
Abstract
Abstract
Previous studies have demonstrated that skin stem cells expressing the pluripotency marker stage-specific embryonic antigen 3 (SSEA3) are easier to reprogram into induced pluripotent stem cells (iPSCs) than skin fibroblasts. Furthermore, it is widely speculated that the undifferentiated state may make stem cells more efficient donor cells for somatic cell nuclear transfer (SCNT). In this study, we isolated SSEA3+ cells from goat skin fibroblast cells (SFCs) using fluorescence-activated cell sorting (FACS) and examined expression of pluripotency markers and in vitro development of cloned embryos following SCNT. Results showed that cell clusters from SSEA3+ cells were consistently positive for alkaline phosphatase staining and pluripotency markers, Nanog, Oct4, Sox2, and SSEA3. The cleavage rate of cloned embryos derived from SSEA3+ cells did not differ compared with SFCs (70.5±0.8% and 68.4±2.1%, respectively), but was significantly higher compared with SSEA3− cells (64.9±1.6%, p<0.05). The blastocyst rate was significantly increased in the SSEA3+ cell group compared with the SFC and SSEA3− cell groups (30.3±1.2% vs. 21.2±0.9 and 19.0±1.0%, respectively, p<0.05). The quality of cloned blastocysts from SSEA3+ cells was higher compared with SFCs and SSEA3− cells, based on total cell number and number of apoptotic cells per blastocyst. These findings suggest that using SSEA3+ cells as donors for SCNT is beneficial for enhancing in vitro development and quality of cloned goat embryos.
Introduction
Recently, two independent research teams isolated and characterized stage-specific embryonic antigen 3–positive (SSEA3+) cells from human skin fibroblasts (Byrne et al., 2009; Kuroda et al., 2010). These cells (named Muse) self-renew, express pluripotency markers, and differentiate into endodermal, ectodermal, and mesodermal cells, both in vitro and in vivo (Kuroda et al., 2010). Furthermore, SSEA3+ cells demonstrated enhanced efficiency to generate iPSCs, whereas iPSCs could not be derived from SSEA3− cells (Byrne et al., 2009; Wakao et al., 2011). We hypothesized that SSEA3+ adult stem cells may therefore be more efficient SCNT donor cells. In this study, we isolated and characterized SSEA3+ cells from goat skin fibroblast cells (SFCs) and investigated whether these cells could be suitable SCNT donor cells.
Materials and Methods
Unless indicated otherwise, all chemicals and reagents were purchased from Sigma Aldrich (St. Louis, MO, USA) and cell media were obtained from Gibco (Grand Island, NY, USA). All trials were conducted in accordance with the Guidelines for the Care and Use of Animals of the College of Veterinary Medicine, Northwest A&F University.
Preparation of goat SFCs
Goat SFCs were isolated from skin biopsies of Saanen dairy goats (Yangling Keyuan Biotechnology Inc., PR China). Skin samples were sterilized with 70% ethanol, washed in phosphate-buffered saline (PBS) solution, and minced into 1-mm3 pieces. Explants were seeded in a 60-mm culture dish and cultured in Dulbecco's modified Eagle medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 2 mM
Isolation of SSEA3+ cells by fluorescence-activated cell sorting
SSEA3+ and SSEA3− cells were isolated as previously described (Byrne et al., 2009). Briefly, SFCs at first-passage were washed twice in PBS and incubated with primary antibody against SSEA3 (1:100, R&D Systems) for 30 min at 4°C. After washing three times, cells were incubated with corresponding secondary phycoerythrin (PE)-conjugated goat anti-rat immunoglobulin G (IgG) antibody (1:500, Santa Cruz) for 30 min at 4°C. Cells were then washed three times and resuspended in 2 mL of ice-cold PBS with 2% FBS. Cells were immediately analyzed and sorted on an EPICS XL-MCL cell sorter (Beckman Coulter). SFCs incubated with secondary antibody were used as a negative control. Gating strategies and data analysis were performed with EXPO32 ADC analysis software (Beckman Coulter). Cells gated within the top 2–4% and the bottom 10% for SSEA3 expression were sorted into the SSEA3+ and SSEA3− populations, respectively. SSEA3+ and SSEA3− cells were allowed to adhere in culture as fibroblasts or were cultured in suspension. The number of chromosomes in the cells was determined using standard techniques (Nguyen and Reijo Pera, 2008).
