
Abstract
Abstract
Fertilization in bovines causes profound changes in the epigenetic profile that affect both DNA methylation patterns and posttranslational histone modifications. These dynamic changes have a great potential for activating pluripotency genes and unfolding certain chromatin regions to recruit different transcription factors. Surprisingly, while the fundamental function of epigenetic remodeling is well understood, the bases of the process are still unknown. Recent developments in epigenetics suggest a multistep demethylation process that would imply the prior modification of the methylated cytosine or methyl group, followed by a DNA repair mechanism implicating enzymes such as activation-induced cytidine deaminase (AICDA) and ten-eleven translocation (TET) dioxygenase. Their functions seem to differ from one species to the other, and they are not yet well characterized in large mammals. Histones have, for their part, many associated and specific lysine demethylases (KDM). Their expression profile in large mammals is not well characterized. We have been interested in characterizing the spatiotemporal expression profile for each of the genes studied to increase our understanding of the molecular interactions following fertilization in early bovine embryo stages. Bovine oocytes and embryos at various preimplantation stages were collected following in vitro fertilization protocol. Total RNA for AICDA, TET1, TET2, TET3, KDM3A, KDM4A, KDM4C, and KDM5B was extracted, reverse transcribed into cDNA, and amplified by real-time PCR. Other embryo pools were collected, and protein localization of the genes studied was characterized. TET3 dioxygenase was present in the very first embryo stages, in contrast to TET1 and AICDA. Histone demethylases KDM3A, KDM4A, and KDM4C were expressed before and after embryonic genome activation, whereas KDM5B was mainly expressed during the blastocyst period. DNA demethylation following fertilization in bovines is not accomplished by AICDA but most probably by TET3. Histone demethylation is carried out by, among others, KDM3A, KDM4A, and KDM4C, which could act in sequence to demethylate histones prior to DNA demethylation of the female chromosomes.
Introduction
T
Epigenetic modifications consist of molecular marks affixed upon DNA and histone tails in a permanent or transitive manner to induce or inhibit gene expression. The fundamental role of epigenetic marks is to regulate gene expression in each cell and determine its functional specificity. Modifications are recognized by specific effector proteins that modify chromatin configuration in a permissive or repressive way, according to epigenetic marks (Couture and Trievel, 2006).
DNA methylation has long been known to be positively correlated with inhibition of gene transcription, mostly by preventing the binding of transcriptional machinery and factors (Bird and Wolffe, 1999). Nevertheless, we now know that the position of the methylation among a gene is the principal factor that determines the effect the epigenetic modification. For example, it has been shown that methylation of CpG-rich sites in gene bodies is present in highly expressed genes of human B cells (Rauch et al., 2009), and that a gene body of weakly expressed genes is hypomethylated in cancer cell lines (Shann et al., 2008). DNA methyltransferases (DNMT) have been known and studied for a long time and have been classified on the basis of their respective function. Maintenance DNMT were first discovered by Bestor who, in 1988, characterized DNMT1 (Bestor, 1988). This enzyme is active in the S phase of the cell cycle and adds methyl groups on neosynthesized DNA strands so that the methylation pattern on the parental strand is transferred to the daughter cells. The second class of DNMT includes methyltransferases that methylate de novo, that is, without following any preexisting pattern. These enzymes are DNMT3a, DNMT3b, and DNMT3L (Bourc'his et al., 2001; Kaneda et al., 2004).
