
Abstract
Abstract
Abnormal epigenetic reprogramming of the donor nucleus after somatic cell nuclear transfer (SCNT) is thought to be the main cause of low cloning efficiency. Following SCNT, the donor nucleus often fails to express early embryonic genes and establish a normal embryonic pattern of chromatin modification. Therefore, in this study, we have attempted to improve epigenetic reprogramming of the donor nucleus and cloned embryos with Zebularine and Scriptaid. Yak fibroblasts were treated with 20 μM Zebularine alone or 20 μM Zebularine plus 0.5 μM Scriptaid for 24 h, whereas yak cloned embryos were treated exclusively with 0.5 μM Scriptaid for 12 h. There was no effect on cellular viability and proliferation after drug treatment. The treatment of fibroblasts with Zebularine or Zebularine plus Scriptaid increased histone acetylation of histone 3 lysine 9 (H3K9), but decreased the level of DNA methylation of Oct-4 and Sox-2 promoter regions. When donor cells were used after Zebularine plus Scriptaid treatment to reconstruct cloned embryos and then treated with Scriptaid, the developmental competence and cryosurvival of embryos were improved significantly. In addition, the relative expression of Oct-4 and Sox-2 were increased significantly. The expression levels of Dnmt-1 and Hdac-1 were significantly decreased when fibroblasts and cloned embryos were treated with Zebularine or Scriptaid. This work provides functional evidence that treatment with Zebularine and Scriptaid modifies the epigenetic status of yak fibroblasts, subsequently enhancing in vitro developmental potential and the quality of yak cloned embryos.
Introduction
Accumulated evidence has shown that treated donor cells or early reconstructed embryos with epigenetic remodeling drugs can significantly improve preimplantation developmental competence (Akagi et al., 2011; Costa-Borges et al., 2010) and the blastocyst quality of cloned embryos (Cui et al., 2011; Lee et al., 2011). Furthermore, an improvement of full-term development has also been reported in mice and bovine regarding production of cloned animals using 5-aza-2′-deoxycytidine (AZA) and trichostatin A (TSA) (Kishigami et al., 2006; Wang et al., 2011b). However, the molecular mechanisms remain unknown.
The yak is well adapted to a high altitude environment at 3500 meters above sea level in the region of the Qinghai–Tibet Plateau and nearby areas of China. However, its production traits and reproduction rates are low and its population is vulnerable to the deteriorating environment. Due to its importance for genetic diversity and as a model for a highland animal, interspecies nuclear transfer (iSCNT) is considered one of the most effective measures for accelerating yak breeding and preventing their extinction (Xiong et al., 2012). Therefore, we carried out a series of experiments to test whether the treatment of nuclear donor cells or reconstructed embryos with the epigenetic remodeling drugs Zebularine and Scriptaid would affect the preimplantation development and blastocyst quality of yak cloned embryos. Zebularine, a cytidine analog containing a 2-(1H)-pyrimidinone ring, is a novel DNA methyltransferase and a cytidine deaminase inhibitor with less toxicity than AZA (Jonathan et al., 2003). Scriptaid is a novel HDAC inhibitor, which improves cloning efficiency and has lower cellular toxicity compared with TSA (Zhao et al., 2009). Yak donor cells were treated with Zebularine alone or Zebularine plus Scriptaid, whereas early embryos were treated exclusively with Scriptaid. We compared the DNA methylation status of Oct-4 and Sox-2 (promoter regions) and the global histone acetylation patterns [histone 3 lysine 9 acetylation (H3K9ac)] of the donor nucleus and compared the developmental competence of yak iSCNT embryos. We also compared the gene expression patterns of donor nucleus and blastocysts using quantitative real-time polymerase chain reaction (RT-qPCR).
