karyotype, 75% cells of EH-KES1 (passage 74) maintained normal karyotypes (15 normal mitotic spreads out of 20), whereas 85% cells of EH-KES2 (passage 59) maintained normal karyotypes (17 normal mitotic spreads out of 20).
KM ESCs maintained pluripotency in 2i medium
For ESCs, the pluripotent state is largely governed by the core transcription factors Oct4, Sox2, and Nanog (Chambers and Smith, 2004; Niwa, 2007; Silva and Smith, 2008). The immunostaining results showed that the EH-KES1 maintained Oct4 expression on gelatin in 2i medium (Fig. 2A). Moreover, the expression of pluripotent markers such as Nanog and Sox2 from EH-KES1 in 2i medium was also observed (Fig. 2A). As a control, the EH-KES1 did not show Oct4 expression in N2B27 medium (Fig. 2A).
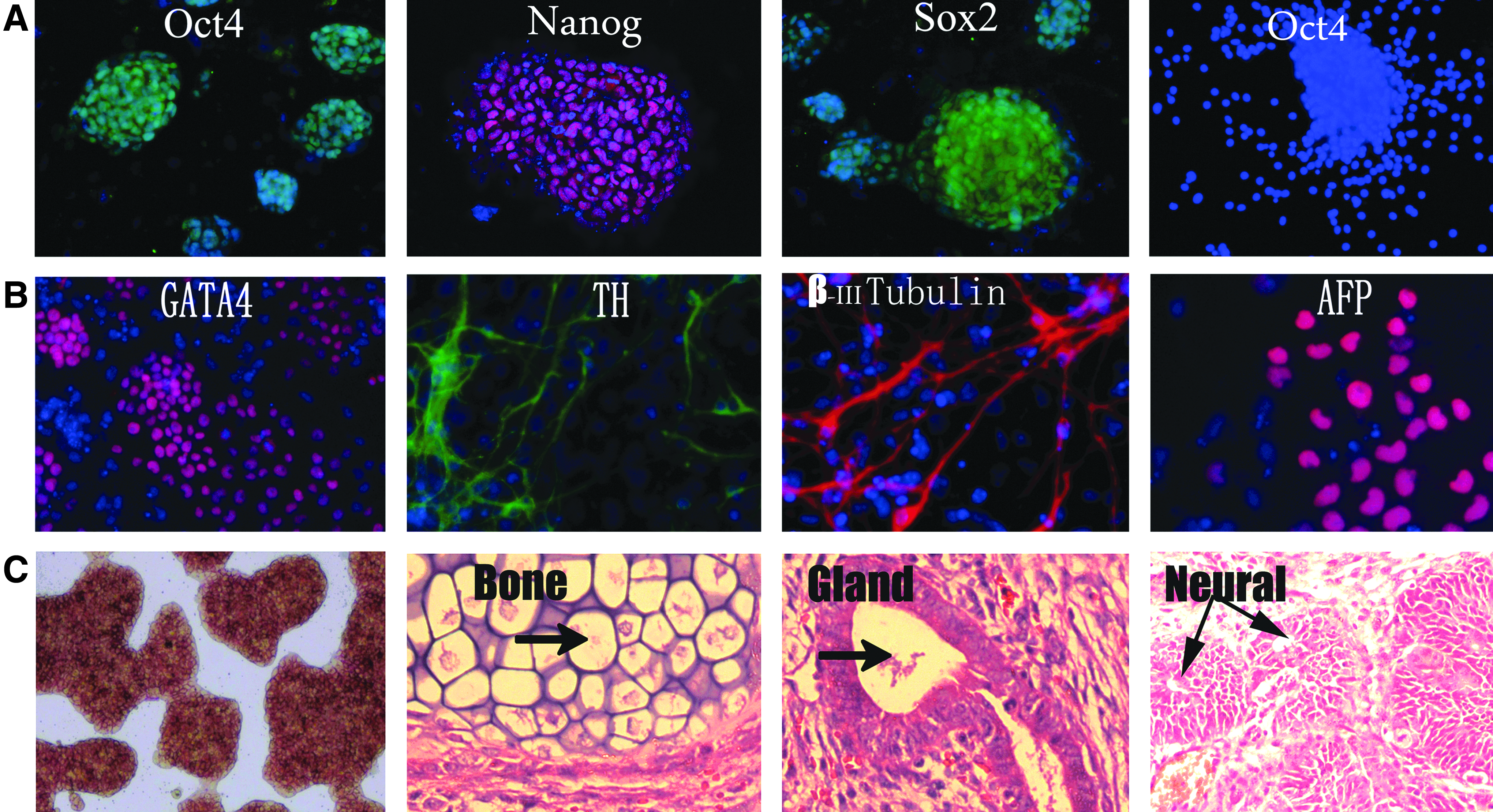
(
To confirm the maintenance of pluripotency for the ESCs lines, we performed ESC differentiation using EB methods as previously described (Keller 1995). Within the EBs, ESC differentiation proceeds on a schedule similar to that in the embryo. After the removal of 2i medium, the dissociated EH-KES1 cells were plated onto Petri dishes (for suspension culture) in the presence of 10% FBS. The immunostaining results showed positive expression of markers for ectoderm (TH and β-III tubulin), mesoderm (GATA-4), and endoderm (AFP) in the outgrowth of EBs (Fig. 2B). The ESCs also stained positive for AP when cultured on gelatin-coated plastics. The injection of EH-KES1 into nude mice revealed that the ESCs can form teratomas in nude mice within 3 months, and the formed tumors contained bone (mesoderm), gland (endoderm), and neural tissues (ectoderm) (Fig. 2C). In summary, these results confirmed that the KM ESCs maintained the pluripotency to differentiate into cells and tissues from all three germ layers in the chemically defined feeder-free condition.
