
Abstract
Abstract
Regenerative therapy is a new strategy for the end-stage heart failure; however, the ideal cell source has not yet been established for this therapy. We expected that the amnion might be an ideal cell source for cardiac regenerative therapy and that the differentiation potency of the human amnion mesenchymal cells (hAMCs) could be improved by overexpression of Oct4, a key factor that maintains the undifferentiated state. A plasmid vector was made by insertion of the Oct4 open reading frame (ORF) under control of a cytomegalovirus (CMV) promoter (pCMV-hOct4) and transfected into hAMCs by electroporation. The optimum induction time was investigated by comparing the quantity of stem cell–specific mRNAs, cardiac-specific mRNAs, and cardiac-specific proteins with time. hAMCs already expressed cardiac-specific proteins such as Nkx2.5 and Connexin43. After pCMV-hOct4 transfection, endogenous Oct4 mRNA and other stem cell markers showed a transient increase. With 5-azacytidine treatment, quantities of the cardiac-specific mRNAs, such as GATA4 and myosin light-chain-2v (Mlc-2v), were increased significantly. After Oct4 overexpression, the highest expression of cardiac-specific mRNAs and stem cell makers was seen at almost the same time. Furthermore, more mature myocardial contraction proteins were observed when hAMCs were induced at specific optimal times after gene transfection. In conclusion, hAMCs were activated to an undifferentiated state by overexpression of Oct4, and their cardiac differentiation potency was improved. Thus, the single-time transfection of the Oct4 expression vector may be a useful strategy for effective cell therapy. The use of cryopreserved hAMCs in cell therapy still requires more investigation.
Introduction
Recently, cell therapy has generated much enthusiasm as a novel potential treatment for heart failure and as a substitute for heart transplantation. Although some cell therapy with somatic cells and somatic stem cells is already performed clinically, the effects are not achieved from differentiation into cardiomyocytes themselves (Deb et al., 2003; Jackson et al., 2001; Murry et al., 2004; Reinecke et al., 2002), but instead from paracrine effects of the transplanted cells (Jain et al., 2001; Orlic et al., 2001a; Orlic et al., 2001b). Therefore, in clinical trials, restoration of the cardiac function as a long-term result and the poor effect of this therapy for those who have a small number of functional cardiomyocytes are new issues (Menasche et al., 2008; Meyer et al., 2009). If we can use cells that beat autonomously like cardiomyocytes, then effective cardiac regenerative therapy will be possible.
Human embryonic stem cells (hESCs) have pluripotency, and they can differentiate into autonomously beating cardiomyocytes (Mummery et al., 2003; Xu et al., 2002). However, hESCs present ethical concerns associated with their origin from human embryos, and they also have the risk of forming teratomas and provoking an immune response. In 2006, Takahashi et al. succeeded in making artificial pluripotent cells, called induced pluripotent stem cells (iPSCs), by inducing four factors—Oct4, Sox2, Klf4, and c-Myc—into fibroblasts using a retroviral vector (Takahashi et al., 2007; Takahashi and Yamanaka, 2006). However, the use of a viral vector, teratoma formation, and the use of an oncogene are hurdles for clinical application.
Human amnion mesenchymal cells (hAMCs), derived from the mesoblast, as is the heart, express some cardiac-specific markers with stem cell markers (Zhao et al., 2005). However, no investigation of cell therapy will be possible by cryopreservation of hAMCs, because the amniotic membrane does not allow delivery.
Some improvements have been reported in iPSC technology. For example, Kim et al. reported that three germ layers were able to differentiate from adult neural stem cells by one factor called Oct4 (Kim et al., 2009a). Okita et al. reported the manufacture of mouse iPSCs by gene transfections multiple times with a plasmid vector (Okita et al., 2010).
We examined the expression of the stem cells markers in cryopreserved hAMCs. Oct4 is considered as a main regulator of ESC pluripotency and self-renewal. Therefore, we investigated whether hAMCs can be transformed to iPS-like cells by a single gene transfection with an Oct4 expression plasmid vector and if this can improve their differentiation potency into cardiomyocytes. To create made-to-order treatment by amnion cryopreservation, we used hAMCs that were cryopreserved once.
