
Abstract
Abstract
Abnormal epigenetic reprogramming, such as histone acetylation, might cause low efficiency of interspecies somatic cell nuclear transfer (iSCNT). This study was conducted to evaluate the effects of trichostatin A (TSA) on the developmental competence and histone acetylation of iSCNT embryos reconstructed from cat somatic cells and bovine cytoplasm. The iSCNT cat and parthenogenetic bovine embryos were treated with various concentrations of TSA (0, 25, 50, or 100 nM) for 24 h, respectively, following fusion and activation. Treatment with 50 nM TSA produced significantly higher rates of cleavage and blastocyst formation (84.3% and 4.6%, respectively) of iSCNT embryos than the rates of non-TSA–treated iSCNT embryos (63.8% and 0%, respectively). Similarly, the treatment of 50 nM TSA increased the blastocyst formation rate of parthenogenetic bovine embryos. The acetylation levels of histone H3 lysine 9 (H3K9) in the iSCNT embryos with the treatment of 50 nM TSA were similar to those of in vitro–fertilized embryos and significantly higher (p<0.05) than those of non-TSA–treated iSCNT embryos (control), irrespective of the embryonic development stage (two-cell, four-cell, and eight-cell stages). These results indicated that the treatment of 50 nM TSA postfusion was beneficial for development to the blastocyst stage of iSCNT cat embryos and correlated with the increasing levels of acetylation at H3K9.
Introduction
Bovine cytoplasm has shown capabilities for supporting in vitro development of iSCNT embryos reconstructed with somatic cells from various unrelated mammalian species such as sheep, pig, monkey, dog, and yak (Dominko et al., 1999; Murakami et al., 2005). Very few reports in the literature describe the capability of bovine oocytes to reprogram the nucleus of felid species. Thongphakdee et al. (2008) reported that no iSCNT cat embryo was able to develop beyond the eight-cell stage. That developmental block of iSCNT cat embryos might be associated with a developmental cell block and mitochondrial incompatibility between the recipient oocytes and donor cells (Thongphakdee et al., 2008).
Incomplete donor nuclei reprogramming and abnormal epigenetic reprogramming (DNA methylation or histone modification) are thought to be related to low efficiency in SCNT-cloned and iSCNT-cloned embryos (Arat et al., 2003; Chen et al., 2006; Lee et al., 2010). Histone acetylation provides the greatest potential for unfolding chromatin to recruit different transcriptional factors. Removal of acetylated groups by histone deacetylases (HDACs) is generally associated with gene silencing (Shi et al., 2008). The relationship between abnormal patterns of histone acetylation and the developmental failure in cloned embryos has been suggested (Shi et al., 2008). Previous reports in the literature have described that in vitro embryo development and full-term development of intraspecies cloned embryos have been improved by epigenetic modification of donor cells or early cloned embryos with trichostatin A (TSA), an HDAC inhibitor that increases histone acetylation, e.g., in pigs (Li et al., 2008; Zhang et al., 2007), mice (Kishigami et al., 2006; Maalouf et al., 2009), and cattle (Sawai et al., 2012). Moreover, the histone acetylation patterns of SCNT embryos treated with TSA reportedly resemble those of naturally fertilized embryos (Shi et al., 2008; Wang et al., 2007). TSA-treated SCNT mouse embryos develop to term because TSA improves nuclear remodeling in one-cell embryos (Maalouf et al., 2009). Therefore, it might be possible to improve the in vitro development of iSCNT embryos reconstructed from cat somatic cell and bovine cytoplast through modification of the histone acetylation level with the treatment of TSA.
This study was conducted to ascertain the effects of TSA at different concentrations on the in vitro developmental competence of iSCNT cat embryos and to investigate the relative intensity levels of acetylation of histone H3 lysine 9 (H3K9ac) in TSA-treated iSCNT cat embryos.