Materials and Methods
All procedures were approved by the Animal Care and Use Committee of Southwest University for Nationalities and performed in accordance with animal welfare and ethics. All chemicals and reagents used were purchased from Sigma-Aldrich Chemical Company (St. Louis, MO, USA) unless otherwise stated. Disposable, sterile plasticware was purchased from Nunclon (Roskilde, Denmark). Fibroblasts and embryos were cultured in the same conditions at 38.5°C in air with 5.5% CO2.
Somatic cell preparation and treatment
Yak ear fibroblasts were obtained from a 6-month-old female by the method described by Xiong et al. (2012). Cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco, USA) supplemented with 10% fetal bovine serum (FBS; Gibco, USA), 1 mM sodium pyruvate, 100 IU/mL penicillin, and 100 mg/mL streptomycin. When cells achieved 80% confluence, they were trypsinized, rinsed, and recultivated randomly into three groups: Treatment with 20 μM Zebularine alone (group A), or with 20 μM Zebularine plus 0.5 μM Scriptaid for 24 h (group B), or without any treatment (control group, group C). In this study, the concentration and duration of treatment were chosen based on previous studies (Billam et al., 2010; Liu et al., 2007) and our preliminary experiments. Those cells were then used as nuclear donors in iSCNT to assess its effect on developmental competence.
Interspecies somatic cell nuclear transfer
The oocytes of bovine were collected and matured in vitro as previously described (Xiong et al., 2012). The nuclear transfer procedure and in vitro embryo culture were performed as described by Wang et al. (2011b) with few modifications. Briefly, the cumulus cells were removed with 0.2% hyaluronidase after bovine oocytes matured in vitro for 22 h. Metaphase II oocytes with the first polar body and evenly granulated ooplasm were selected for enucleation. The first polar body and a small amount of the surrounding cytoplasm were aspirated with a 20-μm (inner diameter) glass pipette. The expelled cytoplasm was stained with 10 μg/mL Hoechst 33342 for 10 min, and the removal of the nuclear material was confirmed under ultraviolet radiation. A single donor cell was placed in the perivitelline space of the enucleated bovine oocyte. The oocyte–cell couplet was fused between a pair of platinum electrodes connected to a micromanipulator in microdrops of Zimmermann fusion medium (35 V for 10 μsec). Reconstructed embryos were stored in modified synthetic oviductal fluid (mSOF) containing 5 μg/mL cytochalasin B for 2 h until activation.
Activation and culture
Successfully reconstructed embryos were activated in 5 μM ionomycin for 5 min followed by 4 h of exposure to 2 mM dimethylaminopurine (6-DMAP) in mSOF. During activation, reconstructed embryos were treated with 0.5 μM Scriptaid (iSCNT–Scriptaid group) during the 4-h incubation in 6-DMAP or without (iSCNT group). All embryos were washed twice in mSOF. iSCNT group embryos were immediately cultured in mSOF, whereas iSCNT–Scriptaid group embryos were incubated in mSOF supplemented with 0.5 μM Scriptaid for an extra period of 8 h. Afterward, iSCNT–Scriptaid group embryos were washed twice in mSOF and cultured in mSOF as an iSCNT group. Both the donor cells and the reconstructed embryos have not been treated with Zebularine or Scriptaid, this group as control group. Cleavage and blastocyst rates of both groups were evaluated at 24 h and 168 h postactivation, respectively.
Assessment of cell viability/cytotoxicity
For the determination of cell viability and cytotoxicity, the 3-(4,5-dimethylthiazol-2-yl) 2,5-diphenyl-tetrazolium bromide (MTT) assay was conducted for treatment groups as previously described (Mosmann, 1983) with minor modifications. Briefly, fibroblasts in six-well plates were incubated with 3 mg/mL MTT at 38.5°C for 2 h; dimethylsulfoxide (DMSO) was added (1:1, by volume) to each well and the reactants were mixed by pipetting until the Formosan (Sigma; no. 88417) was completely dissolved (Selokar et al., 2012). Each plate was read using a Microplate reader (Gene Company Limited, Hongkong, China) at 570-nm wavelength. All analyses were performed in at least three independent replicate cultures. The absorbance ratio of treated to control cells was calculated and presented as relative cell viability.