Materials and Methods
Isolation and culture of hAMCs
The amniotic membrane was peeled mechanically from the chorion of a placenta obtained with informed consent after an uncomplicated Caesarean section. The study and the use of the amnion membrane were approved by the ethics committee of University of Toyama. The hAMCs were isolated as described previously (Zhao et al., 2005). The tissue was minced and treated with 0.25% trypsin (Sigma-Aldrich Japan, Tokyo, Japan) four times. After this treatment, the epithelial cells were almost completely removed (>98%), and mesenchymal cells were isolated from the amniotic tissues. The tissue pieces were placed in Dulbecco's modified Eagle medium (DMEM; Sigma, Irvine, UK) containing 0.75 mg/mL collagenase (Sigma-Aldrich) and 0.075 mg/mL DNase I (Sigma-Aldrich) and were incubated at 37°C for 45 min with shaking at 100 rpm. The dispersed mesenchymal cells were collected by filtration of the mixture through gauze and centrifugation. Isolated hAMCs were seeded onto plastic dishes at 5×104 cells/cm2 in DMEM supplemented with 10% fetal bovine serum (FBS), 2 mM
Flow cytometry analysis
To evaluate cell-surface makers of hAMCs (6th to 7th passage), hAMCs were trypsinized and incubated in DMEM containing 10% FBS for 30 min. These cells were washed twice in phosphate-buffered saline (PBS; Sigma-Aldrich) followed by blocking with 1% Block Ace (Dainippon Seiyaku Co., Osaka, Japan) and were incubated for 30 min at room temperature with fluorescein isothiocyanate (FITC), phycoerythrin (PE), or R-phycoerythrin covalently linked to cyanin 5.1 (PC5)-conjugated monoclonal antibody–specific human markers associated with human stem cell, mesenchymal, and hematopoietic lineages. Antibodies used were: SSEA-4 (clone MC813-70), CD44 (clone G44-26), CD73 (clone AD2) (all from BD Biosciences, San Diego, CA, USA)
Plasmid construction and electroporation
The open reading frame (ORF) of the human Oct4 fragment (1.3 kb) was removed by BamHI/NotI digestion from a pMXs-hOCT4 vector (Addgene plasmid 17217, Addgene Inc, Cambridge, MA, CA, www.addgene.org) (Kitamura et al., 2003; Takahashi et al., 2007) and subcloned into the BamHI/NotI site of a pIRES2-AcGFP1 vector (Clontech, Mountain View, CA, USA, www.clontech.com), instead of the internal ribosome entry site of the Aequorea coerulescens green fluorescent protein (GFP) coding region, under control of the cytomegalovirus (CMV) promoter pCMV-hOct4, 5.3 kb. A positive control to calculate transfection efficiency was created by removing the enhanced (E) GFP fragment from pEGFP-1 (Clontech) (780 bp) and subcloning it into the same position as human Oct4 fragment (pCMV-EGFP, 4.7 kb).
The cryopreserved hAMCs were thawed and seeded onto plastic dishes at the density of 2000 cells/cm2. hAMCs that were 70–80% confluent were harvested twice in 0.1% trypsin/EDTA (Sigma-Aldrich) and PBS. hAMCs (1.0×106) were transfected with 10.0 μg of pCMV-hOCT4 or pCMV-EGFP using a Nucleofector Kit V (Lonza, Cologne, Germany) and program T-20 on the Nucleofector I (Lonza). The cells were seeded in six-well plates at 5.0×105 cells/well. At 24 h after plasmid transfection, the culture medium (DMEM supplemented 10% FBS) was changed, and then was changed two or three times in a week. Experiments were performed done independently twice and termed experiments T1 and T2.
Cardiac differentiation of hAMCs
FBS (20%) containing DMEM was used as a differentiation medium. The hAMCs without plasmid transfection and the pCMV-hOct4 transfected hAMCs at 2, 4, 5, 6, and 8 weeks after transfection were seeded at a density of 1000 cells/cm2 in the differentiation medium. The next day, 10 μM 5-aza-2′-deoxycytidine (5-Aza) (Sigma-Aldrich) was added for 24 h. The differentiation medium was changed two or three times a week for 4 weeks.
RNA isolation and reverse-transcription polymerase chain reaction
Total RNA was extracted from cultured, pCMV-hOCT4 transfected and cardiac differentiated hAMCs using an RNeasy Mini Kit (Qiagen, Tokyo, Japan) according to the manufacturer's instructions. An RNase-free DNase kit (Qiagen) was used during RNA extraction to digest genomic DNA. As a positive control, human heart total RNA was purchased from Clontech. Total RNA (0.5 μg) was reverse transcribed to cDNA using a ReverTra Ace qPCR RT Kit (Toyobo, Osaka, Japan). Regular PCRs were conducted using the following parameters: Denaturation at 95°C for 5 min followed by 40 amplification cycles of denaturation at 95°C for 1 min, annealing at each (annealing temperature) temperature (°C) for 1 min, and extension at 72°C for 1 min, followed by a final extension at 72°C 10 min. GAPDH was used as an internal standard. The PCR products were size fractionated by 2% agarose gel electrophoresis, stained with ethidium bromide, and visualized using ultraviolet illumination. For the detection of Oct4 RNA, we used the following primer sets: the Oct4A primer set was specific for Oct4A, which is localized in the nucleus, sustains stemness properties, and confers self-renewal; the total Oct4 primer sets, which detect both Oct4-A and Oct4-B expression (Mueller et al., 2009).