Materials and Methods
Preparation of recipient oocytes and domestic cat somatic cells for nuclear transfer
Bovine oocytes were matured according to procedures described by Taniguchi et al. (2007) with minor modifications. Cumulus–oocyte complexes (COCs) were cultured in tissue culture medium-199 (TCM-199; Invitrogen, Carlsbad, CA, USA) supplemented with 2.5 μg/mL of taurine (Sigma-Aldrich, St. Louis, MO, USA), 0.02 IU/mL of follicle-stimulating hormone (FSH; Kawasaki Mitaka Seiyaku K.K., Kawasaki, Japan), 5% fetal bovine serum (FBS; Invitrogen), 20 μg/mL of epidermal growth hormone (EGF; Sigma-Aldrich), and 50 μg/mL of gentamicin (Sigma-Aldrich) for 22 h at 38.5°C in a humidified atmosphere containing 5% CO2.
Domestic cat fibroblast cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen) supplemented with 20% (vol/vol) FBS and 50 μg/mL gentamicin at 37°C in a humidified atmosphere containing 5% CO2. Once the fibroblast cells reached complete confluence, cells were trypsinized with 0.25% (wt/vol) trypsin (Invitrogen). They were either frozen for storage or used as donors for nuclear transfer (Kaedei et al., 2010).
SCNT, activation, in vitro culture of embryos, and TSA treatment
SCNT was conducted according to the methods previously described by Taniguchi et al. (2007). Briefly, the zona pellucida above the first polar body was cut with a glass needle and a small volume of cytoplasm was then squeezed out (the metaphase spindle and first polar body were visualized after incubating oocytes in 3 μg/mL of Hoechst 33342; Sigma-Aldrich). A single cat cell was then placed into the perivitelline space of the enucleated oocyte. Couplets were fused and activated simultaneously with a single DC pulse of 2.3 kV/cm for 30 μs delivered by two electrode needles (LF101, Nepa Gene Co. Ltd., Chiba, Japan) connected with a micromanipulator (MO-202D, Narishige Co. Ltd., Tokyo, Japan). To ascertain the effects of different concentrations of TSA on in vitro developmental competence of iSCNT cat embryos, the fused couplets were cultured for 5 h in a modified synthetic oviductal fluid (mSOF) (Kwun et al., 2003) supplemented with 10 μg/mL of cycloheximide (Sigma-Aldrich) and TSA (Wako Pure Chemical Industries Ltd., Tokyo, Japan) with different concentrations (0, 25, 50, and 100 nM). The concentrations of TSA examined in this experiment referred to the previous studies (Akagi et al., 2011; Gómez et al., 2011; Sawai et al., 2012), which demonstrated positive effects of TSA treatment on the acetylation levels or the development of bovine and cat SCNT embryos. The fused couplets were then transferred to mSOF containing TSA at the same concentration and cultured for an additional 19 h. The iSCNT cat embryos were cultured according to the method reported by Kaedei et al. (2010), who demonstrated that 81% of cat–cow iSCNT embryos cleaved by culture at 38.5°C after fusion. After 24 h of TSA treatment, embryos were cultured in mSOF supplemented with 4 mg/mL bovine serum albumin (BSA; Sigma-Aldrich) for 2 days and further co-cultured with bovine cumulus cells in mSOF supplemented with 5% FBS at 38.5°C in a humidified atmosphere of 5% CO2 for an additional 5 days to evaluate their ability for blastocyst formation. At the end of culture, all embryos were stained with Hoechst 33342 for counting the total cell number (Fig. 1) according to procedures described by Kaedei et al. (2010).