Suspension culture
SFCs, SSEA3+, and SSEA3− cells were transferred into a 60-mm Ultra-Low Attachment dish (Corning) and cultured in 4 mL of DMEM containing 10% FBS. A volume equal to one-fifth of the initial culture medium was added to the dish every 2 days. Cells were seeded at an initial concentration of 0.5×104 cells/L. After 7 days in culture, cell clusters were counted, collected, and dissociated using 0.25% trypsin for 15 min, and returned to suspension culture. Two hours before dissociation, Y-27632 was added to the culture medium at a concentration of 10 μM and maintained until the following morning.
Alkaline phosphatase and immunofluorescence staining
Cell clusters derived from suspension culture were washed several times with PBS containing 0.2% polyvinyl alcohol. Staining was performed at room temperature in the dark using the Leukocyte Alkaline Phosphatase kit (Sigma) according to the manufacturer's instructions.
Immunofluorescence staining was performed as described previously (Byrne et al., 2009). Primary antibodies and their dilutions were as follows: Nanog (1:100, Abcam), Oct4 (1:100, Abcam), Sox2 (1:100, Abcam), and SSEA3 (1:100, R&D Systems). Cell clusters were treated with secondary antibodies, Alexa Fluor 488-labeled goat anti-rabbit IgG (Beyotime, A0423) for Nanog, Oct4, and Sox2, or Alexa Fluor 555-labeled goat anti-rat IgG (Beyotime) for SSEA3. Secondary antibodies were diluted 1:500 with Immnol Staining Secondary Antibody Dilution Solution (Beyotime, P0108). Finally, DNA was stained with 4′,6-diamidino-2-phenylindole (DAPI) (Beyotime, C1005) for 3 min. Samples were placed in a drop of PBS for analysis using a Nikon eclipse Ti-S microscope, equipped with a 198 Nikon DS-Ri1 digital camera (Nikon, Tokyo, Japan). Experiments were replicated three times.
Reverse transcriptase-polymerase chain reaction
Total RNA was extracted from SFCs, SSEA3+, and SSEA3− cells using RNAprep pure Micro Kit (Qiagen). First-strand cDNA was generated using PrimeScript® RT reagent Kit Perfect Real Time (TaKaRa) according to the manufacturer's instructions. PCR reactions were performed using Ex Taq DNA polymerase (TaKaRa). Primers used were as follows:
Oct4, sense 5′-GAAGCTGGACAAGGAGAAGCT-3′, antisense 5′-CATGCTCTCCAGGTTGCCTC-3′; Sox2, sense 5′-GAAAAACCAAGACGCTCAT-3′, antisense 5′-CTGGAGTGGGAAGAAGAGGT-3′; Nanog, sense 5′-ATGCCTGAAGAAAGTTACGC-3′, antisense 5′-AGGCTGTATGTTGAGAGGGT-3′; β-actin, sense 5′-CGGTGCCCATCTATGAGG-3′, antisense 5′-GATGGTGATGACCTGCCC-3′.
Nuclear transfer
Nuclear transfer (NT) was performed as previously described (Liu et al., 2011; Zhang et al., 2008). SFCs, SSEA3+, and SSEA3− cells at passages 2–4 were used as donor cells for SCNT to investigate the in vitro development of cloned embryos. Embryos were cultured for 7 days to evaluate their developmental rate.
Differential staining and apoptosis assay of cloned blastocysts
Differential staining and apoptosis assay were carried out in accordance with the methods of our previous study (Su et al., 2011). After 7 days of culture in vitro, cloned blastocysts from SFCs, SSEA3+ and SSEA3− cells were used for differential staining and apoptosis assaying. Blastocyst cell numbers were estimated by counting the total number of nuclei using DAPI, whereas the number of trophectoderm (TE) cells was estimated using immunostaining for CDX2. The number of inner cell mass (ICM) cells was determined as the total number of nuclei minus the number of TE nuclei. Apoptosis was investigated using the DeadEnd Fluorometric TUNEL System (Promega, Madison, WI, USA).
Statistical analysis
All data were analyzed using SPSS16.0 statistical software (IBM Corporation, Somers, NY, USA). Data were tested by one-way analysis of variance (ANOVA) and least significance difference (LSD) tests and reported as mean±standard error of the mean (SEM). For all analyses, p<0.05 was considered significant.
