
Abstract
Abstract
In this study, the effect of the steps involved in zona-free somatic cell nuclear transfer (SCNT) on oocyte transcripts was investigated in sheep. To establish the reliable combined electrical–chemical activation for zona-free oocytes, oocytes were first exposed to an electrical pulse and then treated with 18 chemical activation regimens designed through modifying duration and concentration of ionomycin and 6-dimethyl aminopurine (6-DMAP), which is routinely used for SCNT. Electrofusion-mediated nuclear transfer significantly reduced transcript abundances of CCNB1, POU5F1, NPM2, GMMN, and CX43 compared to intact oocytes. Maximum parthenogenetic blastocyst development was obtained when oocytes were submitted to electric pulse and then to (1) 5 μM ionomycin for 5 or 2.5 min, both followed by 2 h of incubation with 6-DMAP (41.7±1.1, and 42.4±1.4%, respectively), (2) 5 μM ionomycin for 1 min+6-DMAP for 4 h (43.1±1.4%), and (3) 2.5 μM ionomycin for 1 min+6-DMAP for 2 h (42.4±1.4%), with significant differences compared to all the other groups. Statistical assessment of interactions between duration and concentration of ionomycin and duration of 6-DMAP exposure revealed that (1) concentration of ionomycin may be a more important factor than its duration, (2) both a long exposure period and a low concentration of ionomycin had marked decreasing effects on parthenogenetic development of zona-free oocytes, and (3) high duration of exposure to 6-DMAP can reduce parthenogenetic development. Despite an activation preference of parthenogenetic oocytes, a significantly higher rate of cloned blastocyst development was observed when reconstructed oocytes were activated with 5 μM ionomycin for 5 min rather than 2.5 μM ionomycin for 1 min (8.8±2.5 vs. 1.25±2.2%). These results suggested that SCNT steps have determining effects on oocyte transcripts and activation preferences of the reconstituted oocytes compared to intact counterparts. In this sense, reconstituted oocytes may need a higher concentration of ionomycin for a longer period than intact oocytes.
Introduction
Ionomycin is the best-known AOA chemical (Hosseini et al., 2008a). Mechanistically, ionomycin is a small hydrophobic molecule that provokes a monotonic rise in intracellular Ca2+ via stimulating capacitative Ca2+ entry (CCE) in the oocytes through depletion of both endoplasmic reticulum stores and enhancement of Ca2+ influx (Macháty et al., 2002; McGuinnes et al., 1996). However, while almost all species used nearly the same chemical activation protocol, a growing body of evidence indicates that oocytes of each species may respond differently to a given activation protocol (Asgari et al., 2012; Lan et al., 2005; Nasr-Esfahani et al., 2011), and therefore, species-specific modifications of this standard procedure may have important impacts on subsequent development of activated oocytes.
Importantly, the MII-stage oocyte is very sensitive and responsive to stimuli, and comparative studies have shown that all in vitro oocyte manipulation processes may have the potential to trigger partial oocyte activation, and therefore may change the activation threshold of the manipulated oocytes (Hosseini et al., 2012). However, there is no information regarding whether the different steps involved in SCNT may affect activation requirements of the reconstituted oocytes. Furthermore, while electropulses (EP) used for nuclear transfer are of enough power to induce AOA, a limited number of available studies have considered EP in primary parthenogenesis studies along with chemical activation protocols (Du et al., 2005; Hosseini et al., 2008a). Therefore, although one may consider that the activation requirements of cloned oocytes may vary compared to intact oocytes, it is important to evaluate these activation requirements before and after SCNT. In this sense, this study was carried out to investigate the effect of SCNT process on transcript abundances and activation requirements of the reconstituted oocytes.
Materials and Methods
Unless otherwise specified, all chemicals and media were obtained from Sigma Chemical Co. (St. Louis, MO, USA) and Gibco (Grand Island, NY, USA), respectively.
Oocyte preparation and in vitro maturation
The process of oocyte maturation was as described previously (Hosseini et al., 2013). In brief, sheep ovaries were collected at a local abattoir and transported in 0.9% NaCl solution at 28–32°C to the laboratory within 2–3 h. Cumulus–oocyte complexes (COCs) were aspirated from transparent 2- to 6-mm follicles using 20-gauge needles attached to a vacuum pump (80 mmHg), and the follicle content was released in preincubated HEPES-buffered tissue culture medium-199 (TCM-199) (H-TCM), supplemented with 10% fetal calf serum (FCS) and 50 IU/mL heparin. Oocytes with at least three layers of cumulus cells and with a uniform granulated cytoplasm were selected for the experiments. The oocytes were cultured in medium comprised of TCM plus 10% FCS, 2.5 mM sodium pyruvate, 100 IU/mL penicillin, 100 μg/mL streptomycin, 10 μg/mL follicle-stimulating hormone (FSH), 10 μg/mL luteinizing hormone (LH), 1 μg/mL estradiol-17β, and 0.1 mM cysteamine under mineral oil for 20–22 h at 39°C, 5% CO2, and maximum humidity. Oocytes were stripped of surrounding cumulus cells by manual pipetting in 0.1% hyaluronidase. Denuded oocytes were then rested for 15 min before being subjected to zona removal. For this purpose, oocytes were treated with 2.5 mg/mL Pronase (Sigma protease, P 8811) prepared in H-TCM+10% FCS. Zona-free oocytes were washed and incubated in H-TCM+20% FCS to completely remove the enzyme.
Experimental design
Three experiments were conducted in this study. In experiment I, the effect of zona removal and two main steps of SCNT (enucleation and nuclear transfer) on relative abundances of eight developmentally important genes (CCNB1, POU5F1, PAP, CX43, GMNN, NPM2, ATP1A1, and HSP90) and two housekeeping genes (GAPDH and ACTB) were studied and compared with the transcript profile of zona-intact oocytes. In experiment II, the effects of different oocyte activation protocols on parthenogenetic development of zona-free oocytes were assessed. In this sense, through modifying duration and concentration of ionomycin and 6-DMAP, which are routinely used for SCNT, a systematic study using 18 different protocols was designed to determine the optimal activation protocol (Fig. 1). As a control activation group, the most routine activation regime used for parthenogenetic activation (5 μM ionomycin for 5 min followed by 2 mM 6-DMAP for 4 h) was used. Because we could not practically carry out 18 groups simultaneously, a control group was included in each run and the data were presented as the difference from the control group (see Statistical Analysis section). In experiment III, the effects of the best effcient activation protocols (selected based on experiment II) on in vitro development of zona-free reconstructed oocytes were investigated.