The process of DNA demethylation in mammals and the enzymes involved remain unresolved (Nabel et al., 2012). Nevertheless, it is known that, in mice, epigenetic reprogramming capacity is maintained from oocyte to the two-cell embryo, suggesting a persistent presence of reprogramming enzymes at these stages (Bui et al., 2008; Egli et al., 2007; Egli et al., 2009; Yang et al., 2010). Some studies on zebrafish and mouse suggest a role for activation-induced cytidine deaminase (AICDA), a deaminase that could take out the amine group of the methylated cytosine to create a thymine–guanine mismatch that is further repaired by the base excision repair (BER) system and replaced by an unmethylated cytosine (Cortazar et al., 2011; Rai et al., 2008) (see Table 1 for information on genes). Other studies in mouse indicate that a newly discovered family of enzymes, the ten-eleven translocation (TET) dioxygenase, can hydroxylate the methyl group to counteract the recognition of the methyl group by effector proteins (Tahiliani et al., 2009). The TET family is composed of three members (TET 1–3) that first catalyze the transformation of 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC), followed by 5hmC to 5-formylcytosine (5fC) and 5-carboxycytosine (5caC) (Ito et al., 2011). It is suggested that hydroxylation silenced the methylation signal, implying that some methylation-repressed genes would be transcribed (Gu et al. 2011). Additionnally, considering that chromatin structure proteins are recruited on methylated CpG, it is plausible that hydroxylation could impair that process and contribute to chromatine reorganization (Thalhammer et al., 2012). It is also established DNA demethylation happens in an undirect way via the BER system (Dean, 2013). This repair system allows the replacement of methylated cytosine via the recognition of 5fC and 5caC by thymine DNA glycosylase (TDG). This enzyme is involved in the replacement of methylated cytosine (Cortazar et al., 2011) via the recognition of 5caC and 5fC by TDG.
OCT4, octamer-binding transcription factor 4; SOX2, sex determining region Y-box 2; HOXA1, homeobox A1; PRM1, protamine 1; TPNP1, transition protein 1; ARID, AT-rich interacting domain; 53BP1, tumor suppressor p53-binding protein 1; C5HC2, C5HC2 zinc-finger domain; TUDOR, Tudor domain; JARID, Jumonji domain–ARID-containing protein; JmjC, Jumonji C domain; JmjN, Jumonji N domain; PHD, plant homeobox domain; KLF4, Krüppel-like factor 4; ESC, embryonic stem cell, CXXC-type zinc-finger binding domain; pRB, protein retinoblastoma; Cys-rich, cysteine-rich region, methylcytosine dioxygenase catalytic domain; DSBH, double-stranded β-helixo, methylcytosine dioxygenase catalytic domain, metal binding; NLS, nuclear localization signal; APOBEC,apolipoprotein B mRNA-editing, enzyme-catalytic, polypeptide-like.
Histone lysine demethylases (KDM) are better characterized in comparison with proteins involved in DNA demethylation. Previous experiments performed in our laboratoty identified a few KDMs present in bovine oocyte (McGraw et al., 2007) and classified their profile into three groups. The first group represents genes that are both maternally and embryonically transcribed and includes KDM3A. The second consists of genes that are preferentially transcribed from the oocyte genome and poorly transcribed by the embryo, such as high-mobility-group nucleosomal binding domain 2 (HMGN2) (Vigneault et al., 2004b). The third group includes genes barely present before the maternal-to-embryonic transition (MET) that are highly expressed thereafter, suggesting a low or null participation of the protein in the first developmental stages. KDM5B was one of those genes. Several new KDMs are potentially present or involved in early lysine demethylation and have not been characterized in bovine embryos—KDM3A, KDM4A, KDM4C, and KDM5B.
Little is known about epigenetic modifications in early embryo development. In vitro fertilization and embryo transfer are now common techniques that are frequently unsuccessful because of our poor recognition of “good” versus “bad” embryos. Increasing our understanding of epigenetic modifications in the very first steps of life could help increase efficiency of assisted reproduction techniques. More specifically, the aim of our study was to determine which methylation-modifying enzymes are present in early-stage bovine embryos and to analyze their kinetics and cellular localization to assess their potential role in reprogramming both male and female gametes. Using qRT-PCR and immnunohistochemistry, the correspondence between mRNA expression/presence and protein expression of TET3, AICDA, TET2, TET1, KDM3A, KDM4A, KDM4C, and KDM5B during early developmental stages (oocyte to blastocyst) was established.
Material and Methods
All products were obtained from Sigma-Aldrich unless otherwise stated. Only in vitro embryos were used in this study.