Quantitative RT-PCR was performed with Brilliant II Fast SYBR® Green QPCR Master Mix (Stratagene, La Jolla, CA, USA) and analyzed by the Mx3000P Real-Time PCR System (Stratagene). Each sample was run in triplicate. Cycling parameters were 95°C for 10 min followed by 40 cycles of 95°C for 15 sec, 60°C for 30 sec, and 72°C for 30 sec. Relative gene expression was analyzed with the comparative Ct method with glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as the endogenous control after confirming that the efficiencies of the target and the endogenous control applications were approximately equal. Results are presented as target gene expression normalized to that of GAPDH RNA. Values are the mean±standard deviation (SD) of triplicate samples from one representative experiment. Experiments T1 and T2 were completely independent from thawing hAMCs. The PCR primer sets used in RT-PCR and quantitative RT-PCR are shown in Table 1.
F, forward; R, reverse.
Immunocytochemistry
The cultured pCMV-hOct4 (5.3 kb) transfected and cardiac-differentiated hAMCs were fixed in 4% paraformaldehyde/PBS in eight-well chamber slides for 30 min at room temperature and permeabalized with 0.3% Triton X-100 (Sigma-Aldrich)/PBS for 10 min followed by blocking with 1% BlockingOne (Nacalai Tesque, Inc.) for 30 min. The cells were incubated with primary antibodies diluted in 1% bovine serum albumin (BSA; Sigma-Aldrich) and 0.5%Triton-X100 in PBS overnight at 4°C. The following primary antibodies were used to detect the reprogramming factors: Anti-Oct4 (diluted 1:200, clone H-134, Santa Cruz Biotechnology, Santa Cruz, CA, USA), anti-Oct4A (1:200, clone C-10, Santa Cruz), anti-Nanog (1:200, clone N-17, Santa Cruz), anti-Sox2 (1:50, clone 245610, BD Biosciences), and anti-Lin28 (1:50, BD Biosciences). The following primary antibodies were used to investigate the expression of cardiac-specific proteins: Anti-Nkx2.5 (1:200, clone N-19, Santa Cruz), anti-Connexin43 (1:50, Cell Signaling Technology
Statistical analysis
Statistical analyses were performed by Student t-tests. Variables were described by mean±SD. Statistical analysis was performed using StatView v4.58.0.0. p values less than 0.05 were considered statistically significant.
Results
Morphology and the expression status of reprogramming factors and cell-surface markers in hAMCs
Isolated and cultured hAMCs proliferated very well at least until the 10th passage (Fig. 1A). hAMCs just after isolation had a large cytoplasmic area, but during culture their morphology changed to small spindle-shaped cells (Fig. 1B). The immunocytochemistry of transcriptional factors used to produce iPSCs, called reprogramming factors, namely Oct4, Sox2, Klf4, and Lin28 (Takahashi et al., 2007; Takahashi and Yamanaka, 2006; Yu et al., 2007) in hAMCs, are shown in Figure 1C. Oct4, Sox2, Klf4, and Lin28 were detected in the nuclei of almost all hAMCs. Concerning Oct4, the expression of Oct4-A was detected in a minority of the cells, and its expression rate, calculated from three different views under fluorescence microscope, was 10.6%±6.5%. Nanog was detected in few hAMCs (data not shown). The expression of cell-surface markers in hAMCs detected by flow cytometry analysis is shown in Figure 1D. The hematopoietic markers CD14, CD34, CD45, and HLA-DR were not completely expressed. CD44 was detected in 95.8%±5.1% of hAMCs, CD73 in 6.8%±3.2%, CD90 in 66.0%±20.1%, and CD105 in 96.0%±1.6%; these mesenchymal stem cell markers had positive selection. On the other hand, the hESC-specific cell-surface marker SSEA-4 was detected in only 0.4%±0.2% of hAMCs.