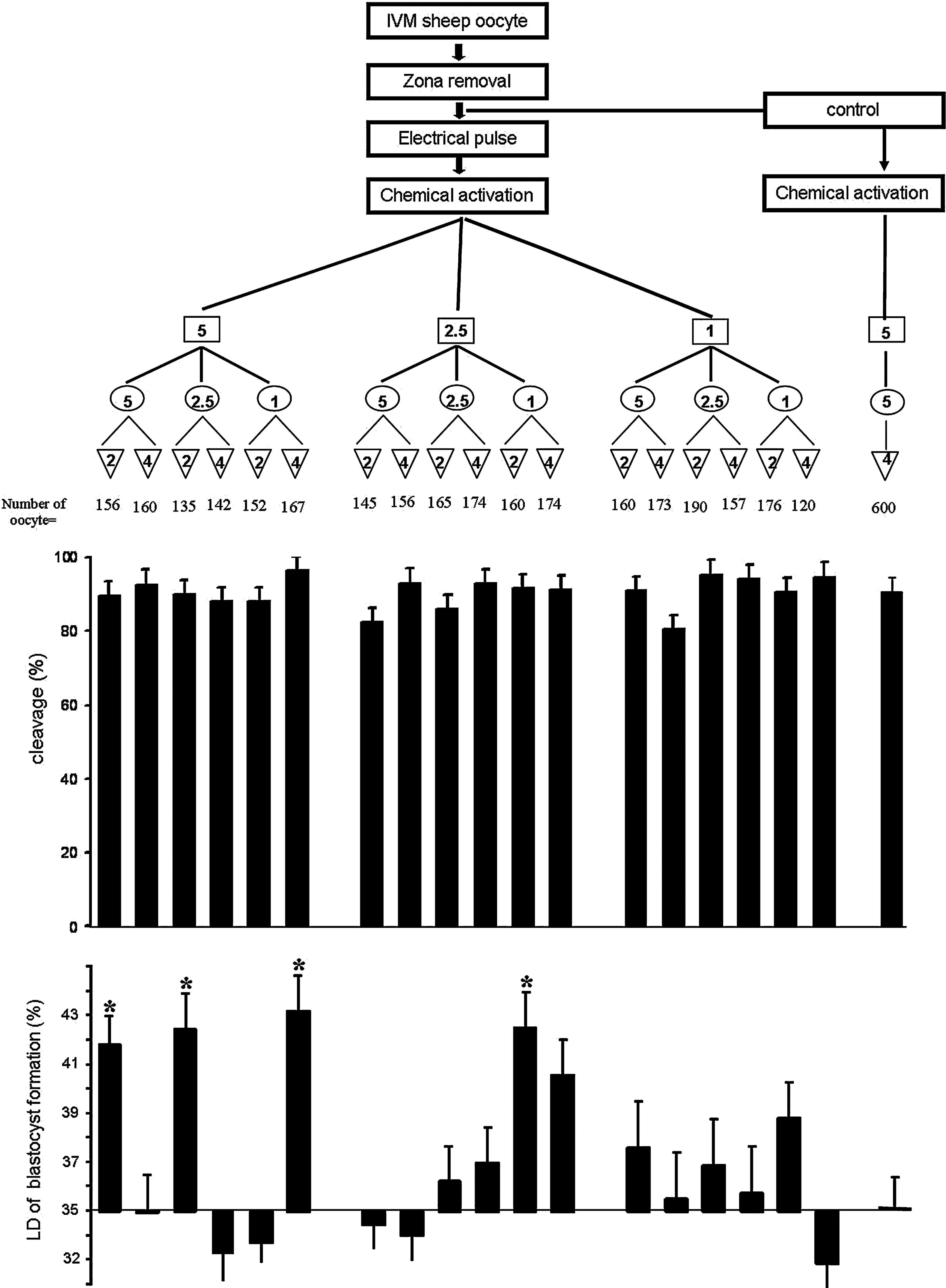
Experimental design of the parthenogenetic activation treatments used for zona-free oocytes and show cleavage rate and the level of difference (LD) for treatments relative to control at blastocyst stage. The LD was obtained by subtracting the value of each treatment group from its related control (common protocol, of 5 μM ionomycin for 5 min then incubation with 2 mM of 6-DMAP for 4 h without electrical pulse). Asterisks indicate significant difference between treatment groups and control group (p≤0.05). (Open squares) Ionomycin concentration ( μM); (open circles) ionomycin duration (minute); (inverted open triangles) 6-DMAP duration (hour).
Experiment I: Effect of SCNT steps on relative mRNA abundances of oocytes
For this purpose, 30 min after zona removal, enucleation, and nuclear transfer, oocytes were used for quantitative real-time PCR (qPCR), as described previously (Hosseini et al., 2012). In brief, a pool of 30 oocytes in each replicate of each experiment were pooled and washed twice in phosphate-buffered saline (PBS), and total RNA from each sample was extracted using the RNeasy Micro Kit (Qiagen®) according to the manufacturer's instructions. The concentration of extracted total RNA was determined by measuring the absorbance at 260 nm in a spectrophotometer. cDNA was synthesized by using a Takara™ First Strand cDNA Synthesis Kit. The quantification of the transcripts was determined by using 25 ng of cDNA, 10 μL of SYBR Green/ROX qPCR Master Mix (2×; Fermentas), and 1 μL of the forward and reverse primers (5 pmol) adjusted to a total volume of 20 μL using nuclease-free water. MII zona-intact oocytes were considered as controls. Primer sequences, annealing temperatures, and the size of amplified products are shown in Table 1. In this study, GAPDH and ACTB were considered as housekeeping genes; by comparing the expression of GAPDH and ACTB to the cDNA template of the experiments, we did not observe important differences in expression levels of these two housekeeping genes for the same groups (data not shown).