Immunodetection of H3K9ac
Donor cells were immunostained with antibodies against H3K9ac (Abcam, Cambridge, UK) as previously described (Wang et al., 2011a). Briefly, cells were washed in phosphate-buffered saline (PBS), fixed with 4% paraformaldehyde in PBS for 30 min, and permeabilized with 0.2% Triton X-100 in PBS for 20 min. Cells were blocked in 2% bovine serum albumin (BSA) in PBS for 15 min and incubated with the primary antibodies (rabbit polyclonal to histone H3 acetyl K9, 1:500) for 1 h. Then cells were washed three times for 5 min each in PBS and incubated for 2 h in the presence of 1:200 diluted fluorescein isothiocyanate (FITC)-labeled secondary antibodies (goat anti-rabbit IgG, Beyotime, China). DNA was stained with 10 μg/mL propidium iodide (PI) solution. As a negative control, immunostaining was performed without primary antibodies.
Bisulfite sequencing (BSP) analysis
The DNAs of each group fibroblasts were treated with the EZ DNA Methylation-Direct™ Kit (Zymo Research, CA, USA) according to the instruction manual. Bisulfite-modified DNAs were amplified with primers designed according to the online MethPrimer software (http://www.urogene.org/methprimer/); the details are listed in Table 1. The PCR products were then cloned into a pMD19-T vector (TaKaRa, DaLian China), followed by verification using colony PCR. Finally, at least nine colonies for each sample were sequenced. Each experiment was repeated three times. Bisulfite sequencing data and C-T conversion rates were analyzed using the BIQ Analyzer software. Methylation data from bisulfite sequencing were analyzed by computing the percentage of methylated CpGs to the total number of CpGs.
BSP, bisulfite sequencing; RT, reverse transcriptase; F, forward; R, reverse; Tm, annealing temperature.
Counting of cell numbers
Ten blastocysts from each group were randomly selected and stained with 10 μg/mL Hoechst 33342 for 15 min after permeabilization in 4% paraformaldehyde in PBS for 30 min at room temperature and then mounted on a slide in 5 μL of glycerol. The total number of cells was counted under fluorescence using a Nikon eclipse Ti-s microscope (Nikon, Japan) with a digital camera.
Embryo vitrification and warming
The blastocysts of yak iSCNT were vitrified and warmed with a vitrification system (Vitrolife, Sweden) using the Freeze-Kit 1™ and Thaw-Kit 1™ according to the instruction manual. Briefly, embryos were placed for freezing into Cryo-PBS for 5 min, and then gently transferred into EFS 1 for 10 min and EFS 2 for 5 min, respectively. Finally, every two embryos were loaded into ministraws and placed into liquid nitrogen for at least 2 weeks. For thawing, the straws were removed from liquid nitrogen and kept in air for 30 sec. The embryos were gently expelled into EST 1 for 5 min, and then transferred to EST 2 for 5 min and EST 3 for 7 min. Finally, embryos were placed in Cryo-PBS for 6 min and moved into culture medium. Survival of vitrified blastocysts was determined according to re-expansion rates after culture in mSOF for 12 h.