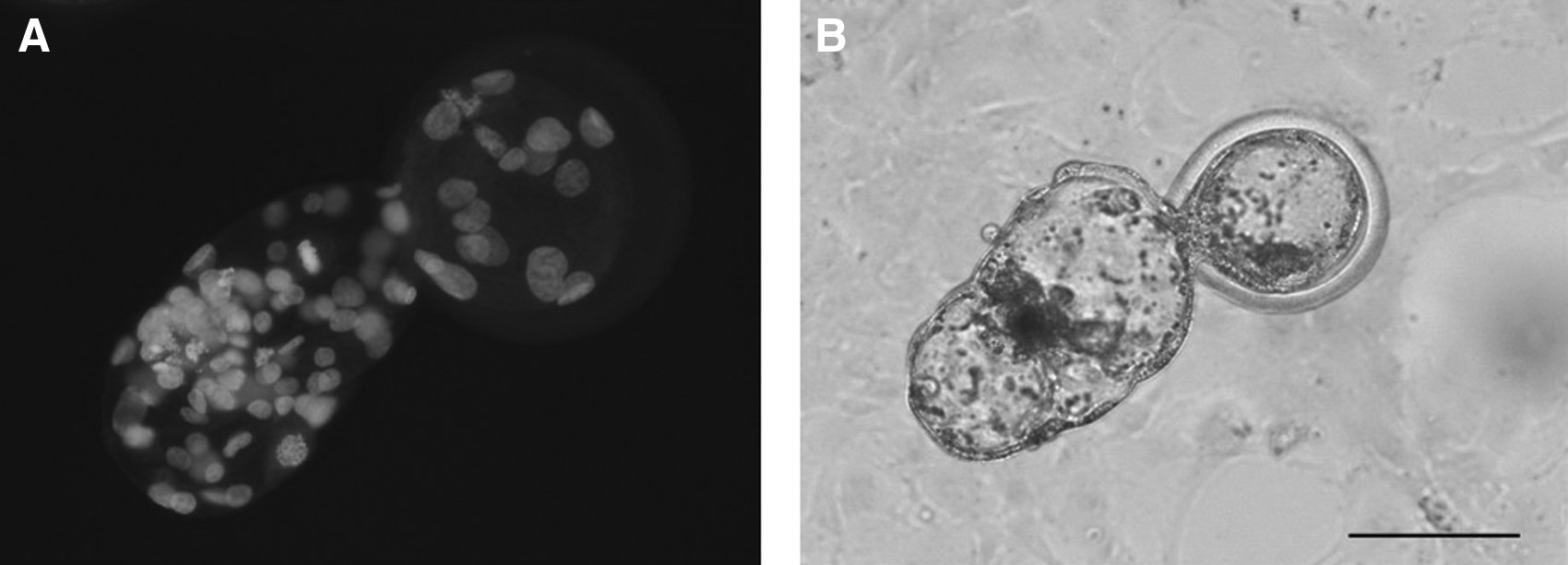
Images of Hoechst 33342 staining (
Parthenogenetic embryos served as embryo developmental controls. In vitro–matured bovine oocytes were activated by a single DC pulse of 2.3 kV/cm for 30 μs using electrode needles with the same methods as those described for iSCNT embryos. The oocytes were then cultured in mSOF containing 10 μg/mL cycloheximide, 5 μg/mL cytochalasin B, and different concentrations (0, 25, 50, or 100 nM) of TSA for 5 h. The activated oocytes were transferred to mSOF medium with TSA at the same concentration and cultured for an additional 19 h. After 24 h of TSA treatment, the oocytes were cultured and monitored as noted for iSCNT embryos.
Fluorescent immunodetection of acetylation on H3K9 in iSCNT cat embryos
Fluorescent immunodetection of H3K9ac in iSCNT cat embryos was performed in the TSA and non-TSA (control) treatment groups. The concentration of 50 nM TSA, the most suitable one for the development of iSCNT cat embryos (Table 1), was used for TSA treatment after fusion. The two-cell stage embryos were collected 24 h postfusion (end point of TSA treatment), whereas the four-cell and eight-cell stage embryos were collected at day 3 of the culture. To compare with naturally fertilized embryos, in vitro-fertilized (IVF) bovine embryos were used. IVF was carried out according to the method described by Taniguchi et al. (2007). The two-cell stage embryos were collected at 24 h postinsemination (PI), and the four- and eight-cell stage embryos were collected at 48 h PI for fluorescent immunodetection of AcH3K9.
Data are expressed as mean±standard error of the mean (SEM). Four replicated trials were conducted.
Couplets were reconstructed from cat somatic cell and bovine cytoplasm and treated with TSA with various concentrations for 24 h after fusion.
Mean values in the same columns with different superscripts are significantly different (p<0.05).
TSA, trichostatin A; iSCNT, interspecies somatic cell nuclear transfer.