Oocyte collection and in vitro maturation
The procedure for producing bovine embryos from slaughterhouse oocytes was essentially the same as published before (Cagnone et al., 2012). Briefly, 3- to 6-mm healthy follicles from dairy cow ovaries were aspirated to collect cumulus–oocyte complexes (COCs). After several washes in HEPES-buffered Tyrode lactate solution (TLH), COCs were selected on the basis of their visible quality—i.e., a minimum of three to four layers of cumulus cells surrounding the oocyte and absence of dark spots in the cytoplasm—and matured in groups of 10 in 50-μL drops of maturation buffer under mineral oil for 24 h at 39°C in a humidity-saturated atmosphere, under 5% CO2. The maturation medium was tissue culture medium-1999 (TCM-199) supplemented with 10% fetal bovine serum (FBS), 0.1 μg/mL follicle-stimulating hormone (Folltropin V; Bioniche), 0.33 mM pyruvic acid, and 50 μg/mL gentamicin.
In vitro fertilization
After maturation, expanded COCs were washed twice in TLH solution and transferred in groups of five to 48-μL drops of in vitro fertilization (IVF) medium under mineral oil. The IVF medium was composed of stock Tyrode lactate solution supplemented with 0.6% fatty acid-free bovine serum albumin (BSA), 0.2 mM pyruvic acid, 10 μg/mL heparin, and 50 μg/mL gentamicin. To each droplet, 2 μL of PHE (2 mM penicillamine, 1 mM hypotaurine, 250 mM epinephrine) was added to stimulate sperm motility.
The spermatozoa used consisted of a cryopreserved mixture of ejaculates from five bulls (Centre d'Insémination Artificielle du Québec). The spermatozoa were thawed in 37°C water for 1 min, put on a discontinuous Percoll gradient (2 mL of 45% Percoll over 2 mL of 90% Percoll), and centrifuged at 700×g for 30 min at 26°C. After discarding the supernatant, live spermatozoa were counted on a hemacytometer to obtain a concentration of 106 cells/mL and resuspended in IVF medium. Finally, 2 μL of the sperm suspension (final concentration=4×104 cells/mL) was added to each IVF droplet containing the matured COCs, and incubated in a humidified atmosphere at 38.5°C under 5% CO2 for 16–18 h.
In vitro culture
For embryo culture, a three-step modified synthetic oviduct fluid culture system containing minimum essential medium, essential and nonessential amino acids, 0.5 mM glycyl-glutamine, and 0.4% fatty acid-free BSA under embryo-tested mineral oil was used. The embryo culture dishes were incubated at 38.5°C with 6.5% CO2, 5% O2, and 88.5% N2 in 100% humidity. Briefly, after fertilization, presumptive zygotes were denuded mechanically and washed three times in TLH supplemented with fatty acid–free BSA and were placed in groups of 10 in 10-μL droplets of synthetic oviduct fluid 1 (SOF1) with nonessential amino acids (1×) and 3 μM EDTA. Embryos were transferred to new 10-μL droplets of SOF2 containing nonessential (1×) and essential (0.5×) amino acids 72 h postfertilization and once again 120 h postfertilization in 20-μL droplets of SOF3 containing nonessential (1×) and essential (1×) amino acids. Medium was replaced three times to prevent toxicity due to ammonium accumulation and nutrient depletion caused, respectively, by amino acid degradation and embryo metabolism. The glucose concentrations used in SOF1, SOF2, and SOF3 were, respectively, 0.2, 0.5, and 1.0 mM. Blastocyst development was assessed at day 7 postfertilization. Nonhatched blastocysts were transferred, washed three times in phosphate-buffered saline (PBS), collected in groups of 10 in small volumes of PBS into 0.5-mL microtubes, and stored at −80°C until RNA extraction.
RNA extraction and reverse transcription
Total RNA from each replicate was extracted from snap-frozen embryos conserved at −80°C for less than 2 months and purified using the PicoPure RNA Isolation Kit (Life Technologies). After DNase digestion (Qiagen), quality and concentration of extracted RNA was assessed by Bioanalyzer (Agilent). All extracted samples showed good quality with an RNA integrity number >7.5. One picogram of in vitro–transcribed green fluorescent protein (GFP) RNA was added before extraction as a technical external control for RNA extraction and reverse transcription.