F, forward; R, reverse.
Experiment II: Effect of different activation protocols on developmental competence of zona-free oocytes
As shown in Figure 1, zona-free oocytes at 22 h postmaturation (hpm) were first incubated in fusion medium (0.28 M mannitol in 290 mOsm fusion buffer free of Ca2+ and Mg2+) for 30 sec, and then carefully placed between the platinum wires (0.5 mm apart) of the fusion chamber filled with fusion medium. The oocytes were then exposed to two direct currents (1.75 KV/cm for 30 μsec and 1 sec delay) of electric pulse. This pulse setting was selected based on previous experiments carried out for electrofusion of somatic cells to the enucleated oocytes in SCNT experiments (Hosseini et al., 2008a). The electropulsed zona-free oocytes were exposed to three different concentrations (5, 2.5, and 1 μM) and durations (5, 2.5, and 1 min) of ionomycin treatment followed by incubation with 2 mM of 6-DMAP for 2 or 4 h (Fig. 2). Activated zona-free oocytes were cultured in groups of six in wells of the wells (WOWs, 20 μL) (Vajta et al., 2000) in a modified formulation of synthetic oviductal fluid (mSOF) (Hosseini et al., 2008b). Cleavage and blastocyst rates were determined on days 3 and 8 postactivation, respectively.
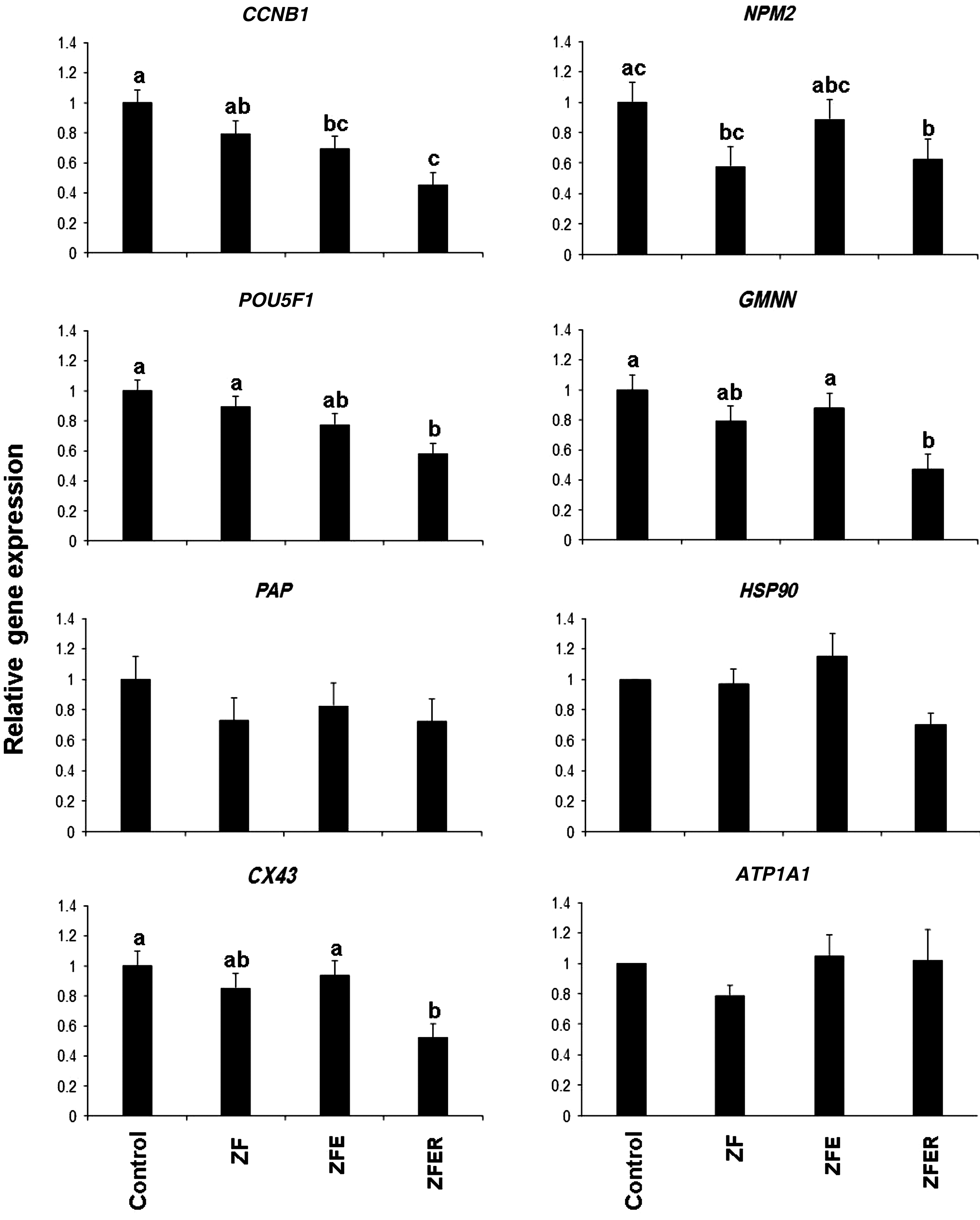
Quantitative analysis of mRNA abundances of Cyclin-B, Oct4, PAP, Cx43, NPM2, GMNN, HSP90, and ATPase in zona-free (ZF), zona-free enucleated (ZFE) zona-free enucleated-renucleated (ZFER) and zona-intact (control) MII sheep oocyte. Values with at least one common letter are not significantly different at p≤0.05.