All of the following steps were carried out at room temperature (RT) and all solutions were prepared in 10% FBS/phosphate-buffered saline (PBS), unless otherwise stated. Embryos were fixed in 3.7% paraformaldehyde overnight at 4°C, permeabilized with 0.1% TritonX-100 (Sigma-Aldrich)/PBS for 40 min, and stored in 1% (wt/vol) BSA/PBS overnight at 4°C. Permeabilized embryos were incubated with 10% goat serum (Nichirei, Tokyo, Japan)/PBS for 1 h to block nonspecific binding before incubated with primary antibody (5 μg/mL of rabbit polyclonal acetyl-histone H3K9 antibody or 5 μg/mL of rabbit polyclonal histone H3 antibody (Cell Signaling Technology Inc., Danvers, MA, USA) in a moisture chamber overnight. Normal rabbit immunoglobulin G (IgG; Dako, Kyoto, Japan) was used as the negative control. Embryos were subsequently incubated in 4 μg/mL of Alexa 594–conjugated goat anti-rabbit IgG secondary antibody (Invitrogen) for 1 h in a moisture chamber before counterstained with 5 μg/mLl of 4′,6-diamidino-2-phenylindole (DAPI; Invitrogen). Images were obtained with a fluorescence microscope (Nikon Eclipse 80i; Nikon, Tokyo, Japan) equipped with a Nikon DS-Ri1 digital camera (Nikon). Then images in jpeg format were acquired using the NIS-Element D 3.1 (Nikon) imaging software package running on a workstation (Dell Optiplex 960 PC; Dell Inc., Austin, TX, USA).
Semiquantification of fluorescence intensities in the embryos
The fluorescence images of each nucleus within an embryo were taken under the following conditions: Alexa Fluor 594 dye, DAPI, and in bright fields. The signal intensities of fluorescence from H3K9ac, histone H3, and DAPI-nucleic acid staining were measured automatically using imaging software under the area of nucleus by manually outlining a limited area of each nucleus within an embryo, except overlapping or folded nuclei. Fluorescence intensities of embryonic cytoplasm and background were quantified using the same method. The mean intensity in each examined nucleus was recorded. The relative intensity levels of H3K9ac in each nucleus were calculated using the following formula.
Relative intensity levels of histone H3 in each nucleus were also calculated using the same formula. Subsequently, average values of relative intensity levels of H3K9ac and histone H3 in each embryo were calculated. These average values of each embryo were used for additional calculations to ascertain the average value of relative intensity levels of H3K9ac and histone H3 in each treatment. The data compensation in each treatment during the experiments was performed using the average value of relative intensity levels of H3K9ac and histone H3 in control samples without TSA treatment.
Statistical analysis
Data related to the developmental rates of embryos were expressed as mean±standard error of the mean (SEM). Percentage data of embryonic development and intensity levels of H3K9ac and histone H3 in embryos were subjected to arc-sin transformation before analysis of variance (ANOVA). Transformed data were tested by the Kruskal–Wallis test, followed by Fisher's protected least significant difference (PLSD) post hoc test. Differences with a probability value (p) of 0.05 or less were considered statistically significant.
Results
In vitro development of iSCNT cat embryos and bovine parthenogenetic embryos
The effects of TSA concentration on the in vitro development of iSCNT cat embryos and bovine parthenogenetic embryos are shown, respectively, in Tables 1 and 2. Treatment of iSCNT cat couplets with 50 nM TSA significantly increased the rates of total cleavage and embryos developed to the morula stage compared with the couplets without TSA treatment (p<0.01) (Table 1). When the iSCNT cat couplets were treated with 25 nM and 50 nM TSA, 0.7% and 4.6% of fused couplets developed to the blastocyst stage, respectively. In contrast, no couplet without TSA treatment developed beyond the 16-cell stage. An increase of TSA concentration to 100 nM did not enhance embryo development to the blastocyst stage.
Data are expressed as mean±standard error of the mean (SEM). Four replicated trials were conducted.
The parthenogenetic embryos were treated with TSA at various concentrations for 24 h.
Mean values in the same columns with different superscripts are significantly different (p<0.05).
TSA, trichostatin A.