Detection of transcripts by RT-qPCR
The expression of pluripotent transcripts (Oct-4 and Sox-2), DNA methyltransferase (Dnmt-1), and histone deacetylase (Hdac-1) genes was detected by RT-qPCR performed as described previously (Xiong et al., 2012). Donor cells were collected and processed for RNA extraction using TRIzol (Invitrogen, Carlsbad, CA, USA) in accordance with the manufacturer's instructions. Three blastocysts from the same batch of iSCNT were randomly selected for total mRNA extraction with the Cells-to-Signal™ Kit (Ambion Co., Austin, TX, USA) according to manufacturer's instructions. Synthesis of cDNA was performed using the cDNA synthesis kit (TaKaRa, Dalian, China) according to the manufacturer's instructions. The RT-qPCR was performed using the CFX96 detection system (Bio-Rad Laboratories Inc., Hercules, CA, USA) with SYBR Premix Ex Taq™ II (TaKaRa). Primers for all genes were designed as cross-introns by Primer 5.0 software (Premier Biosoft International, Palo Alto, CA, USA) and were based on bovine and yak RNA sequences in the GenBank National Center for Biotechnology Information (NCBI) database (Table 1). Glyceraldehyde-3-phosphate dehydrogenase (Gapdh) was initially used as a housekeeping reference gene as in a previous study (Smith et al., 2007). Reaction mixtures (20 μL) contained cDNA, 10 μL SYBR Green premix, 0.8 μL of the gene-specific primers, and nuclease-free water. Negative controls were cDNA-free reaction mixtures and controls from cDNA synthesis. Thermal cycling conditions were 95°C for 5 min, followed by 40 cycles of 95°C for 10 s, and Tm for 60 s. Each sample was measured in triplicate. Melting curve analysis was performed to check for primer specificity. Amplification efficiency for each cDNA and growth condition was determined as described (Bustin et al., 2009). The relative quantification method (2−ΔΔCt) was used to quantify the gene expression levels of each target gene relative to Gapdh for each sample to determine the relative amount of target mRNA.
Statistical analysis
The experiment was repeated at least three times for each treatment group. Total fluorescence intensity was measured with Image-Pro Plus 6.0 software (Media Cybernetics, Silver Spring, MD, USA) after background subtraction. Data were analyzed with one-way analysis of variance (ANOVA) and Tukey's least significant difference (LSD) test using SPSS 13.0 software (SPSS Inc., IL, USA), and differences were considered significant at p<0.05.