Results
Characterization of SSEA3+ cells isolated from skin fibroblasts
Three skin fibroblast cell lines were isolated from different goats (Fig. 1A). No differences were observed in morphology and proliferation between the different SFC lines. Fluorescence-activated cell sorting (FACS) analysis of first passage SFCs revealed 2–4% were SSEA3+. We subsequently isolated and cultured the approximate top 2% and bottom 10% of the SSEA3/PE-fluorescing cells as SSEA3+ and SSEA3− cells, respectively (Fig. 1B). Both populations were subjected to adherent or suspension culture. After adherent culture, cell morphology and proliferation were similar to that of SFCs. Under suspension culture, SSEA3+ cells formed cell clusters at 3 days, which grew in diameter to about 50–100 μm by 7 days (Fig. 1C). In contrast, SSEA3− cells did not generate clusters whereas SFCs formed a few cell clusters with a diameter of <50 μm after 7 days of suspension culture. Cell clusters from SSEA3+ cells proliferated for more than six passages in the presence of Y-27632; however, cell clusters from SFCs were difficult to expand in culture.

Isolation and characterization of SSEA3+ cells. (
Cell clusters from SSEA3+ cells were consistently positive for alkaline phosphatase (AP) staining (Fig. 1D). RT-PCR analysis showed strong expression of Nanog, Oct4, and Sox2 in SSEA3+ cells, no expression in SSEA3− cells, and weak expression of only Nanog in SFCs (Fig. 1E). Immunofluorescence staining confirmed that cell clusters from SSEA3+ cells were consistently positive for the pluripotency markers Nanog, Oct4, Sox2, and SSEA3 (Fig. 2). However, only some cell clusters from the SFCs were positive for AP staining and pluripotency markers (Fig. 2, SFCs). All cell lines had normal karyotypes at third passage.
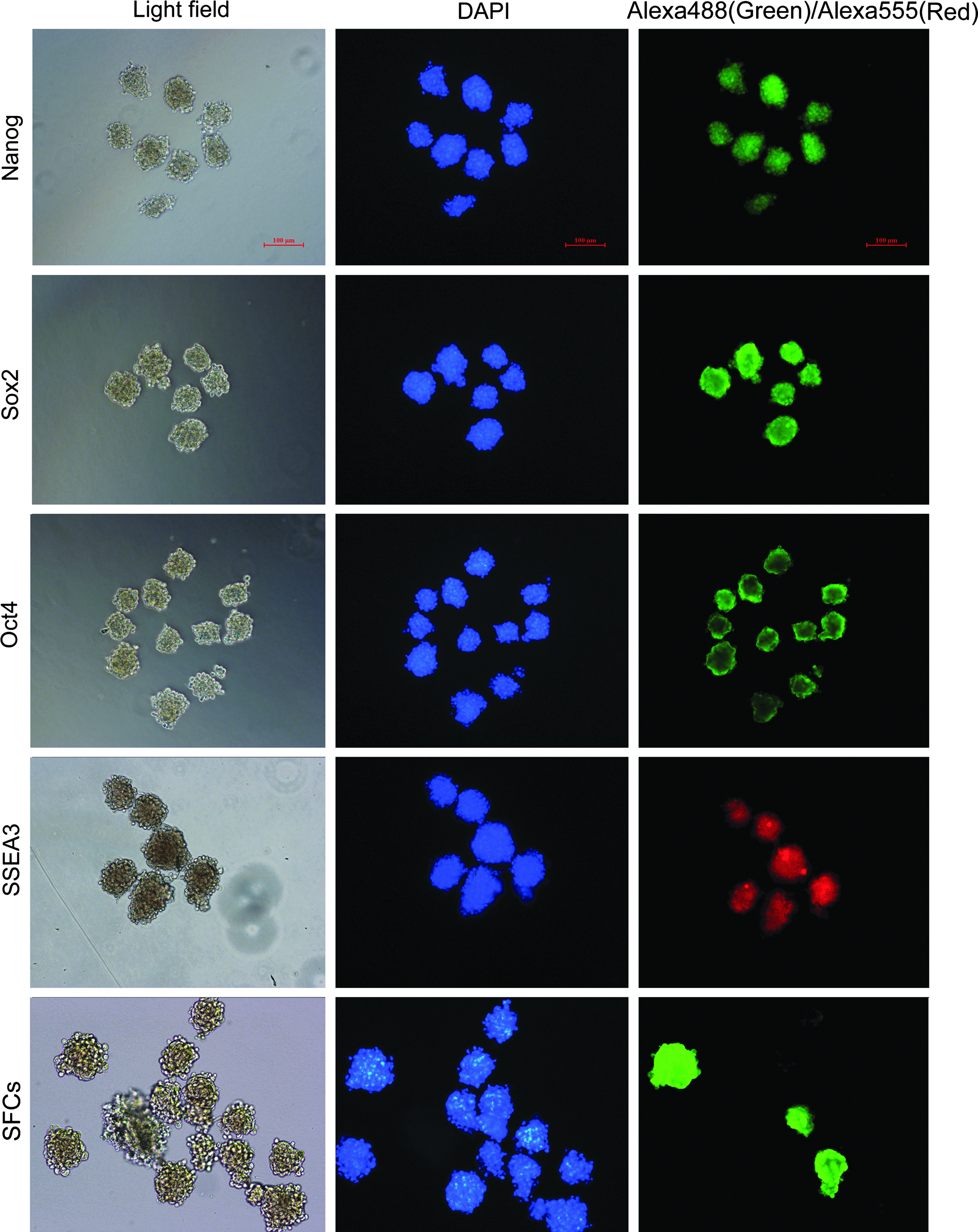