Experiment III: Effect of selected activation protocols on developmental competence of cloned embryos
During the experiment III, because it was difficult to include all experimental groups of experiment II, we selected the two best activation protocols determined during experiment II and used them for activation of zona-free SCNT oocytes. As a control, the routine process of AOA (5 μM ionomycin for 5 min followed by 4 h of incubation with 2 mM 6-DMAP) was also considered for control zona-free oocytes. The process of zona-free SCNT was carried out as described previously (Nasr-Esfahani et al., 2011). For enucleation, zona-free oocytes were incubated in H-SOF containing 3 mg/mL polyvinyl alcohol (PVA) and 5 mg/mL H33342 for 5 min. Oocytes were then washed and transferred into HEPES-buffered (H-SOF) plus PVA and 10% FCS droplets on the microscope stage (Olympus; IX71, Japan) equipped with Narishige micromanipulators (Olympus, Japan). Using 100× magnification, oocytes were enucleated under constant ultraviolet (UV) exposure (Oback et al., 2003). As soon as the chromosome mass (under UV light) was aspirated into the enucleation pipette (10–15-mm outer diameter, perpendicular break), the cytoplast and karyoplast were separated with a brief kick by hand on the warm stage. For nuclear transfer (NT), trypsinized donor cells were suspended in H-SOF with 0.5% FCS in 5-mL centrifuge tubes. Through making serial dilutions in three to four droplets, a population of 10–20 individual cells was added to a drop of 10 mg/mL phytohemagglutinin (PHA-P) in H-SOF. Oocytes were then picked up individually and gently pushed over a single cell, as the oocyte rolled upon the cell. For electrofusion, 10–15 reconstructed oocytes were first equilibrated for up to 2 min in isotonic (290 mOsm) fusion buffer. A total of 10–15 couplets were automatically aligned using alternating current before being subjected to two direct currents as described for parthenogenetic activation. Successfully fused couplets were activated using the selected activation protocols determined in experiment II. Reconstructed oocytes then cultured and devaluated as described for parthenogenetic embryos.
Statistical analysis
In the first experiment, one-way analysis of variance (ANOVA) was used for parametric analysis of differences of transcripts abundances between zona-intact, zona-free, and reconstructed oocytes (SPSS 15). In experiment II, the percentage of blastocysts in parthenogenetic activation, transformed to level of difference (LD) from the control before analysis. The level of difference was obtained by subtracting the value of each group from its related control. Therefore, for each set of treatments, a common activation protocol was considered as control group (5 μM ionomycin for 5 min and 4 h of 6-DAMP). To study the effect of different variables, a factorial experiment was carried out using SAS software (SAS 9.1). In this model, the three variables included were concentration of ionomycin (three levels: 5, 2.5, and 1 μM), duration of ionomycin (three levels: 5, 2.5, and 1 min) and 6-DMAP (two levels: 4 and 2 h), and their interactions. The differences among means were analyses using the Tukey multiple comparison test. In experiment III, in vitro development data were examined using a one-way ANOVA followed by Tukey multiple comparison post hoc tests using SPSS15. All experiments in this study were repeated at least three times, and the differences were considered to be significant at p≤0.05.
Results
Effect of SCNT process on transcripts abundances
As shown in Figure 2, for all genes assessed, neither zona removal nor enucleation significantly affected the relative abundances of transcripts, except for RNA abundance of CCNB1, which was significantly reduced following enucleation compared to zona-intact oocytes. However, transcript abundances of CCNB1, POU5F1, NPM2, GMMN, and CX43 were all significantly reduced after electrofusion of enucleated oocytes with the nuclei donor cells. Unlike other genes, the reduction in abundances of PAP, ATP1A1, and HSP90 was not affected by the SCNT steps.
Effect of combined electrical–chemical activation protocol on developmental competence of zona-free sheep oocytes
Overall, the cleavage rates ranged from 80.5±3.9 to 96.3±3.9% with the mean of 90.4±3.9%. Blastocyst rates also ranged from 30.82±3.1 to 42.47±3.1% with the mean of 37.27±3.1%. As shown in Figure 1, comparison of cleavage rate between different protocols revealed no significant difference. However, blastocyst rates were affected (p≤0.05) by the activation protocol, and the highest blastocyst rates compared to control (35±1.1%) were obtained in the following groups: (1) EP+5 μM ionomycin for 5 min+6-DMAP for 2 h (41.7±1.1%); (2) EP+5 μM ionomycin for 2.5 min+6-DMAP for 2 h (42.4±1.4%); (3) EP+5 μM ionomycin for 1 min+6-DMAP for 4 h (43.1±1.4%); and (4) EP+2.5 μM ionomycin for 1 min+6-DMAP for 2 h (42.4±1.4%). The lowest blastocyst rate was observed in EP+1 μM ionomycin for 1 min+6-DMAP for 4 h (30.82±1.1%), which was significantly lower compared to four aforementioned groups (Fig. 1).
Effects of concentration of ionomycin and duration of ionomycin and 6-DMAP on activation of zona-free ovine oocytes
To evaluate the independent effect of each factor (concentration of ionomycin, duration of ionomycin, and duration of 6-DMAP), two factors were considered as constants (i.e., nonvariable) and the effect of third factor was evaluated. Therefore, when duration of exposure to ionomycin and 6-DMAP was considered as constant, the difference in blastocyst rate compared to control was significantly decreased with reducing concentration of ionomycin. Therefore, the blastocyst rates obtained with 5 μM and 1 μM ionomycin were significantly different (Fig. 3A). Similarly, when concentration of ionomycin and duration of exposure to 6-DMAP were considered as a constant, increased duration of exposure to ionomycin negatively affected the difference in blastocyst rate compared to control. Therefore, the values of these parameters were significantly different between 1-min and 5-min exposure groups (Fig. 3B). Likewise, when concentration and duration of ionomycin were considered as constant, increased duration of 6-DMAP exposure (2 h vs. 4 h) significantly reduced blastocyst rate compared to control (Fig. 3C).