Results
Effects of Zebularine and Scriptaid on the cell proliferation and viability
The cell proliferation and viability of Zebularine or Zebularine plus Scriptaid treatments were evaluated by the MTT assay. As shown in Figure 1, there was no significant difference between treated groups and the control group, indicating that 20 μM Zebularine and 0.5 μM Scriptaid have little cytotoxicity on yak fibroblasts.
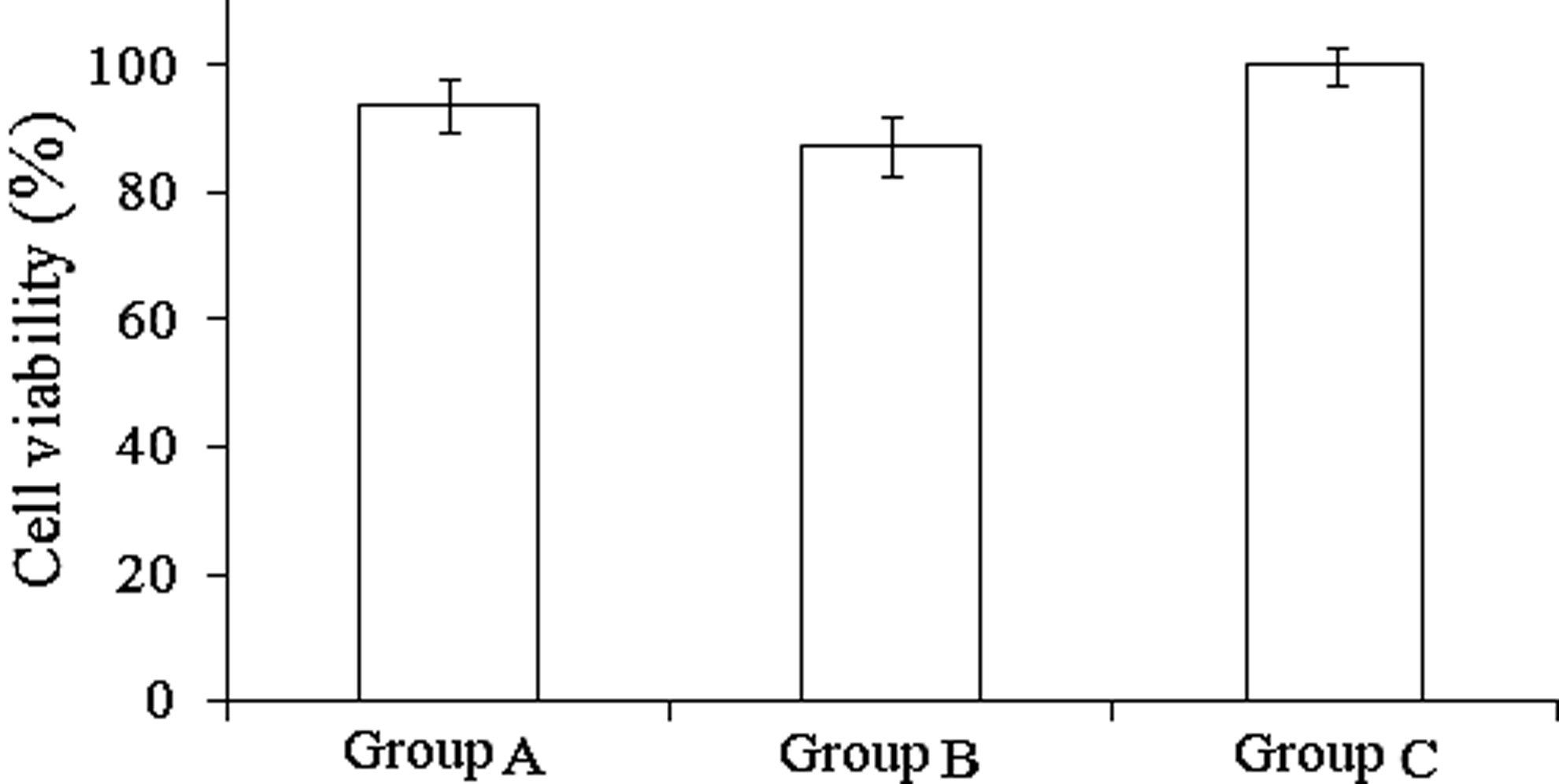
Cells were treated with Zebularine and Scriptaid for 24 h, after which cell viability was determined by MTT assay. (
Effects of Zebularine and Scriptaid on H3K9ac and DNA methylation
To change the global histone acetylation and DNA methylation patterns, fibroblasts were treated with Zebularine alone or Zebularine plus Scriptaid. As determined by immunocytochemistry, both drugs were effective in increasing H3K9ac (Fig. 2). Compared to control cells, fibroblasts treated with Zebularine alone showed an increase in H3K9 acetylation of about 1.4-fold, whereas fibroblasts treated with Zebularine plus Scriptaid showed an increase of about 1.9-fold. As for the results of BSP (Fig. 3), there was significant demethylation of pluripotency gene promoter regions after treating the yak fibroblasts with Zebularine alone or Zebularine plus Scriptaid. In addition, there was significant difference between the Zebularine group (group A) and Zebularine plus Scriptaid group (group B).

Effects of Zebularine and Scriptaid on H3K9ac and relative fluorescence intensity of H3K9ac in yak fibroblasts. H3K9ac in Zebularine alone or Zebularine- plus Scriptaid-treated cells were detected by immunofluorscence analysis (upper) and presented as mean±standard deviation (SD) (lower). Means without a common superscript lowercase letter differed (p<0.05). All images were taken in the same magnification (200×).

DNA methylation analysis of pluripotency genes by bisulfite sequencing of Oct-4 and Sox-2 promoter regions in yak fibroblasts after Zebularine and Scriptaid treatment. Schematics indicate the position of analyzed CpG islands in promoter regions. (Black circles) Methylated CpGs; (white circles) unmethylated CpGs. Within a group, means without a common lowercase letter differed (p<0.05).