The measured amounts of GFP in each pool at the end of the real-time PCR validate and measure the efficiency of the extraction and reverse transcription for each pool extracted, and this quantitative value was used to correct the levels of the other genes measured in these pools. Three pools of 10 germinal vesicule (GV) oocytes, metaphase II (MII) oocytes, or embryos of each stage were used for extraction. RNA and oligo(dT) primers (1 μM) were heated for 5 min at 65°C, then chilled on ice before the addition of the Sensiscript Reverse Transcriptase (Qiagen) components. The reverse transcriptase (RT) reactions were performed at 37°C for 2 h. For a detailed procedure, see Vigneault et al. (2004b). Oligo(dT) primers were used instead of random hexamers to measure the polyadenylated form of the mRNAs studied, which are related to the population of mRNAs susceptible to be translated into the corresponding protein.
Real-time PCR
The primers for TET1, TET2, TET3, AICDA, KDM3A, KDM4A, KDM4C, and KDM5B amplification were designed with IDT PrimerQuestSM software (Intergated DNA Technologies) using sequences from the National Center for Biotechnology Information (NCBI). Primers used and annealing temperature are listed in Table 2. For detailed LightCycler (Roche) procedure and quantification, refer to Vigneault et al. (2004b).
Statistical analysis
The level of mRNA for each gene subjected to statistical analysis was normalized using the GFP external control (McGraw et al., 2007; Vigneault et al., 2004b). External controls instead of internal controls have been used because of the difficulty of finding a gene that is stable enough from oocyte to blastocyst. Early embryonic development is very dynamic and dramatically modifies the embryo from one stage to another, thus external control is a better normalization solution (De Sousa et al., 1998; Vigneault et al., 2004a). The value obtained for each gene, within each pool of cDNA, was divided by the value obtained for GFP in the same cDNA pool. Statistical analysis was performed using Prism software (version 5.02, Graph Pad software Inc.). Relative mRNA abundance for a given gene was measured and compared between embryo stages from GV to the eight-cell stage using the Pearson test. We only considered those stages and omitted the blastocyst stage to compare maternal-origin mRNA before embryonic genome activation. Pearson correlations were based on a significance level of 0.05.
Immunoblotting
Immunoblotting was performed on bovine tissues to ensure the specificity of antibodies, except for KDM5B, which was tested on a HepG2 cell lysate because of its low expression level in other bovine tissues. Bovine tissues were directly lysed in 2× Laemmli buffer (Laemmli, 1970), resolved on standard 8% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gels, and transferred onto nitrocellulose membranes (Osmonics) using a semidry transfer apparatus following the Tris/CAPS discontinuous buffer protocol from BioRad (BioRad Laboratories). The transfer was performed at 400 mA/cm2 for 1 h at room temperature. Membrane blotting was performed as follows: The membrane was blocked in blocking solution composed of 5% of ECL Block (GE Healthcare) in Tris-buffered saline and Tween 20 (TBST; 0.1%) for 90 min at room temperature and then incubated with the first antibody overnight at 4°C. Membranes were washed three times for 15 min with TBST (0.1%), and incubated with secondary antibody goat anti-rabbit immunoglobulin G (IgG) (H+L) horseradish peroxidase (HRP) conjugate (Molecular Probes) for KDM4A, KDM4C, and TET3 and goat anti-mouse IgG (H+L) HRP conjugate (Molecular Probes) for KDM5B, and diluted 1:100,000 in 4% nonfat dry milk in TBST (0.1%). The membranes were washed three times with TBST (0.05%) and revealed with an ECL Advance Western Blotting Detection kit from Amersham (GE Healthcare). First antibody dilutions were 1:100, 1:500, 1:200, 1:200, and 1:500 for TET3, KDM3A, KDM4A, KDM4C, and KDM5B, respectively. TET3 (no. GTX121453) antibody was from GeneTex, 5hmC (no. ab106918, not tested in immunoblot but used in immunofluorescence) and KDM5B (no. ab56759) antibodies were from Abcam, KDM3A (no. AV32911) was from Sigma-Aldrich, KDM4A (no. 5328) was purchased from Cell Signaling, and KDM4C (no. bs-5934R) was from Bioss.