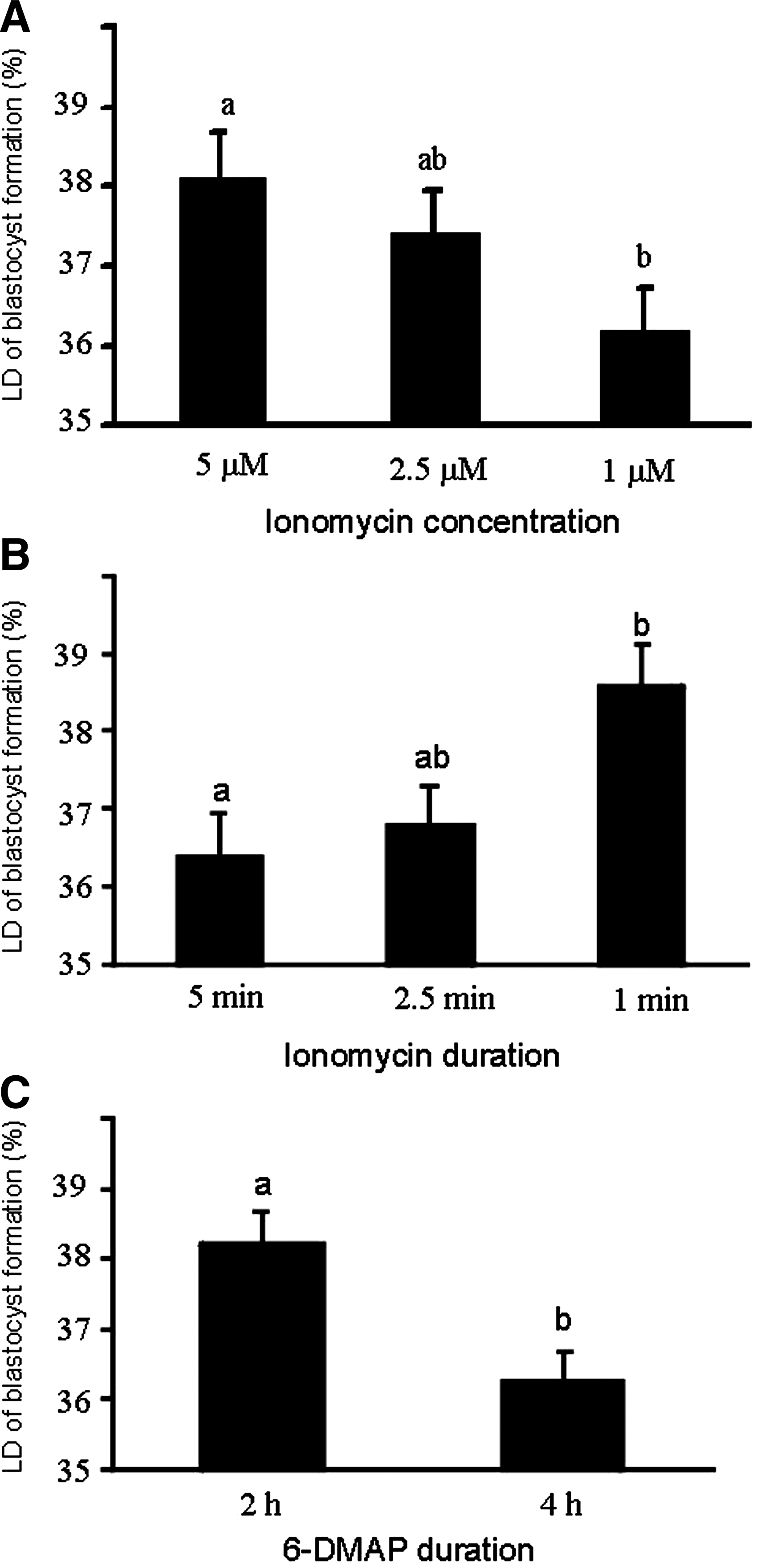
Effect of ionomycin concentration (
Evaluation of interactions effects between concentration of ionomycin, duration of ionomycin, and duration of 6-DMAP
To evaluate the interactions between each parameter, one parameter was considered as constant and the effect of the other two variables with each other were considered. When the duration of ionomycin was considered as constant, the results (Fig. 4A) showed that increased duration of exposure to 6-DMAP at low (1 μM) concentration of ionomycin significantly reduced the difference in blastocyst rate compared to control. Similarly, when concentration of ionomycin was considered as constant, increased duration of exposure to both ionomycin and 6-DMAP significantly decreased blastocyst formation compared to control (Fig. 4B). However, when the duration of 6-DMAP exposure was considered constant (Fig. 4C), the interactions between different concentrations and durations of ionomycin did not show a meaningful trend. Figure 4C shows that 2.5 μM ionomycin for 1 min had the potential to be tested for activation of zona-free reconstituted sheep oocytes.