Effects of Zebularine and Scriptaid on in vitro development and quality of cloned embryos
Because the treatment of fibroblasts with Zebularine plus Scriptaid resulted in a significant increase of H3K9 acetylation, we decided to evaluate the use of these cells as nuclear donors for iSCNT. Developmental rates to the cleavage and blastocyst stages of the embryos were analyzed (Table 2). Treatment of both donor cells and early embryos with epigenetic modification drugs had no effect on the cleavage rates, but the ability of cloned embryos to develop into blastocysts was significantly improved after the treatment. Moreover, the total cell number of cloned embryos significantly increased after treatment with epigenetic modification drugs. In addition, the cryosurvival rate of vitrified–thawed cloned blastocysts derived from early embryos treated with 0.5 μM Scriptaid was significantly higher (p<0.05) than that of control group (Table 3).
Values with different superscripts (a–c) within a column are significantly different (p<0.05).
iSCNT, interspecies somatic cell nuclear transfer; SEM, standard error of the mean.
Values with different superscripts (a, b) within a column are significantly different (p<0.05).
iSCNT, interspecies somatic cell nuclear transfer.
Effects of Zebularine and Scriptaid on mRNA expression
The relative abundance of the gene transcripts was studied (Fig. 4). Compared with the control group, donor cells treated with Zebularine alone or Zebularine plus Scriptaid had significantly increased expression levels of Oct-4 and Sox-2 (p<0.05), along with significantly decreased the expression level of Dnmt-1 (Fig. 4A). Transcription levels of Hdac-1 were decreased significantly after treatment with a combination of Zebularine and Scriptaid (p<0.05), although there were no significant differences between the control group and Zebularine treatment group. Using yak fibroblasts after pretreatment with Zebularine plus Scriptaid as donor nucleus resulted in reconstructed yak iSCNT embryos. Early cloned embryos treated with Scriptaid for 12 h resulted in significantly decreased expression of Hdac-1, and significantly increased expression of Oct-4 than the control group (Fig. 4B).
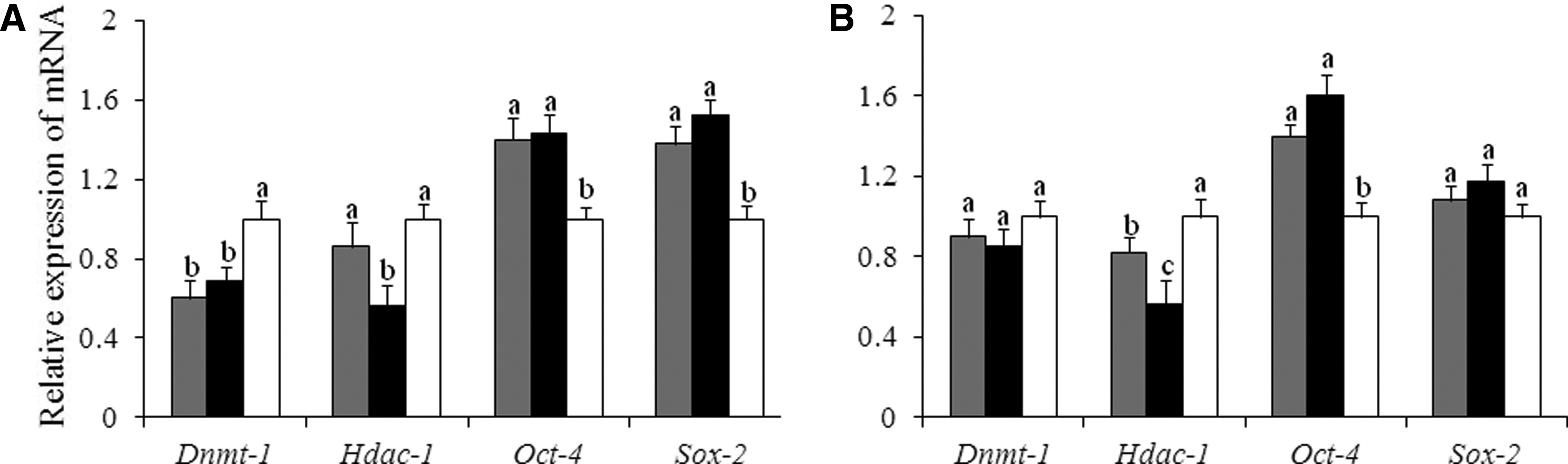
Effects of epigenetic modification drugs on the relative abundance of transcript levels in donor cells and cloned embryos. (
Discussion
Although a large number of species have been cloned successfully using SCNT technology, cloning efficiency is still extremely low (Gao and Latham, 2004). Related studies have provided accumulated evidence that the low efficiency of SCNT was due to incomplete nuclear reprogramming (Chung et al., 2009; Lagutina et al., 2010). Therefore, there have been attempts to improve epigenetic reprogramming of donor cells and cloned embryos, including treatment of donor cells and early embryos with epigenetic modification drugs (e.g., TSA and AZA) to enhance the developmental potential of cloned embryos by altering epigenetic marks in the nucleus. Due to the high toxicity of AZA and TSA, an alternative strategy has been to find less toxic drugs