Bovine parthenogenetic embryos with the treatment of 50 nM TSA yielded a significantly higher rate of blastocyst formation than embryos without TSA treatment (p<0.05) (Table 2). However, an increase of TSA concentration to 100 nM showed negative effects on the rates of embryo cleavage and blastocyst formation than other TSA treatment groups showed.
Characterization of acetylation of H3K9 in iSCNT cat embryos with or without treatment of 50 nM TSA in comparison to bovine IVF embryos
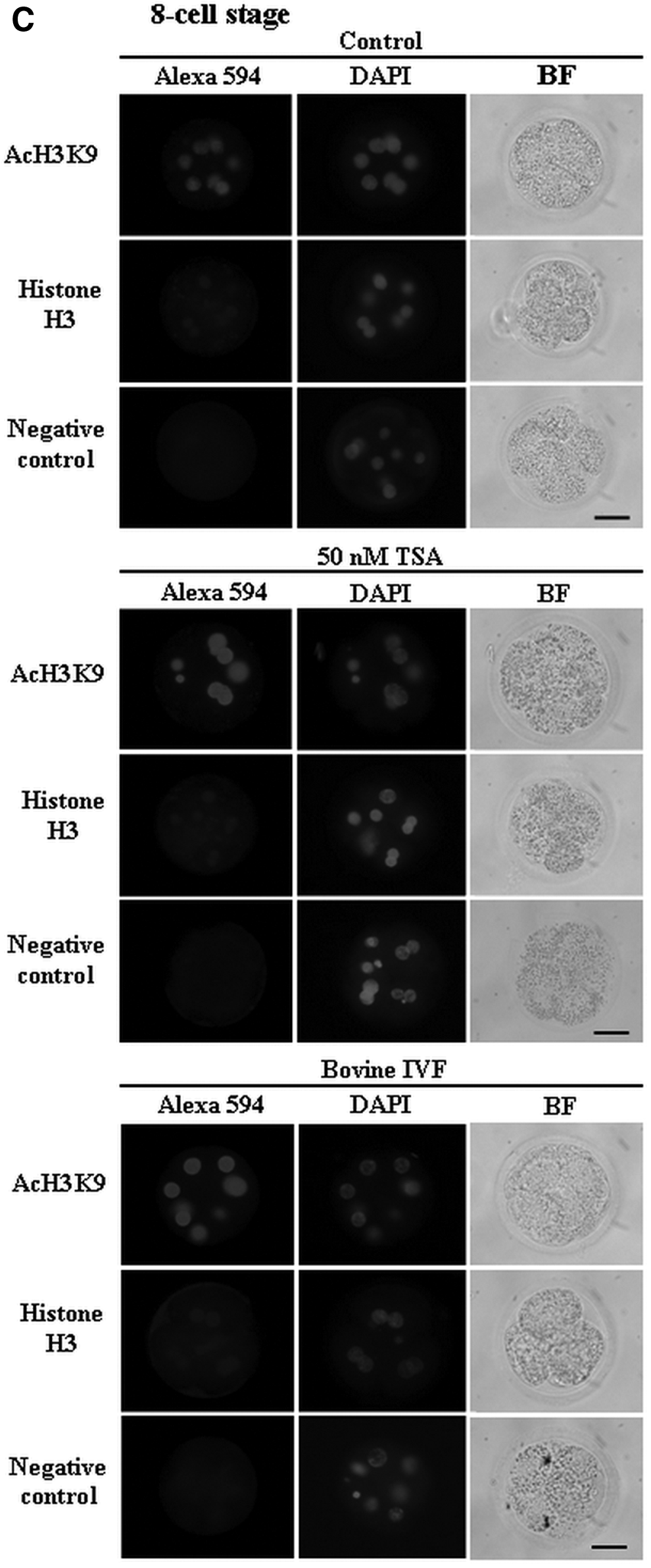
The levels of acetylation of H3K9 in iSCNT cat embryos were evaluated with or without the treatment of 50 nM TSA in comparison to bovine IVF embryos (Figs. 2 and 3). All nuclei of iSCNT cat embryos at the two-cell, four-cell, and eight-cell stages showed positive immunoreactivity in H3K9ac and histone H3, irrespective of the TSA treatment (Fig. 2). The acetylation levels of H3K9 in the nuclei of embryos with TSA treatment were significantly higher (p<0.05) than those of embryos without TSA treatment (control), irrespective of the embryonic development stage (Fig. 3). Nuclear intensities of H3K9ac in TSA-treated embryos at the four-cell and eight-cell stages were similar to those in bovine IVF embryos at the same stage, whereas the intensity of TSA-treated embryos at two-cell stage were significantly higher (p<0.05) than that of two-cell–stage IVF embryos. In contrast, the levels of histone H3 in the embryos were similar in all embryonic stages between the three groups. No significant difference in the background intensity was found between the three groups (p>0.05). The levels of acetylation of H3K9 in iSCNT cat embryos significantly deceased from two-cell to four-cell and eight-cell stages (p<0.05), irrespective of the TSA treatment. Similarly, the acetylation level of H3K9 in bovine IVF embryos at the two-cell stage was higher than that of embryos at the four-cell or eight-cell stage (p<0.05). In contrast, no difference of histone H3 levels in the embryos was found among the embryonic stages, irrespective of the TSA treatment.