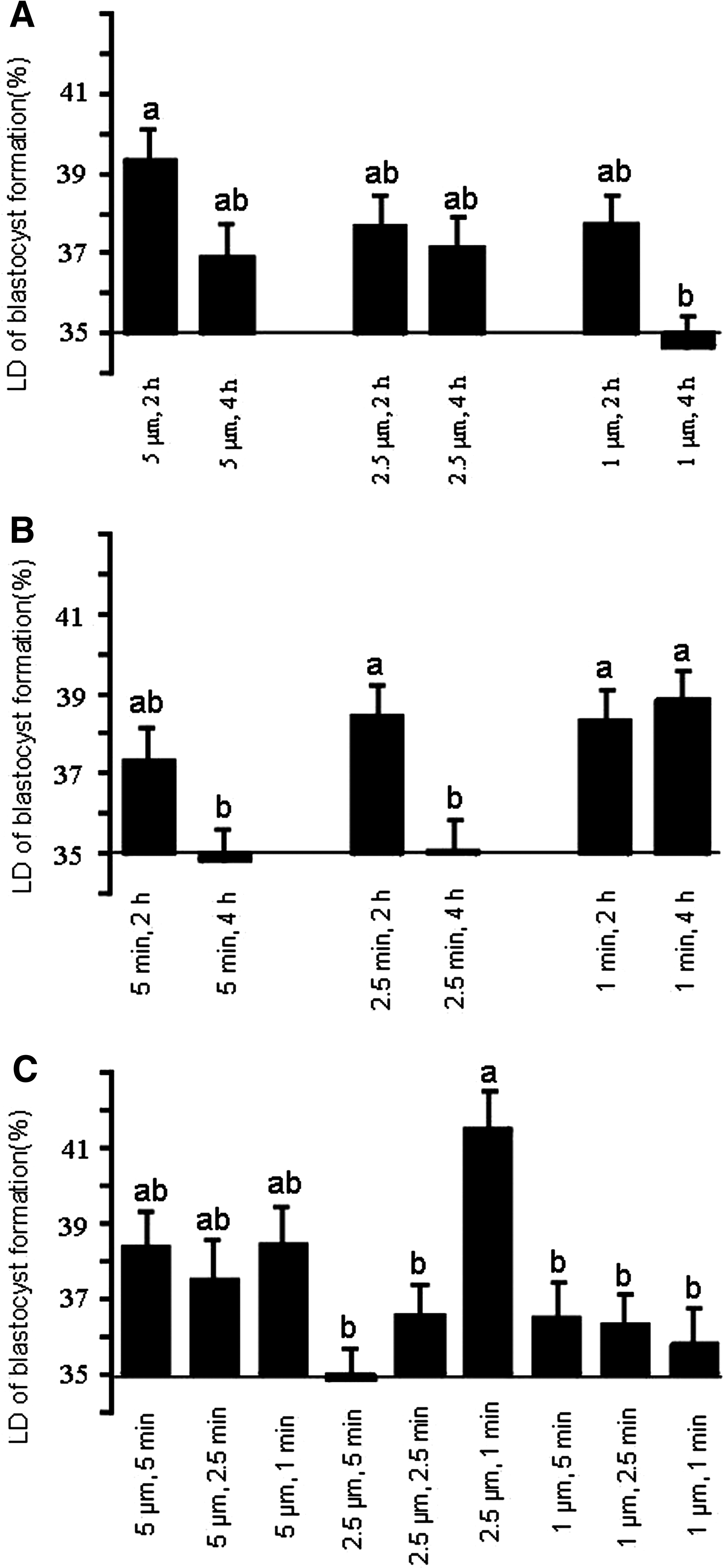
Effect of interaction between ionomycin concentration and 6-DMAP duration (
Effect of different activation protocols on in vitro development rates of zona-free reconstituted sheep oocytes
On the basis of the results of oocyte activation in experiment II, we selected two oocyte activation regimes (5 μM ionomycin for 5 min and 2.5 μM ionomycin for 1 min both followed by 2 h treatment with 6-DMAP) for activation of the reconstituted sheep oocytes. As shown in Figure 5, cleavage rates of the reconstituted oocytes activated with 5 μM ionomycin for 5 min were not significantly different from those reconstituted oocytes treated with 2.5μμM ionomycin for 1 min (79.46±7.2 vs. 90.6±2.9%). However, blastocyst yield of the reconstituted oocytes in the former group (8.8±2.5%) was significantly higher than the latter protocol (1.25±2.2%).
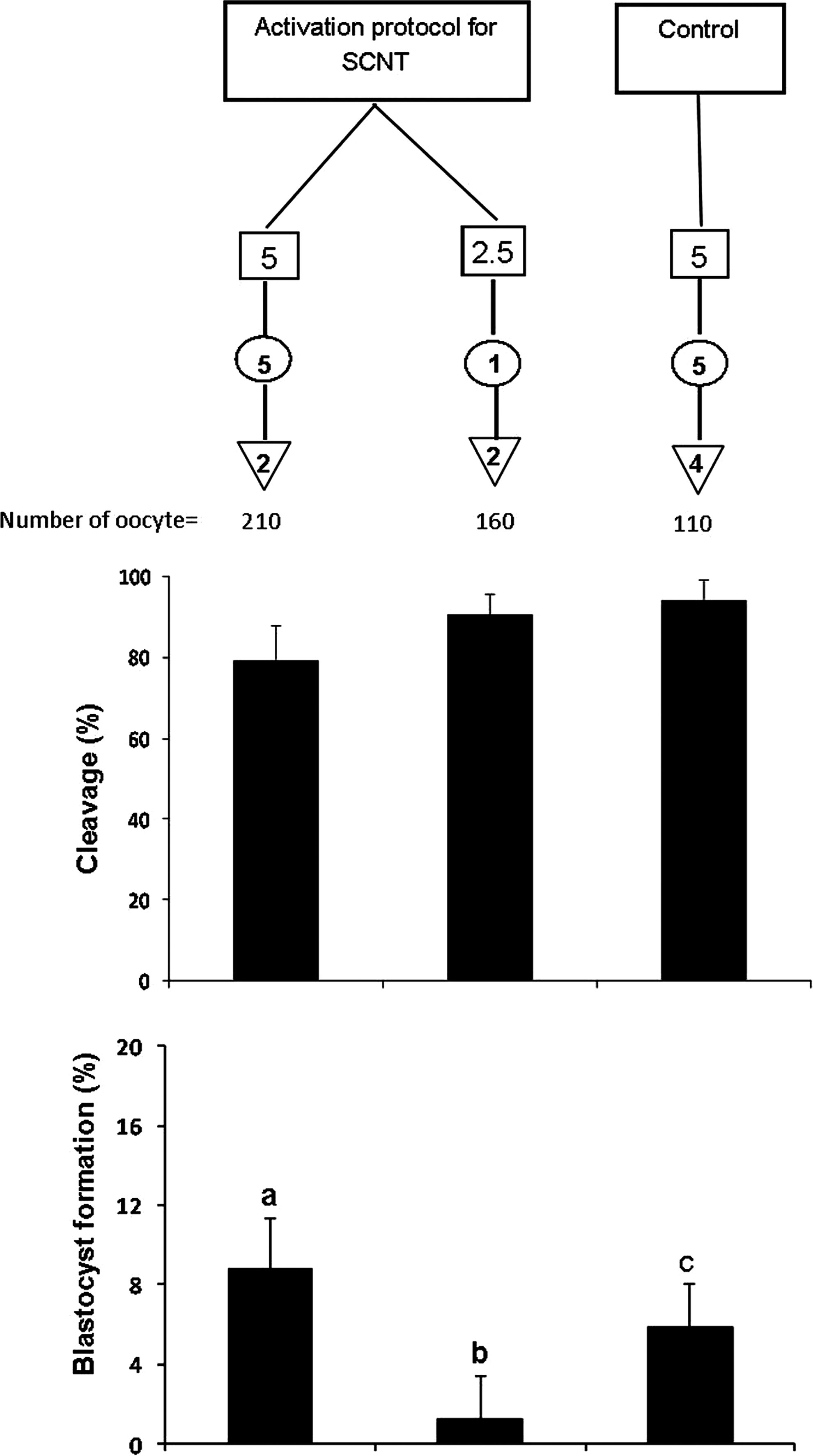
Effect of activation methods on cleavage and blastocyst rate of zona-free sheep oocytes in SCNT process. (Open squares) Ionomycin concentration ( μM); (open circles) ionomycin duration (minute); (inverted open triangles) 6-DMAP duration (hour). Values with at least one common letter are not significantly different at p<0.05.