Immunofluorescence staining of cell clusters from SSEA3+ cells. Nanog, Oct4, and Sox2 were labeled by Alexa Fluor 488 (green); SSEA3 was labeled by Alexa Fluor 555 (red); DNA was stained with DAPI (blue). Immunofluorescence staining of SFCs indicated that only some cell clusters from SFCs were positive for pluripotency markers. Scale bars, 100 μm. Color images available online at www.liebertpub.com/cell
In vitro development of cloned embryos
In vitro development of cloned embryos from SSEA3+ cells is presented in Table 1. Cloned embryos from SFCs and SSEA3− cells under adherent culture were used as controls. Experiments were replicated six times. There was no difference in fusion rate among the three cell types (81.4±2.3%, 84.2±2.2%, and 82.3±1.7%, respectively). The cleavage rate in cloned embryos derived from SSEA3+ cells did not differ compared with SFCs (70.5±0.8% and 68.4±2.1%, respectively), but was significantly higher compared with SSEA3− cells (p<0.05). The percentage of fused embryos that developed to the blastocyst stage was significantly increased in the SSEA3+ cell group compared with the SFC and SSEA3− cell groups (p<0.05).
Values with different superscripts within columns are significantly different (p<0.05).
SFC, skin fibroblast cells; SSEA3+, stage-specific embryonic antigen 3–positive cells; SSEA3−, SSEA3-negative cells.
Quality of the cloned embryos
Cloned blastocyst quality from SFCs, SSEA3+, and SSEA3− cells was assessed in terms of total cell number, ICM/TE ratio, and apoptosis (Fig. 3). Total cell number, TE cell number, and ICM cell number of cloned blastocysts from SSEA3+ cells were all significantly higher compared with SFCs and SSEA3− cells, but no difference was observed for the ICM/TE ratio between the three groups (Table 2). No significant difference was observed between groups with respect to the rate of blastocyst cell apoptosis; however, blastocyst apoptotic cell numbers were lower in the SSEA3+ cell group compared to the SFC and SSEA3− cell groups (p<0.05) (Table 3). This indicated that the quality of cloned blastocysts from SSEA3+ cells was higher compared with SFCs and SSEA3− cells, based on total cell number and apoptotic cell number per blastocyst.