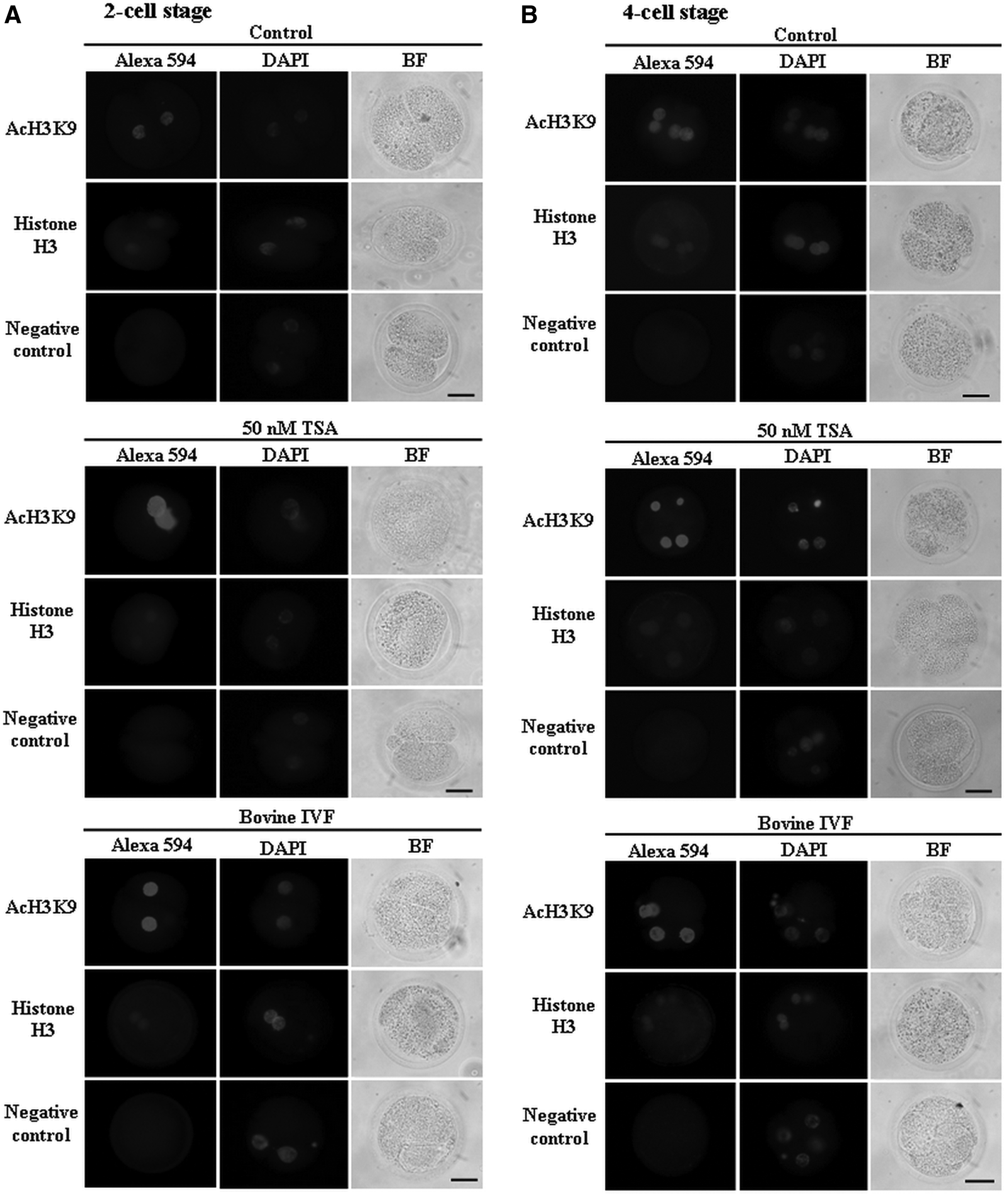
Immunolocalization of acetylation on H3K9 (AcH3K9) and histone H3 in the two-cell (

Relative intensity levels of acetylation on H3K9 (AcH3K9) and histone H3 in the two-cell (
Discussion
The iSCNT technique holds great promise for the conservation of wild or endangered animal species. However, incomplete nuclear reprogramming, low blastocyst rate, and abnormal epigenetic reprogramming remain as major obstacles to the availability of iSCNT (Shi et al., 2008; Wu et al., 2010). Our results show that, in both of the cat iSCNT and bovine parthenogenetic embryos, the treatment of 50 nM TSA for 24 h, respectively, following fusion and activation contributed to significantly higher rates of blastocyst formation compared to the TSA nontreatment group. These results are in agreement with several studies on intraspecies SCNT, e.g., in pigs (Zhao et al., 2010), sheep (Hu et al., 2012), and cattle (Sawai et al., 2012), in which the treatment with 50 nM TSA leads to a significant increase in the blastocyst formation rate of SCNT embryos. Maalouf et al. (2009) demonstrated that treatment with 5 nM TSA not only enhanced the development of mouse SCNT embryos but also increased the numbers of inner cell mass and live offspring. In contrast, no differences between the development rates of bovine SCNT embryos treated with 5 nM and 500 nM of TSA have been reported (Akagi et al., 2011; Sawai et al., 2012). In the present study, we found that an increase of TSA concentration to 100 nM exhibited negative effects on the development of cat iSCNT and bovine parthenogenetic embryos. It has been suggested that treatment of TSA with high concentration or long-term exposure results in developmental defects after implantation (Kishigami