Immunofluorescence
GV- and MII-stage oocytes, two-, four-, and eight-cell embryos, as well as blastocysts were fixed in 4% paraformaldehyde (PFA)/PBS for 10 min at room temperature, washed several times in TBST (0.1%), and then permeabilized in 1% Triton X-100/TBST for 1 h. Tissues were washed again in TBST, and antigens were blocked in 5% nonfat dry milk/TBST for 1 h at room temperature. Antibodies were added to fresh blocking solution according to those concentrations: 1:1000, 1:2000, 1:1000, 1:1000, 1:500, and 1:250 for TET3, 5hmC, KDM3A, KDM4A, KDM4C, and KDM5B, respectively. Samples were incubated overnight at 4°C in antibody solution and washed in TBST several times. The secondary antibody (CF™ 555 Goat anti-rabbit/goat/mouse, corresponding to the species of the first antibody) was diluted at 1:2000 in freshly made blocking solution. Samples were incubated for 45 min followed by four washes of 15 min in TBST, and incubated in 4′,6-diamidino-2-phenylindole (DAPI) 1:1000 for nuclear staining. Samples were mounted on slides with 9 μL of PBS and observed with confoncal microscope LSM700 Zeiss (Zeiss).
Results
KDM3A, KDM4A, and KDM5B were selected in accordance with previous work in our laboratory (McGraw et al., 2007). We also selected KDM4C on the basis of an mRNA expression database of microarrays in bovine oocytes (McGraw et al., 2007). Other histones demethylases are present at the GV stage, but the lack of functional antibody limited our investigation.
Immunoblots
First, antibody specificity in bovine tissues was demonstrated on immunoblots (Fig. 1). All of the antibodies presented were positive for the protein in bovine tissue. A contaminating band located at 62 kDa was present on immunoblots of KDM4C and KDM5B and corresponds to BSA, epitopes of which are recognized by many antibodies.
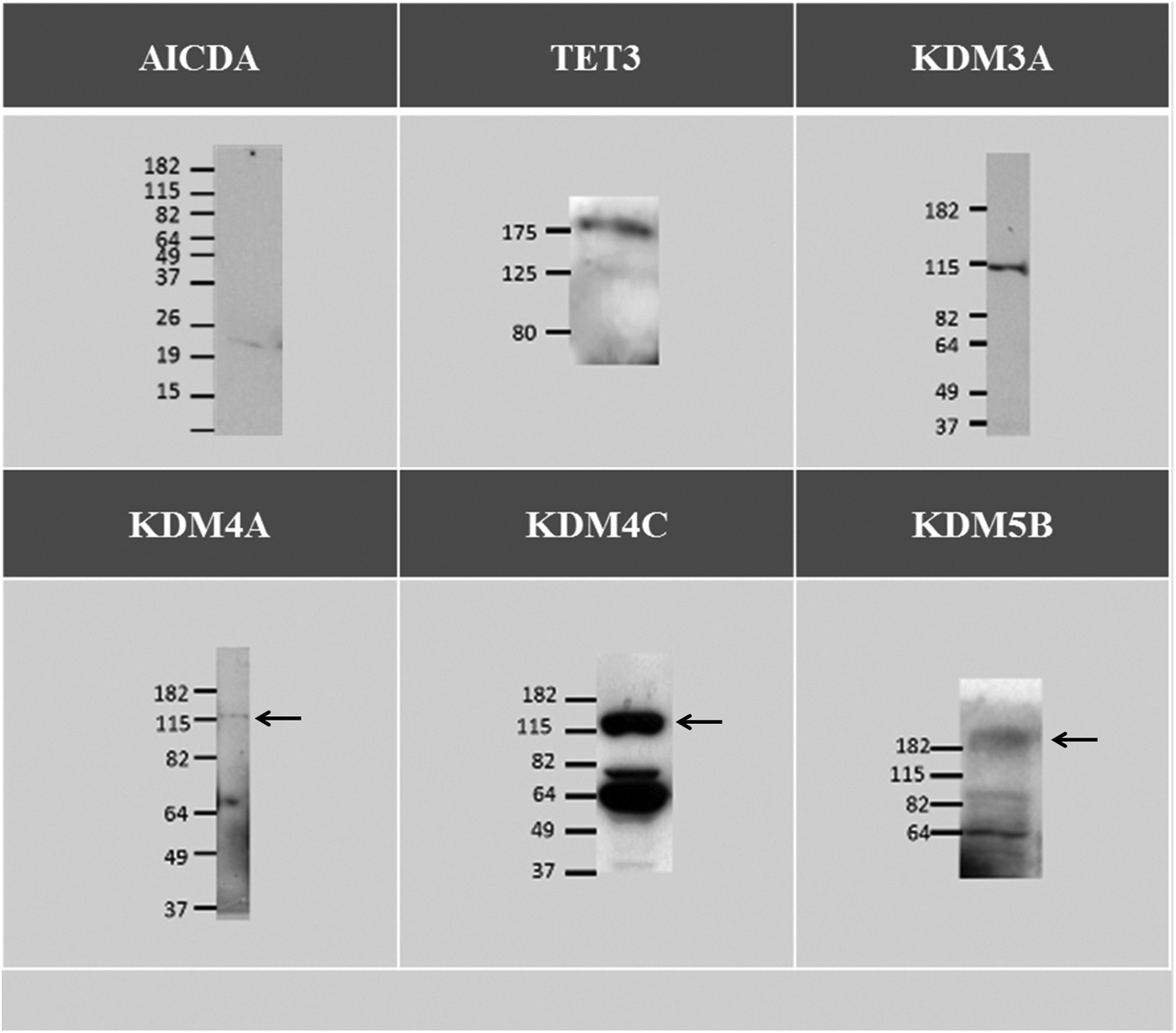
Immunoblot of AICDA, TET3, KDM3A, KDM4A, KDM4C, and KDM5B performed on 25 bovine GV oocytes, 50 bovine GV oocytes, 10 μg of bovine ovary lysate, 10 μg of bovine oviduct lysate, 10 μg of bovine ovary lysate, and 15 μg of HepG2 lysates, respectively. The arrows indicate the specific band.