KM ESCs can form chimeras and transmit through the germ line
We then assayed the potential of EH-KES1 to form germ-line chimeras. To obtain chimeras, clumps of EH-KES1 (10–20 cells) were transferred into microdrops containing zona-free, eight-cell 129 mice embryos. The embryos were cultured overnight and then transferred to recipient females. A total of 176 blastocysts were obtained and transferred to 20 pseudo-pregnant 129 mice. Among the five pups obtained, two chimeras apparently showed white coat color, an indication of the presence of EH-KES1. These chimeras were then mated with KM mice; one female chimera produced two offspring with white coat color (Fig. 3A), indicating that the EH-KES1 cells maintained germ-line transmission ability.
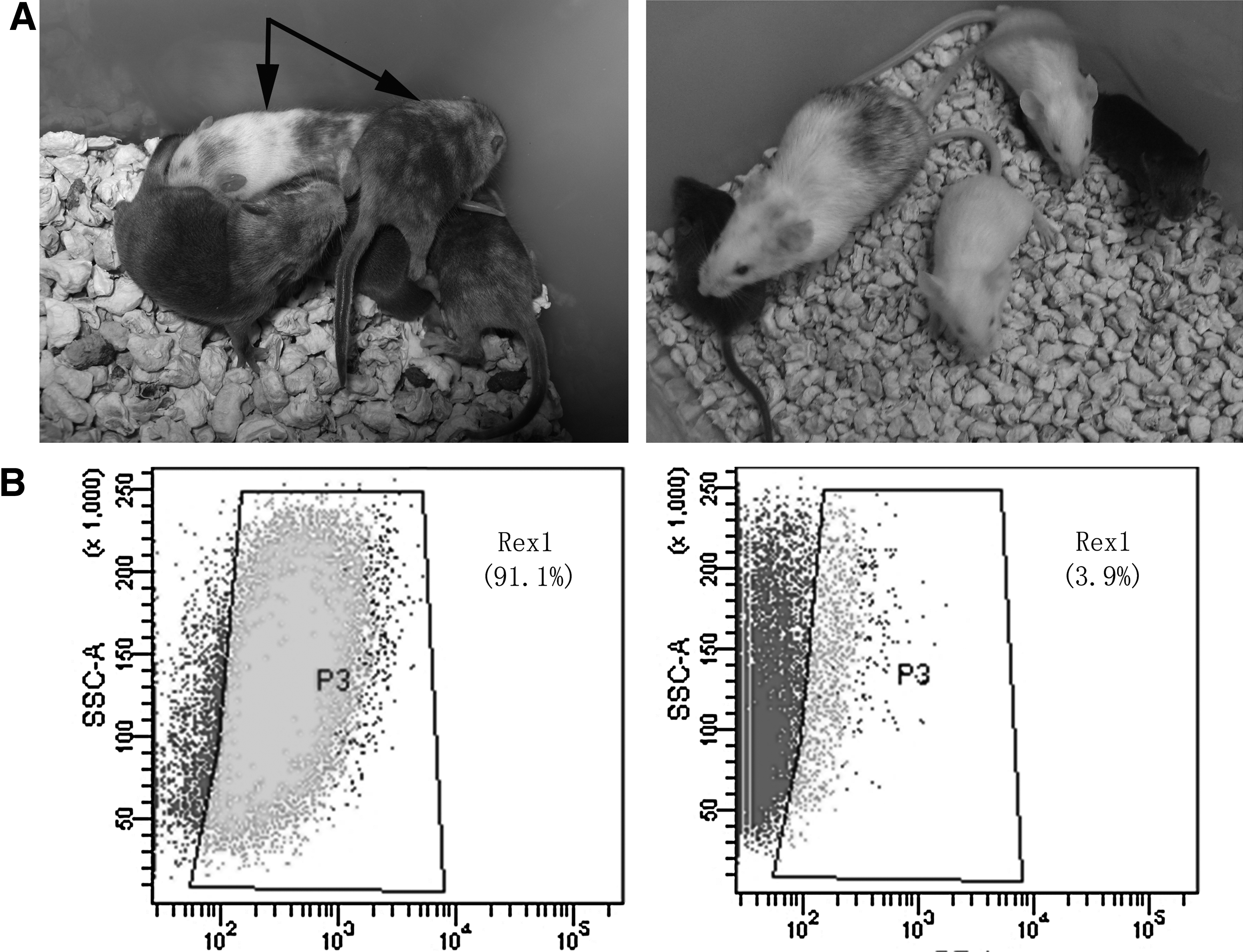
(
To analyze the mechanism underlying the KM ESCs self-renewal, we cultured the EH-KES1 cells on gelatin-coated plastics in two different culture media: (1) 2i medium; (2) DMEM medium supplied with 20 ng/mL LIF and 10% FBS. After two passages, a flow cytometry assay was performed to determine the Rex1 expression of EH-KES1. Our results showed that 91.1% of the cells cultured in 2i medium showed Rex1 expression (Fig. 3B). In contrast, only 3.1% of the cells cultured in LIF and FBS medium maintained Rex1 expression (Fig. 3B). These results suggest that LIF is unnecessary and insufficient for maintaining Rex1 expression of the KM ESCs.
Discussion
ESCs were initially cultured on a layer of feeders and serum to obtain the necessary factors such as LIF (Smith, 2001) and bone morphogenetic proteins (BMPs) (Ying et al., 2003a) for self-renewal and pluripotency. However, authentic KM murine ESCs have never been established by using LIF and serum conditions, and KM mice were generally believed to be unsuitable for the isolation of ESCs. Our results showed that 1 μM PD0325901 combined with 3 μM CHIR99021 can efficiently maintain the long-term self-renewal of the KM ESCs (Fig. 2A). Feeders can produce LIF and may mask any effects of the 2i medium. In this study, we maintained KM-ESCs on gelatin-coated plastics to confirm that KM ESCs could maintain long-term self-renewal without addition of LIF (Fig. 1B).