et al., 2006; Zhao et al., 2009). Moreover, the effect of TSA depends on treatment conditions and the donor cells (Akagi et al., 2011; Sawai et al., 2012). Therefore, reduction of the development of iSCNT cat embryos may result in part from the exposure of TSA with a high concentration. In contrast to our results, TSA treatment has been shown to have no effects on the embryonic development of iSCNT, e.g., guar–cow (Srirattana et al., 2012), human–rabbit (Shi et al., 2008), and sei whale–cow (Bhuiyan et al., 2010). The difference in results might be associated with the TSA applications (concentration, timing, and the onset of treatment), species-specific effects, and phylogenetic distance between the oocyte and somatic cell donor. The selection of optimized TSA applications for different species might be an important key to improve the success rate in animal SCNT (Wang et al., 2011), especially for iSCNT, for which the genetic distance between the donor cell and recipient cytoplast is important.
In this study, all nuclei of iSCNT cat embryos at the two-cell, four-cell, and eight-cell stages showed positive immunoreactivity in AcH3K9 and histone H3. The levels of histone H3 were not significantly different between the TSA-treated embryos and nontreated (control) embryos in any examined stage. This fact shows that TSA did not affect the levels of histone H3 as it did on the acetylation levels. Significantly higher acetylation levels of H3K9 in iSCNT cat embryos were observed in all embryonic stages of the TSA-treated embryos as compared to those of control embryos. In the TSA treatment group, moreover, a high level of acetylation of H3K9 was observed in the two-cell stage embryos, and decreased at the four-cell to eight-cell stage embryos.