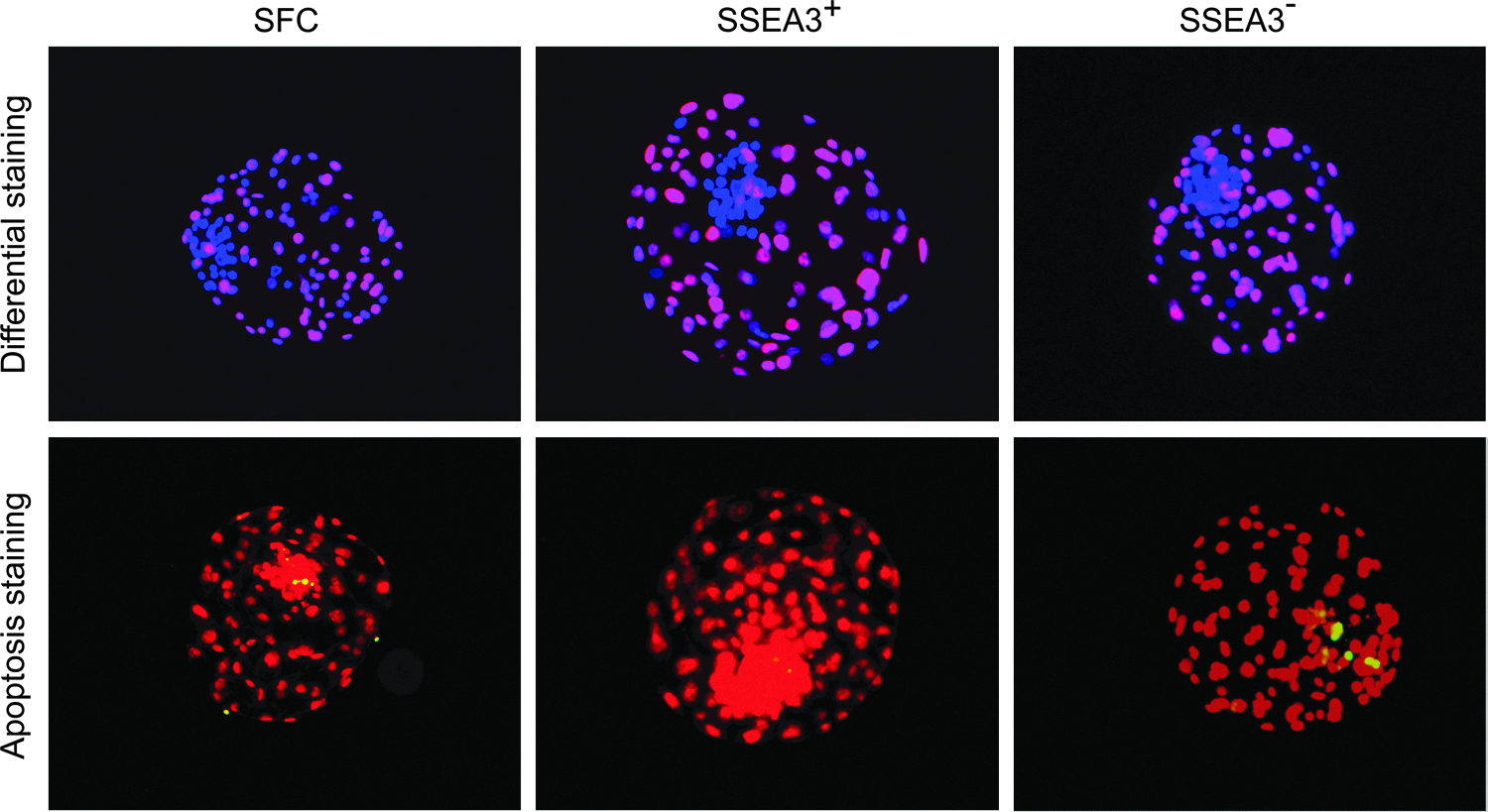
Differential staining and apoptosis assay in cloned blastocysts. Blastocyst cell numbers were estimated by counting the total number of nuclei using DAPI (blue). The number of trophectoderm (TE) nuclei was estimated using immunostaining for CDX2 (red). ICM cell number was assessed as the total number of nuclei minus the number of TE nuclei. Apoptotic nuclei were labeled by terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) (green), DNA was stained with PI (red), and co-localization is observed as yellow. Color images available online at www.liebertpub.com/cell
Values with different superscripts within columns are significantly different.
ICM, inner cell mass; TE, trophectoderm; SFCs, skin fibroblast cells; SSEA3+, stage-specific embryonic antigen 3–positive cells; SSEA3−, SSEA3-negative cells.
The percentage of embryos containing at least one apoptotic nucleus.
Average number of apoptotic nuclei per embryo.
Values with different superscripts within columns are significantly different.
SFC, skin fibroblast cells; SSEA3+, stage-specific embryonic antigen 3–positive cells; SSEA3−, SSEA3-negative cells.
Discussion
In this study, we isolated a subpopulation of cells expressing SSEA3 derived from goat SFCs. We have demonstrated that the SSEA3+ cells express pluripotency markers and enhance the in vitro development and quality of cloned embryos.