Differentiation of ESCs involves autoinductive stimulation of ERK signaling by fibroblast growth factor-4 (FGF4) (Kunath et al., 2007; Stavridis et al., 2007). Addition of PD0325901 inhibited ESC differentiation, whereas CHIR99021 maintained overall viability of ESCs when ERK signaling was totally inhibited (Ying et al., 2008). Moreover, downregulation of GSK3 can facilitate the Oct4 expression and self-renewal ability of human and mice ESCs (Murray et al., 2004; Sato et al., 2004). In this study, we showed that KM ESCs could maintain Oct4 expression in the 2i medium (Fig. 2A). Moreover, the EH-KES1 maintained the capacity for incorporation into the developing embryo and then transmitting through the germ line (Fig. 3A).
Our results also showed that the KM ESCs maintained Rex1 expression in the 2i medium and quickly lost Rex1 expression after being transferred into LIF and serum medium (Fig. 3B). Previous reports showed that ESCs maintained in 2i medium were rather homogeneous in expression of pluripotency markers (Wray et al., 2011). In contrast, ESCs maintained in LIF plus serum were heterogeneous in the expression of pluripotency markers such as Nanog, Rex1, and Klf4 (Chambers et al., 2007; Hayashi et al., 2008; Toyooka et al., 2008). A recent report showed that expression of Rex1 was important for maintaining the self-renewal capacity of ESCs (Marks et al., 2012) and the Rex1-negative ESCs had largely lost self-renewal ability. Consistent with previous studies, our results show that Rex1 may serve as a key pluripotent regulator of the KM ESCs.
KM mice have several advantages, such as high disease resistance, large and frequent litters, and rapid growth. The derivation of germ-line–competent KM ESCs can facilitate targeted genetic manipulation in this valuable biomedical model species. Moreover, chemically defined conditions also provide a better-tuned environment for studying the modeling of gene expression during ESCs self-renewal.
Footnotes
Acknowledgments
This work was supported by the National Science Funds for distinguished Young Scholar (No. 30925037), the Science Fund for Creative Research Group of NSFC (No. 30921006), and National High Technology Research and Development Program (No. 2012AA020806).
Author Disclosure Statement
The authors declare that no conflict of financial interest exists.