These results suggest that TSA might provide the ability to modify the patterns of deacetylation–reacetylation on histone lysine residue in iSCNT cat embryos. After the chromatin of cat cell was possibly deacetylated by HDACs in bovine oocytes, the reacetylation of histone lysine residue (e.g., H3K9) might have occurred during nuclear formation and development to the two-cell stage of iSCNT cat embryos within a limited time of nuclear reprogramming. It has been reported that deacetylation events occurring during oocyte activation are independent from reactivation of the genes responsible for the ability of donor cells to develop to blastocysts after SCNT (Rybouchkin et al., 2006). Moreover, we observed that the treatment of 50 nM TSA increased the rates of cleavage and blastocyst formation compared to the TSA nontreatment group.
These observations indicate that the effect of TSA is most likely to be associated with the events of re-acetylation in the stages of pronuclear formation to the two-cell stage. In these stages, high acetylation of H3K9 is necessary to establish embryonic epigenetic characteristics and gene expression in cloned embryos (Stein et al., 1997; Worrad et al., 1995). However, the underlying mechanism of TSA to increase the acetylation level of histone lysine residue in the early stage of iSCNT cat embryos has remained unclear. Reportedly, TSA strongly induces acetylation of the genome by blocking the HDAC enzyme (Lee et al., 2010), which changes the chromatin structure, enhances DNA demethylation, and increases the transcriptional activity of the donor cell genome (D'Alessio et al., 2007). Maalouf et al. (2009) also suggested that the induction of histone acetylation by TSA improves opening of the chromatin, sustaining mobility and relocalizing of constitutive heterochromatin as well as other genomic sequences. Consequently, we suppose that TSA treatment improves nuclear remodeling of iSCNT cat embryos via modified histone acetylation, which is important for early embryo development and subsequent stages.
In the present study, we observed that the iSCNT embryos without TSA treatment were unable to develop beyond the 16-cell stage. Embryonic genome activation (EGA) at the early embryonic stage is the most important event for early embryo development (Meirelles et al., 2004). The developmental failure to the blastocyst stage in iSCNT rhesus monkey–cow embryos probably resulted from the downregulation of EGA, in which the impaired nucleologenesis and aberrant nucleolar formation were involved (Song et al., 2009). The EGA occurs at the five-cell to eight-cell stages in domestic cat (Hoffert et al., 1997) and the eight-cell to 16-cell stages in cow (Camous et al., 1986). Therefore, the developmental arrest at eight-cell to 16-cell stages of iSCNT cat embryos without TSA treatment might be related to insufficient reprogramming of donor nuclei and/or epigenetic status before EGA. However, homologous intensity patterns of histone acetylation from the morula to blastocyst stages between IVF and SCNT embryos have been reported (Wu et al., 2010).
In this study, we observed that the acetylation levels on histone H3K9 in TSA-treated four-cell and eight-cell iSCNT cat embryos more closely resemble bovine IVF embryos. In contrast, intensities of H3K9ac in non-TSA treated iSCNT cat embryos at four-cell and eight-cell stages were clearly lower than those of IVF embryos, indicating that an aberrant histone acetylation before embryonic genomic activation induced the developmental arrest at eight-cell to 16-cell stages (Wu et al., 2010). Moreover, the treatment of TSA has been suggested to support a more accurate regulation of developmental genes at the early development (Maalouf et al., 2009). These results indicate that the normal reprogramming of epigenetic markers including histone acetylation before EGA is the key to the success of iSCNT embryonic development.
In conclusion, the treatment of 50 nM TSA for a total of 24 h after fusion of the couplets between bovine recipient cytoplasts and cat donor cells improves their development to the blastocyst stage by modifying the acetylation levels of H3K9 so as to be similar to those of naturally fertilized embryos before embryonic genome activation. To ascertain the effect of TSA treatment on the development of iSCNT cat embryos to develop to term, additional investigation with embryo transfer is required.
Footnotes
Acknowledgments
The authors express their gratitude to Dr. Atthaporn Roongsitthichai for critical reading of this article. We also thank the staff of the Meat Inspection Office of Kitakyushu City, Japan, for supplying bovine ovaries. This study was supported in part by a grant from the Ministry of Education, Culture, Sports, Science and Technology to T.O. (22580320).
Author Disclosure Statement
No competing financial interests exist.