Using real-time PCR, it was impossible to detect any TET1 or AICDA transcripts in our samples. To confirm these results, immunofluorescence and western blot protocols were performed. Unfortunately, no TET1 antibody was available for bovine tissue. Surprisingly, immunoblotting revealed a weak band suggesting the presence of some AICDA protein (Fig. 1) despite the undetected mRNA. Analysis of oocytes from 1- to 2-mm follicles failed to reveal AICDA mRNA as well (results not shown). We performed real-time PCR of TET2 on the same samples and obtained significant results (data not shown) indicating that TET2 is present in GV stage and decreased until the eight-cell stage. However, its level of expression in very low (1:100 compared to TET3).
TET3 is present at the time of paternal demethylation
Real-time-PCR experiments for TET3 at different embryo stages showed an mRNA profile similar to genes of maternal origin (Fig. 2). The peak level of mRNA quantification was reached at the GV and two-cell stages and continuously decreased from there to MET. At the eight-cell-stage, mRNA was barely detectable and remained low at the blastocyst stage. From GV to the eight-cell stage, the Pearson determination coefficient was 0.3901 (p value 0.0299) (Table 3). Immunofluorescence pictures of TET3 support that tendency (Fig. 3). At the GV stage, TET3 localization was diffuse in the cytoplasm and showed low predominance in the nucleus. At the MII stage, TET3 seemed to be relocate into the nucleus and was closely associated to chromatin, which is condensed at that precise moment. Interestingly, an unexpected pattern appeared at the pronucleus stage. TET3 was present throughout the paternal pronucleus, with two spots intensely colored, but was also present in the maternal pronucleus in the form of a condensed spot. The protein was then excluded from the nucleus at the two-cell stage and was no longer present at the eight-cell stage.

Relative abundance of TET3 transcripts through early developmental stages as shown by qRT-PCR analysis. Statistical analysis shows a Pearson determination coefficient (r2) of 0.3901 and a p value (two-tailed) of 0.0299.