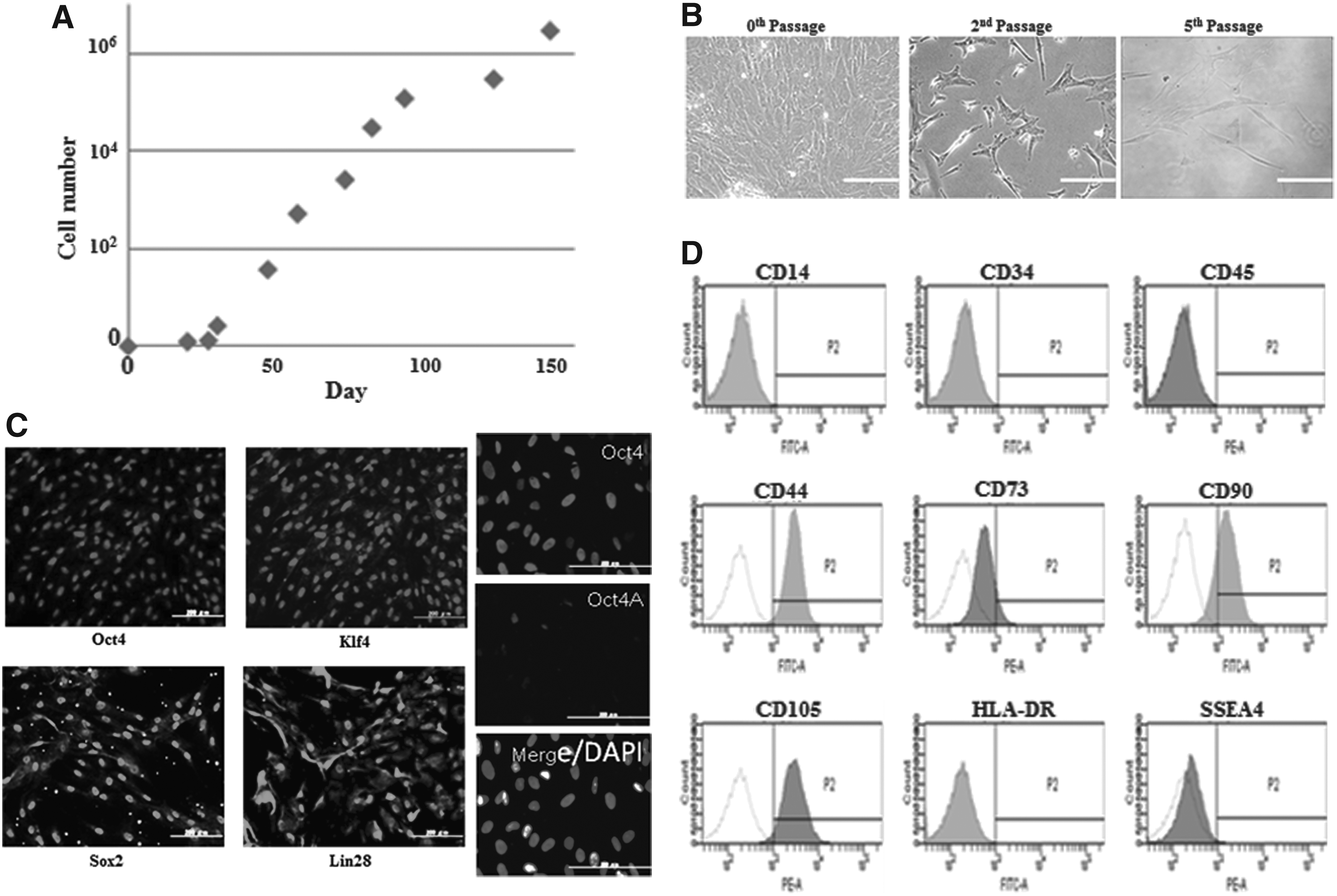
The characteristics of cultured hAMCs (proliferation potency, morphology, and expression status of stem cell–specific markers). (
Change of reprogramming factors expression after pCMV-hOct4 transfection
The transfection efficiency of hAMCs with pCMV-EGFP using Nucleofector Kit V (Lonza
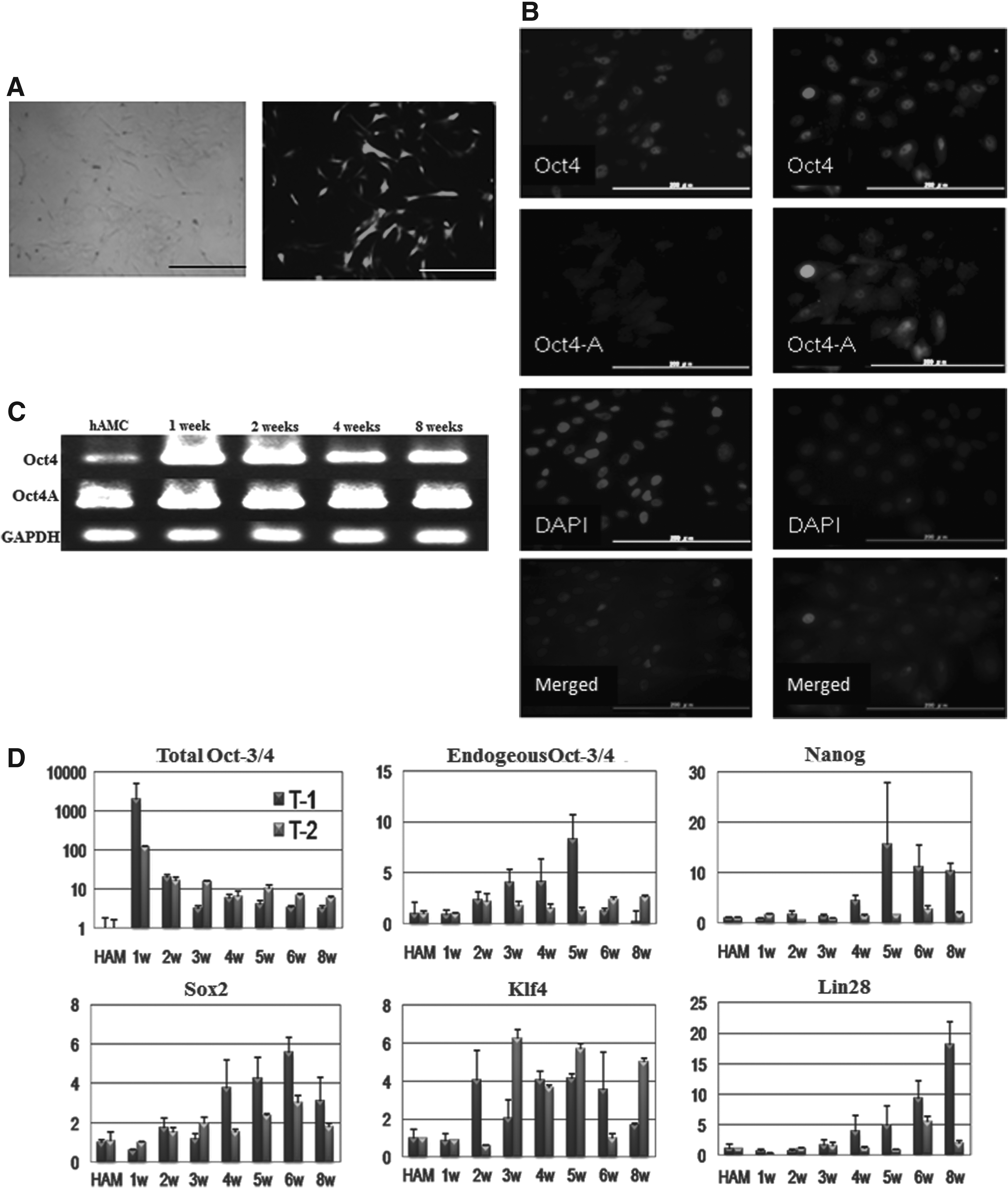
The changes in expression of reprogramming factors in hAMCs after pCMV-hOct4 transfection. (
The expression of cardiac-specific mRNAs in hAMCs after 5-Aza treatment
The cultured hAMCs originally expressed Nkx2.5, which is a cardiac-specific transcriptional factor, cardiac Troponin T (cTnT), and β-myosin heavy chain (βMHC). Four weeks after 5-Aza treatment, the expression of myosin light chain-2a (Mlc-2a) and myosin light chain-2v (Mlc-2v) was detected. Oct4 overexpressing hAMCs showed high expression of the differentiation markers Mlc-2a and Mlc-2v and also the cardiac-specific transcription factor GATA4 (Fig. 3A). According to the comparison of Mlc-2v and GATA4 between induced hAMCs and hAMCs after pCMV-hOct4 transfection by real-time PCR (Fig. 3B), the highest amount of GATA4 mRNA was detected when it was induced at 4 weeks after pCMV-hOct4 transfection in experiment T1 (27.9-±1.8-fold of induced hAMCs), and 8 weeks after in T2 (24.3-±8.0-fold). The highest amount of Mlc-2v mRNA was detected at almost same time as GATA4, 5 weeks after pCMV-hOct4 transfection in experiment T1 (23.3-±10.2-fold of induced hAMCs) and 8 weeks after in experiment T2 (75.7-±4.5-fold). On the other hand, we could not detect Nkx2.5 mRNA in the hAMCs treated with 5-Aza at 2 weeks or 8 weeks after pCMV-hOct4 transfection in experiment T1.