Discussion
Cloning steps and oocyte transcripts
The oocyte is a unique cell committed to reprogramming fertilizing sperm and to supporting early stages of embryonic development until the species-specific stage of zygote genome activation (Sirard, 2010) that occurs, for example, around the second to third cell cycle in sheep embryos (Hosseini et al., 2012). In this sense, and considering the huge list of oocyte transcripts, we selected some candidates genes based on their roles of regulating different cell functions described thereafter. The results obtained provided indications that the steps involved in a zona-free method of SCNT can influence the oocyte transcripts. In general, the relative amounts of all genes were affected by all steps (zona removal, enucleation, renucleation), but these effects were further aggravated by reconstruction mediated by electrofusion pulses. The exact mechanism by which zona-removal affected mRNA abundances is not clear, but it is interesting to note that Ribas et al. (2006b) observed that zona removal significantly reduced DNA methylation in the resultant mouse embryos at early stages. They showed that “Removal of the zona pellucida immediately after recovery of fertilized eggs caused a significant reduction in DNA methylation at the two- and four-cell stage in mouse embryos.” Among RNAs that were not affected by SCNT steps is PAP, a RNA essential for extension of the poly(A) tail to activate RNA and a key regulatory step in gene expression in early embryonic development (Seli et al., 2005). The RNA contents that were just affected by the reconstruction procedure were NPM2, POU5F1, CX43, and GMNN, which are involved in pluripotency (Vigneault et al., 2009), gap junction communication and regulation mechanisms (Pant et al., 2005), DNA replication, and reprogramming during early embryogenesis (Sylvestre et al., 2010), respectively.
Geminin (GMNN) is an important gene that is highly expressed in the oocyte. GMNN inhibits DNA replication and activates SOX2 during neural development via interactions with chromatin remodeling complexes (Papanayotou et al., 2008). Importantly, Gonzalez et al. (2006) showed that lack of GMNN during mammalian development has an important role in cell differentiation during early embryonic development. It is important that the expression levels of GMNN in bovine parthenogenetic embryos were lower than in the control in vitro fertilization (IVF) in the study of Sekhavati et al. (2012). Nucleoplasmin 2 (NPM2) is a major regulator of sperm chromatin decondensation (Burns et al., 2003; Tamada and Thuan, 2006). Importantly, using a novel approach, Betthauser et al. (2006) supplemented the recipient oocytes with NPM2 by direct injection into bovine oocytes and observed that over 200 genes were upregulated in these injected SCNT-derived oocytes. Almost of these genes were those that have previously been shown to be downregulated in SCNT embryos compared to fertilized ones. Expression levels of CCNB1 are considered to be an indirect indication of maturation/M-phase promoting factor (MPF) activity (Kikuchi et al., 2000). Therefore, the decrease in expression of CCNB1 following zona removal and enucleation–renucleation results in MPF decrease. Lee and Campbell (2006) reported that enucleation, which removes small volume of cytoplasm, may reduce MPF and mitogen-activated protein kinase (MAPK) activities. However, a reduction in MPF activity due to enucleation may be species-specific, because studies on porcine oocytes have failed to show a decline in MPF associated with the enucleation procedure (Goto et al., 2002). MPF is considered as the main inhibitor of meiosis resumption and the MPF level is related to activation threshold of MII-arrested oocytes (Asgari et al., 2012). In this sense, one may argue that decreased levels of CCNB1 (and hereby MPF) and NPM2 in reconstituted versus intact oocytes may account for higher sensitivity of zona-free oocytes to oocyte activation and can provide a platform on which to conclude that reconstituted oocytes may need special activation requirements different from those of intact oocytes. This is in agreement with other studies demonstrating that zona-free oocytes are more sensitive and responsive to artificial activation than zona-intact ones, and so modifications may be needed in both the concentration and duration of treatments (Vajta et al., 2003). Moreover, considering the role of MPF, NPM2, POU5F1, and GMMN in nuclear remodeling and reprogramming events (Sylvestre et al., 2010), one may argue that a decrease in these transcripts also accounts for the reduced reprogramming capacity of oocytes and, hence, lower blastocyst yield rate of SCNT versus parthenogenetic embryos, the points that are discussed below. To this should be added also the effect of UV exposure of oocyte for aspiration of the MII spindle. Indeed, while we did not evaluate this potential effect, a number of studies have criticized the detrimental effect of UV-assisted enucleation on oocyte developmental competence (Gil et al., 2012; Versieren et al., 2012). In this sense, emerging evidence indicates that the SCNT procedure that eliminates the essential need of UV in oocyte enucleation may improve cloning efficiency (Gil et al., 2012; Hosseini et al., 2013). For example, we have recently developed a manual method of oocyte enucleation using demecolcine (which induces a clearly visible cytoplasmic protrusion) (Hosseini et al., 2013). This method is not only considered as an alternative to UV-based methods of oocyte enucleation but also brings feasibility and safety to oocyte enucleation.