Several recent studies have demonstrated that a rare population of SSEA3+ cells exist in human dermal tissues and can be efficiently isolated in vitro from skin fibroblasts by FACS (Byrne et al., 2009; Kuroda et al., 2010; Vega Crespo et al., 2012; Wakao et al., 2011). These SSEA3+ cells are able to form cell clusters in suspension culture, which are positive for pluripotency markers and AP staining, and can differentiate into endodermal, ectodermal, and mesodermal cells, both in vitro and in vivo (Kuroda et al., 2010). These results suggested that the SSEA3+ cell is a type of adult stem cell in skin. However, there are no reports for isolating SSEA3+ cells from species other than human. In this study, we isolated and characterized SSEA3+ cells from goat SFCs. Consistent with our previous study, we demonstrated that about 2–4% of the SFCs were SSEA3+ at first passage. After suspension culture, SSEA3+ cells formed cell clusters, which were consistently positive for AP staining and the pluripotency markers Nanog, Oct4, Sox2, and SSEA3. This finding suggested that SSEA3+ cells may be found in the skin across many mammalian species. Questions regarding their location within skin tissue, cell-surface markers expression, and differentiation potential will require further study.
In general, the adult dermis contains several types of stem or progenitor cells, and the proportion of SSEA3+ cells within the cultured skin fibroblast population is very small. Expansion culture of SSEA3+ cells is important for their basic research and practical applications. However, previous studies have demonstrated that the proliferation of SSEA3+ cells is not very high and ceases in suspension culture when the cell clusters reach a defined size (Kuroda et al., 2010). Although, proliferation of SSEA3+ cells can be reinitiated by transfer to adherent culture, the proportion of SSEA3+ cells rapidly decreased to 18% after 1 week in adherent culture (Vega Crespo et al., 2012). In this study, we expended SSEA3+ cell numbers in suspension culture by adding Y-27632, a selective inhibitor of the p160-Rho-associated protein kinase (Zweigerdt et al., 2011). The result showed that 10 μM of Y-27632 enabled cell expansion greater than six passages. Further investigation is required to decipher the optimal culture conditions for SSEA3+ cell expansion.
Although cloned and transgenic goats have been produced by SCNT, the efficiency for producing live offspring is disappointingly low (Keefer et al., 2002; Liu et al., 2011; Wan et al., 2012). It has been hypothesized that undifferentiated stem cells may be easier to reprogram by recipient oocytes during SCNT. In goats, cloned embryos derived from ES-like cells showed enhanced in vitro development compared with those derived from adult fibroblast cells and lymphocytes (Dutta et al., 2011). Therefore, we expect SSEA3+ cells may be used for genetic modification and SCNT to produce transgenic animals. In the present study, we demonstrated that the blastocyst rate was higher in the SSEA3+ cell group compared with the SFC and SSEA3− cell groups (p<0.05), and cloned blastocyst quality from SSEA3+ cells was high in terms of total cell number and apoptosis. Our findings provide evidence for the hypothesis that a less differentiated cell can increase cloning efficiency. In this study however, the donor SFCs, SSEA3− and SSEA3+ cells, were cultured under different conditions, so the effect of culture condition on the development of cloned embryos can not be excluded.
In agreement with our results, NT using porcine skin stem cells (Zhu et al., 2004), porcine MSCs (Faast et al., 2006; Kumar et al., 2007), porcine NSCs (Zheng et al., 2009), and mouse NSCs (Inoue et al., 2007) exhibits enhanced development of cloned embryos. Conversely, reports indicate that mouse hematopoietic stem cells (Inoue et al., 2006; Sung et al., 2006), bovine and porcine MSCs (Colleoni et al., 2005), and mouse NSCs (Mizutani et al., 2006) show a similar or lower developmental rate comparable to somatic cells. Therefore, the use of undifferentiated adult stem cells as suitable donor cells warrants further investigation. Previous studies have indicated that both the differentiation and epigenetic state of the donor nucleus influence the reprogramming efficiency of cloned embryos (Blelloch et al., 2006). However, the ideal epigenetic state of the donor cell to facilitate efficient reprogramming following SCNT remains unclear. In this study, SSEA3+ and SSEA3− cells were isolated from a skin fibroblast cell line and demonstrated differential reprogramming potential following SCNT. Therefore, comparison analysis between SSEA3+ and SSEA3− cells may help elucidate the mechanisms of reprogramming following SCNT.
In conclusion, our results suggest that SSEA3+ cells from goat SFCs form cell clusters in suspension culture and consistently express the pluripotency markers, and that using SSEA3+ cells as donor cells can enhance the in vitro development and the quality of cloned embryos.
Footnotes
Acknowledgments
We thank Mr. Jintao Hu for technical assistance in using FACS. This work was supported by the National Major Project for Production of Transgenic Breeding (No. 2011ZX08007-004) and the National High Technology Research and Development Program of China (863 Program) (No. 2011AA100303).
Author Disclosure Statement
The authors declare no conflicts of interest exist.