to treat somatic cells. Both Zebularine and Scriptaid are commonly used in therapeutic treatments in humans, but with epigenetic side effects (Chen et al., 2011). Zebularine is a novel DNA methyltransferase inhibitor, which was developed as a more stable and less toxic drug (Yoo et al., 2004), preventing the enzyme from methylating position 5 of cytosines clustered in regulatory CpG islands. Previous studies showed the ability of Zebularine to sustain the demethylation state of the 5′ region of tumor suppressor genes and other methylated genes in T24 and HT-29 cells (Cheng et al., 2004). Scriptaid is a novel HDAC inhibitor, which has a relatively higher histone acetylation activity and lower cellular toxicity compared with TSA (Zhao et al., 2009). It has been reported that Scriptaid treatment could significantly increase the in vitro and in vivo development of cloned embryos (Van Thuan et al., 2009). Because there are no data regarding the use of Zebularine in SCNT, the concentration and treatment time used in the present study were from studies regarding its effect on tumoral cells (Billam et al., 2010; Liu et al., 2007). Scriptaid concentration was based on previous works related to its use in SCNT in bovine and pig (Wang et al., 2011a; Whitworth et al., 2011). On the basis of the MTT assay, there was minimum toxicity of Zebularine and Scriptaid on the proliferation and viability of yak fibroblasts.
The epigenetic status of the donor nucleus has an important effect on the developmental potential of cloned embryos produced by SCNT. The epigenetic state of the donor nucleus must be erased and an embryonic epigenetic state pattern should be established in cloned embryos (Wang et al., 2011b). There is growing evidence suggesting that the developmental defects in cloned embryos correlated with the epigenetic modifications to the chromatin of donor cells and proper handing with epigenetic modification drugs can improve the epigenetic patterns (Kishigami et al., 2006). Lee et al. (2010) reported that treating cat cells with TSA before SCNT significantly increased the H3K9ac level of the donor nucleus. Gómez et al. (2011) showed that pretreated donor cells with TSA improved the H3K9ac and H3K9me2 level; Sangalli et al. (2012) treated bovine fibroblasts with AZA and TS, which increased histone acetylation. Likewise, we observed that acetylation in H3K9 of yak fibroblasts was significantly increased when they were treated with Zebularine alone or Zebularine plus Scriptaid for 24 h. However, Shi et al. (2003) found that sodium butyrate (NaBu) has no obvious effect on the histone acetylation levels of donor cells. Travers et al. (2002) suggested that the histone acetylation levels of human CD34+ hematopoietic progenitor cells are not significantly changed after TSA treatment. Those inconsistent results may due to the different sensitivities of the distinct cells to drugs; also, the treatment programs in those studies are inconsistent. Moreover, the mechanisms of different drugs on modifying the epigenetic reprogramming of donor cells are also inconsistent and require further investigation.