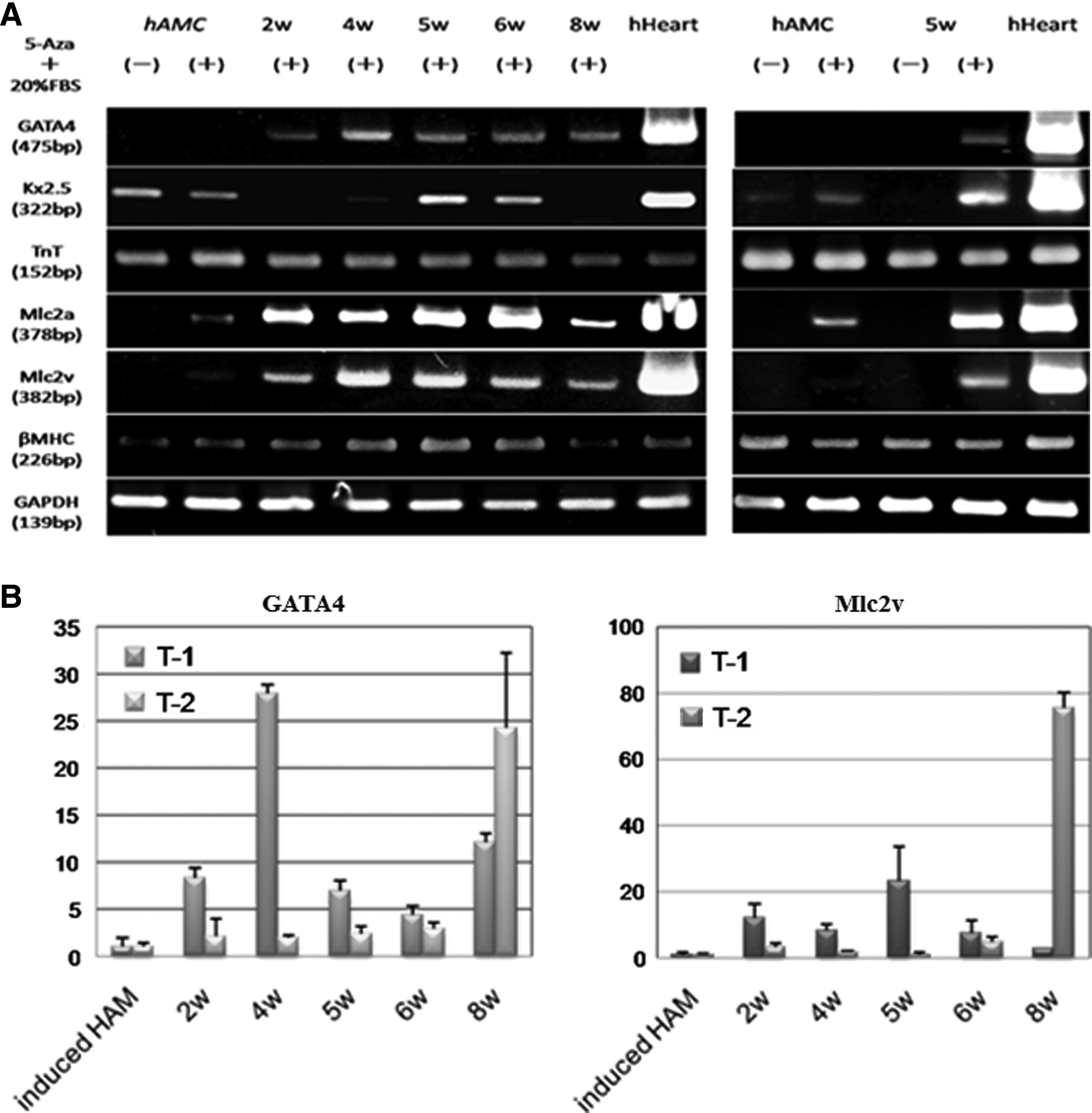
Expression of cardiac-specific mRNAs after 5-Aza treatment. (
Immunocytochemistry of the cardiac-specific proteins
hAMCs originally expressed Nkx2.5 protein in their nuclei. Connexin 43, a protein presenting in the gap junctions between adjacent cells that makes the transmission of the action potential during cardiomyocytes possible, was detected around hAMCs (Fig. 4A). Following 5-Aza treatment 4 weeks after transfection, some of hAMCs expressed cardiac MHC, Mlc-2v, and cTnT (Fig. 4B). As with the change of mRNA, the intensity of these cardiac-specific contraction proteins in pCMV-hOct4 transfected and then induced hAMCs was changed more than that of induced hAMCs without transfection. At the optimal induction time, 5 weeks after transfection in experiment T1, structure of cardiac MHC parallel to that in mature cardiomyocytes was observed (Fig. 4C).