Interaction effects of concentration and duration of ionomycin and duration of 6-DMAP on parthenogenetic activation of zona-free sheep oocytes
The results presented in Figure 1 indicate that despite the fact that parthenogenetic blastocyst development differences from control were significant in some groups, the presence of three variables (concentration and duration of ionomycin and duration of 6-DMAP) make the conclusion regarding the exact contribution of each variable on embryo development difficult. Therefore, during the analysis, it was decided to consider one or two of the variables as constant and the results were revaluated (Figs. 3 and 4). These comparisons revealed that despite no difference in cleavage, the blastocyst rates were reduced with decreasing concentrations of ionomycin from 5 to 2.5 μM and then 1 μM (Fig. 3A). However, when the concentration was considered as constant, maximum blastocyst yield of zona-free oocytes was obtained when the duration of exposure to ionomycin was reduced to 1 min (Fig. 3B). Therefore, these results provided a platform on which to conclude that (1) overall, the concentration of ionomycin may be a more important factor than its duration, and (2) both long exposure period and low concentration of ionomycin had marked decreasing effects on parthenogenetic development of zona-free oocytes.
When the duration of ionomycin was considered constant (Fig. 4A), it was observed that prolonged (4 h) exposure to 6-DMAP has a nonbeneficial, even detrimental, effect on parthenogenetic development. The same held true when the concentration of ionomycin treatment was considered constant (Fig. 4B). Therefore, a second conclusion was that high duration of exposure to 6-DMAP can reduce parthenogenetic development, and, as a consequence, the duration of 6-DMAP treatment was suggested to be kept at a minimum (2 h) for parthenogenetic activation of zona-free oocytes. Although the overall trend of our previous study on zona-free goat oocytes (Nasr-Esfahani et al., 2011) is similar to the present study, in this previous study we did find a difference between the parthenogenetic development of oocytes incubated with 6-DMAP for 2 or 4 h. However, in agreement with our present results, it has been suggested that shorter exposure to 6-DMAP (Lan et al., 2005) (4 h vs. 2 h) results in a higher blastocyst formation rate, because prolonged exposure to 6-DMAP can contribute to compromised spindle rotation and small pronuclei formation (Leal and Liu, 1998; Szollosi et al., 1993), which subsequently impairs embryo development and SCNT outcome.
On the basis of the literature, two reasons can be considered for a milder activation protocol requirement of zona-free oocytes, not only in sheep (present study), but also in goat (Nasr-Esfahani et al., 2011) and bovine (Vajta et al., 2003) oocytes, including: (1) Preactivation state of ovine and caprine zona-free oocytes, due to their observed propensity to partially extrude their nuclear material as a cortical protrusion and progress toward the anaphase–telophase II stage (Baguisi et al., 1999; Chen et al., 2007; Nasr-Esfahani et al., 2011), which needs further investigation, and (2) increased susceptibility of zona-free oocytes to activation triggering and enhanced resilience of the oolemma to ion exchange, which also enhances fusion of membranes (Booth et al., 2001; Zimmermann and Vienken 1982).
Activation requirements of zona-free oocytes may change after SCNT
On the basis of the results of parthenogenetic activations in experiment II, two activation protocols (5 μM ionomycin for 5 min, 2.5 μM ionomycin for 1 min, both followed by 2 h treatment with 6-DMAP) were used for activation of zona-free reconstituted oocytes. Importantly, contrary to our expectations, a significantly higher rate of blastocysts was observed when reconstructed oocytes were activated with 5 μM ionomycin for 5 min rather than 2.5 μM ionomycin for 1 min (which have been resulted in relatively highest rate of blastocyst formation in experiment II).
Mechanistically, appropriate activation of matured oocytes requires the intracellular Ca2+ concentration to increase above a certain threshold level (Alberio et al., 2001; Macháty et al., 2002), which can be induced by certain concentrations and durations of ionomycin shown here for parthenogenetic activation of zona-free oocytes. Importantly, during SCNT, oocytes are exposed to a high degree of manipulation, and this may result in leakage of Ca2+ from the endoplasmic reticulum along with influx of external Ca2+(Tian et al., 2007), which may leave reconstituted oocytes nonresponsive to the low activation stimuli. Therefore, one may conceive that these reconstituted oocytes may need a higher concentration of ionomycin for a longer period than that intact oocytes need.
The rate of blastocyst observed with both of these protocols was consistently higher than the rate of the routine AOA protocol (5 μM ionomycin for 5 min followed by a 4-h exposure to 6-DMAP). Alexander et al. (2006) demonstrated that oocyte activation agents such as 6-DMAP and cycloheximide (CHX) have differing effects on meiotic or mitotic nuclei. Their results showed that 6-DMAP may act in a different way in the activation of parthenogenetic embryos compared to SCNT embryos.
Conclusion
The zona-free SCNT procedure, despite being a simple, fast, and efficient procedure compared to conventional cloning procedures, requires considerable modifications due to removal of the zona (Oback et al., 2003; Ribas et al., 2005; Vajta et al., 2003), and, more importantly, particular species-specific modifications may be needed (Ribas et al., 2006a). This study provides indications that SCNT steps have determining effects on oocyte transcripts and activation preferences of the reconstituted oocytes compared to intact counterparts. In this sense, the optimal condition for activation of zona-free oocyte post reconstitution is 5 μM ionomycin for 5 min followed by 2 h of incubation with 2 mM 6-DMAP.
Footnotes
Acknowledgments
This study was funded by a grant from Royan Institute of IRI. The authors would like to gratefully thank Mrs. Mansouri for statistical analysis of data, and Mr. Khajo for ovary preparation.
Author Disclosure Statement
No competing financial interests exist.
H.S.M. and N.E.M.H. conceived and designed the experiments; K.M., H.M., H.S.M., O.S., F.M., A.M., and N.E.M.H. performed the experiments; and H.S.M. and N.E.M.H. wrote the paper.