DNA methylation is another critical component of the epigenetic status of donor nucleus. The methylation status of the donor cell nucleus can strongly influence the efficiency of nuclear transfer–derived blastocysts. That is possible because the DNA methylation status of donor nucleus affects the gene expression in early cloned embryo. However, the pattern of DNA methylation is abnormal in cloned embryos (Bonk et al., 2008), and abnormal DNA hypermethylation is believed to be associated with the low success rate of SCNT (Simonsson and Gurdon, 2004). Therefore, previous attempts to improve SCNT efficiency were by changing the DNA methylation level of the donor nucleus using demethylating drugs (Enright et al., 2005). A positive effect was observed, but those drugs also had strong toxicity (Enright et al., 2005). In the present study, the donor cells were treated with Zebularine and Scriptaid with minor toxicity. The methylation status of Oct-4 and Sox-2 promoter regions was detected, and we found that the methylation of Oct-4 and Sox-2 promoter regions was decreased significantly and the cloning efficiency was enhanced predominantly when donor cells treated with Zebularine plus Scriptaid.
Several studies have been performed to improve cloning efficiency by treating donor cells and embryos with epigenetic modification drugs. Kishigami et al. (2006) found that treating early cloned embryos with TSA significantly improved the in vitro developmental competence and full-term development of cloned mice embryos. Zhao et al. (2009) reported that Scriptaid treatment significantly improved the in vitro and in vivo development of cloned pig embryos. Our previous study showed that treatment of both donor cells and early cloned embryos with AZA and TSA significantly improved the ability to develop into blastocysts and the birth rate of cloned bovine (Wang et al., 2011b). However, it has also been reported that there was no positive effect on in vitro and in vivo developmental competence of cloning embryos in rabbits and pigs (Meng et al., 2009; Martinez-Diaz et al., 2010). This inconsistency may be due to different species or the use of different drugs and the different treatment concentrations and times.
In the present study, we chose yak fibroblasts after treatment with Zebularine plus Scriptaid as donor nuclei for iSCNT, and treated early cloning embryos with Scriptaid alone. The blastocyst formation rates were significantly higher than the controls (group iSCNT). In addition, donor cells and early cloned embryos treated with epigenetic modification drugs significantly increased the capability of cryosurvival of cloned blastocysts than in the untreated group. Perhaps the increased acetylation and decreased methylation levels of the donor nucleus facilitate nucleus reprogramming and subsequently enhance the developmental competence of cloned embryos.
Furthermore, we tested whether the patterns of DNA methylation and histone acetylation affect gene expression. Expression of Oct-4 and Sox-2 is essential for proliferation of fibroblasts and early embryonic development. Both were expressed appropriately after nuclear transfer and correlated tightly with the cloning efficiency (Simonsson and Gurdon, 2004). Dnmt-1 and Hdac-1 are required to maintain the DNA methylation and histone acetylation patterns of the donor nucleus and cloned embryos; absence or overexpression of Dnmt-1 and Hdac-1 will cause abnormal embryonic development (Kurihara et al., 2008). The relative expression levels of Oct-4 and Sox-2 were improved after Zebularine and Scriptaid treatment; however, Dnmt-1 and Hdac-1 expression had a downward trend. Therefore, we presumed that there was close relation between gene expression and epigenetic modification, and the improvement of cloned embryonic development might be caused by the correction of gene expression after drug treatment.
Conclusion
Our work provides functional evidence that treatment of both nuclear donor cells and yak cloned early embryos with the epigenetic modification drugs used in the present study has a significant effect on the epigenetic patterns of donor cells, the preimplantation development of cloned embryos, and mRNA expression patterns. However, further studies should be performed to ascertain the effects of Zebularine and Scriptaid on full-term development of yak iSCNT embryos.
Footnotes
Acknowledgments
The present study was supported by the National Science and Technology Program of China (No.2012BAD13B06) and the Fundamental Research Funds for the Central Universities of Southwest University for Nationalities (11NFW03).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.