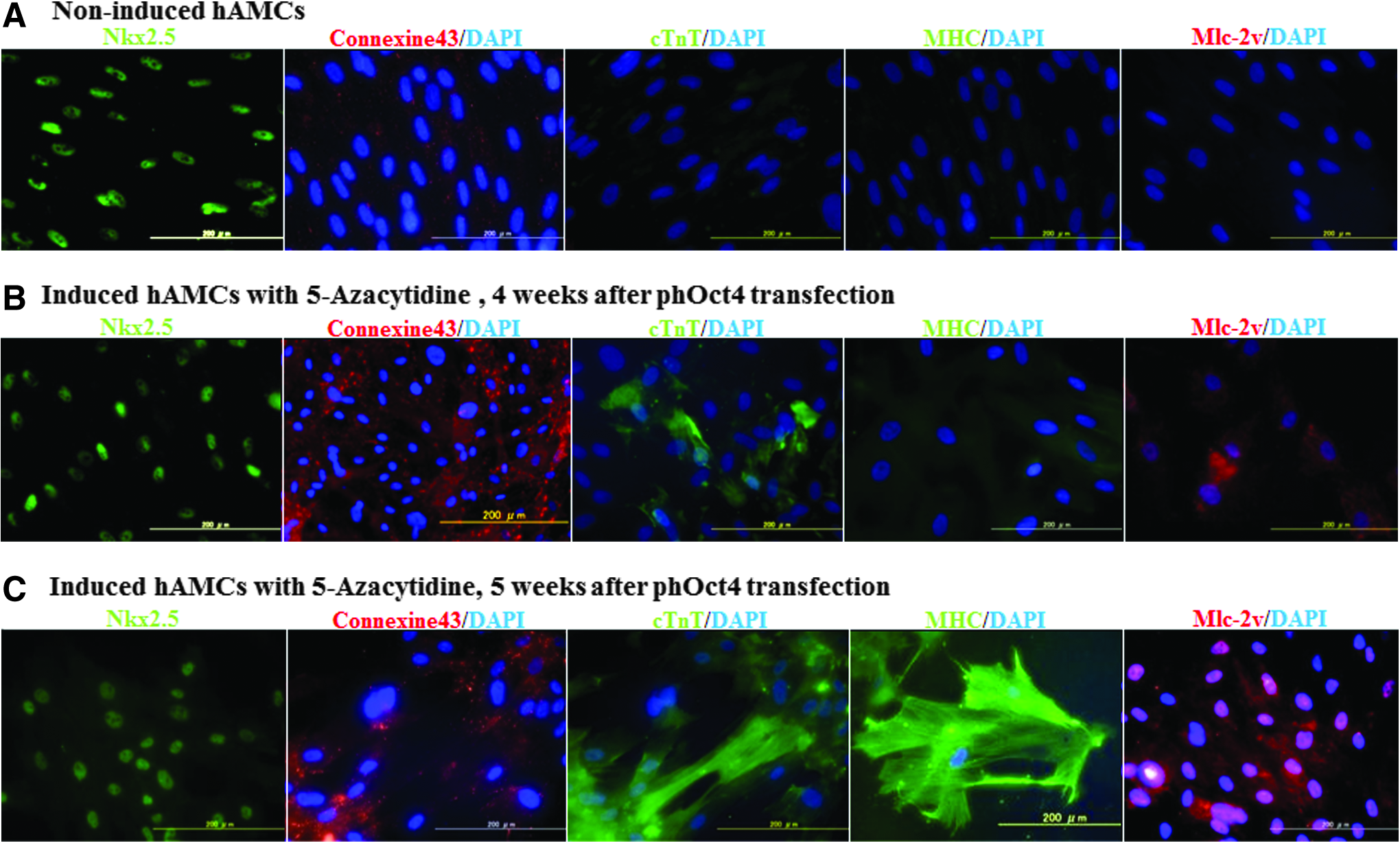
Immunocytochemistry of cardiac-specific makers. (
Discussion
Most of the isolated hAMCs expressed the stem cell–specific transcriptional factors Oct4, Sox2, Klf4, and Lin28, which are known reprogramming factors (Takahashi et al., 2007; Takahashi and Yamanaka, 2006; Yu et al., 2007). However, one of the stem cell–specific surface makers SSEA-4 was expressed in only 0.4% of hAMCs according to flow cytometry analysis. Oct4 is considered a main regulator of ESC pluripotency and self-renewal. Different mRNAs are generated by alternative splicing from the Oct4 gene (Takeda et al., 1992), resulting in two Oct4 protein isoforms: Oct4-A and Oct4-B. Only the Oct4-A protein sustains stemness properties and confers self-renewal; Oct4-B does not seem to function as a transcription factor (Cauffman et al., 2006; Lee et al., 2006). In our results of RT-PCR and immunocytochemistry with Oct4-A specific or nonspecific primer sets and antibodies (Mueller et al., 2009), Oct4 was detected in hAMCs, but the majority of them expressed Oct4-B. Therefore, most of the hAMCs do not have pluripotency even though they express most of the reprogramming factors.
The pCMV-Oct4 transfected hAMCs showed high expression of Oct4-A, and the expression of the other reprogramming factors was also increased after pCMV-hOct4 transfection. This change of these mRNAs might mean that the overexpressed exogenous Oct4-A worked to reprogram the hAMCs into an undifferentiated state. Recently, it has become evident that the transcriptional factor Oct4 lies at the center of a gene regulatory network that maintains pluripotency (Babaie et al., 2007; Boyer et al., 2005; Kim et al., 2008; Lengner et al., 2008; Nichols et al., 1998); for example, Oct4 and Sox2 interact with the Nanog promoter (Rodda et al., 2005). In such interactions, the expression of other reprogramming factors was induced by the overexpression of Oct4, and it is thought that Oct4-overexpressing hAMCs were activated into an undifferentiated state.
According to the results of RT-PCR and immunocytochemistry of cardiac-specific markers, Oct4 overexpression by plasmid electroporation not only brought hAMCs closer to the undifferentiated state, but also improved cardiac differentiation potency when they were induced at a specific time. hAMCs that induced cardiac differentiation at the optimal time showed high expression of cardiac differentiation markers and the fiber structure of contraction proteins, which seemed to be more mature contractile proteins. The optimal differentiation induction time was determined when high expression of endogenous Oct4 and Nanog, the target gene of Oct4, was shown in both experiments T1 and T2. The activation into the undifferentiated state in hAMCs contributed to the improvement of the cardiac differentiation potency of hAMC. According to reports of iPSC manufacture, it takes around 4 weeks for establishment of human iPSCs (Takahashi et al., 2007; Yu et al., 2007). Furthermore, it takes around 8 weeks or longer to obtain one-factor iPSCs (Kim et al., 2009a). On the basis of these reports, we consider that the optimal differentiation induction times in our experimental system is at 4–5 weeks from gene transfection in experiment T1 and at 8 weeks in experiment T2. We were convinced that these were the times necessary for hAMCs to be activated from the time of gene transfection into the undifferentiated state through the interaction of the variety of transcription factors.
However, the differentiation efficiency from the time of gene transfection with plasmid vector is poor. Because of this, it is thought that too-high expression of Oct4 disturbs cardiac differentiation. Actually, in the induction of differentiation at 2 weeks after pCMV-hOct4 transfection, not only is some cardiac-specific gene manifestation found to be low, but the expression of Nkx2.5 mRNA, which is originally expressed in hAMCs, is also lost. We suspected that the hAMCs are close to the undifferentiated state and lose the differentiation markers. Also, it is thought that the expression level of Oct4 itself has an influence on the differentiation efficiency. According to Rodriguez's work (Rodriguez et al., 2007), the reduced expression of Oct4 in hESCs promotes upregulation of the mesodermal markers and endodermal differentiation and the elevated level of Oct4 promoted upregulation of markers indicative of endoderm derivatives. On the other hand, Kim et al. reported that exogenous Oct4 improved the proliferation and differentiation potencies of adult adipose tissue stromal cells through epigenetic reprogramming of stemness genes into functional neurons, but the differentiation potencies into mesoderm were prominently attenuated (Kim et al., 2009b). We suspected that the changes of Oct4 expression in our system, the transient high expression of Oct4, and gradual decrease of the expression of exogenous Oct4 caused the improvement of cardiac differentiation potencies.
Although some improvement of the cardiac differentiation potency of hAMCs was seen in our system, we could not find automatically beating cells. We used 5-Aza and 20% FBS-containing medium for induction of cardiac differentiation. It has been reported that 5-Aza can act on mouse bone marrow mesenchymal stem cells to cause their differentiation into beating cardiomyocytes (Tomita et al., 2007), but there is no information regarding the conditions might enable us to produce beating cells from hAMCs. The amnion membrane does not enable delivery, so there is no infection when it is used after cryopreservation; in addition, there are ethical problems with the use of amnion. To make our system using hAMCs optimal, further investigations of differentiation procedures and induction times are needed.
In conclusion, the cryopreserved and then cultured hAMCs showed some stem cell–specific markers. Furthermore, Oct4 single overexpression by plasmid transfection contributed to activating hAMCs to the undifferentiated state through the transcriptional interaction of some reprogramming factors and improved their potency to cardiac differentiation.
Footnotes
Acknowledgment
This work was supported in part by a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Sciences, and Technology of Japan No. 20390366.
Author Disclosure Statement
The authors declare that there are no conflicts of interest.