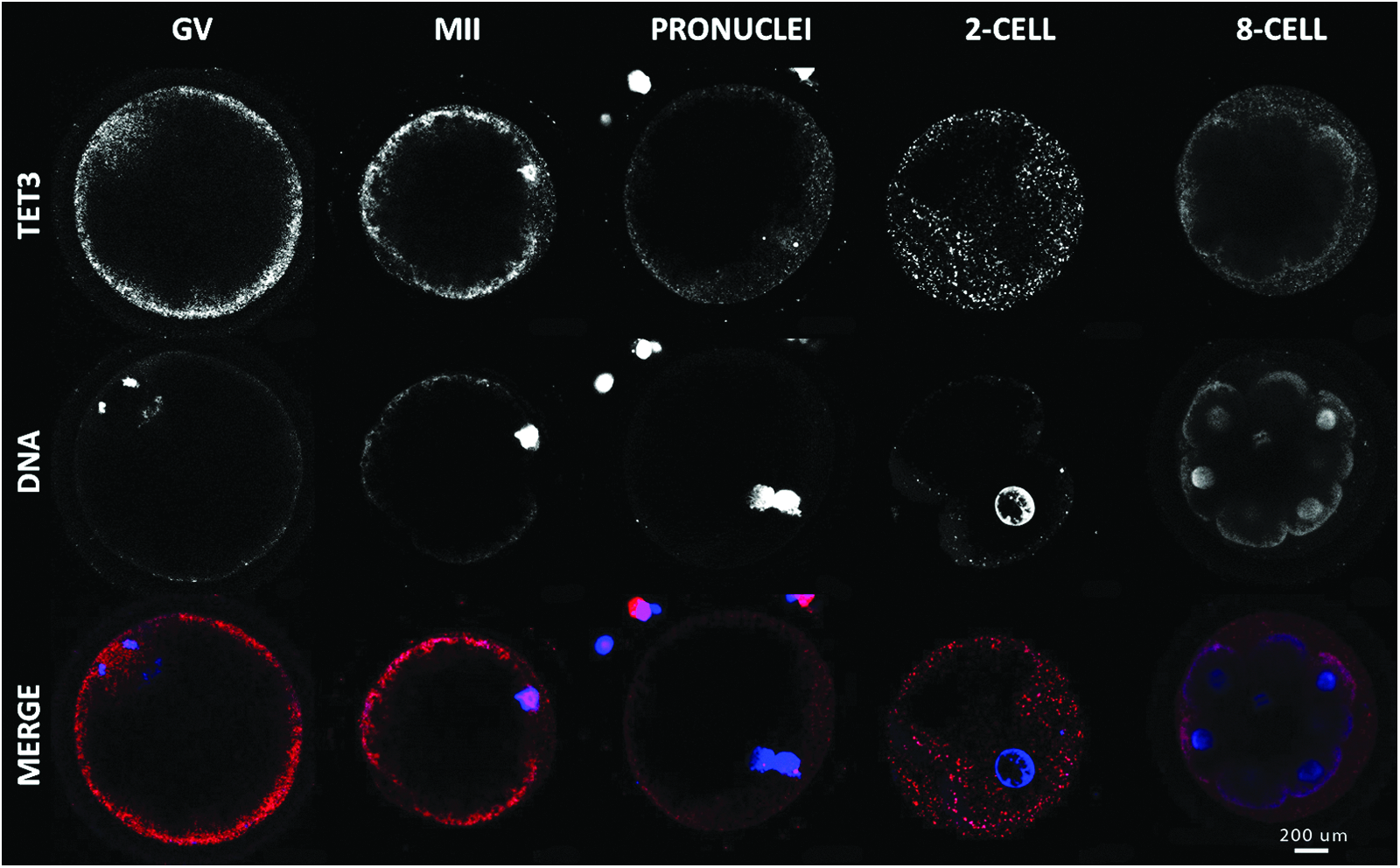
Immunolocalization of TET3 in oocyte and early embryo development. Confocal representation of oocytes (GV and MII) and embryos (male pronucleus, male and female pronucleus, two-, early eight-cell) stained in red with anti-TET3 antibody and in blue with DAPI to visualize the DNA. Magnification, 400×. Color images available online at www.liebertpub.com/cell
Immunofluorescence of 5hmC showed a similar pattern (Fig. 4). The GV stage was labeled by two punctuate spots in the nucleus and a single spot in the maternal pronucleus. 5hmC was spread out in the paternal pronucleus and was maintained at later stage.
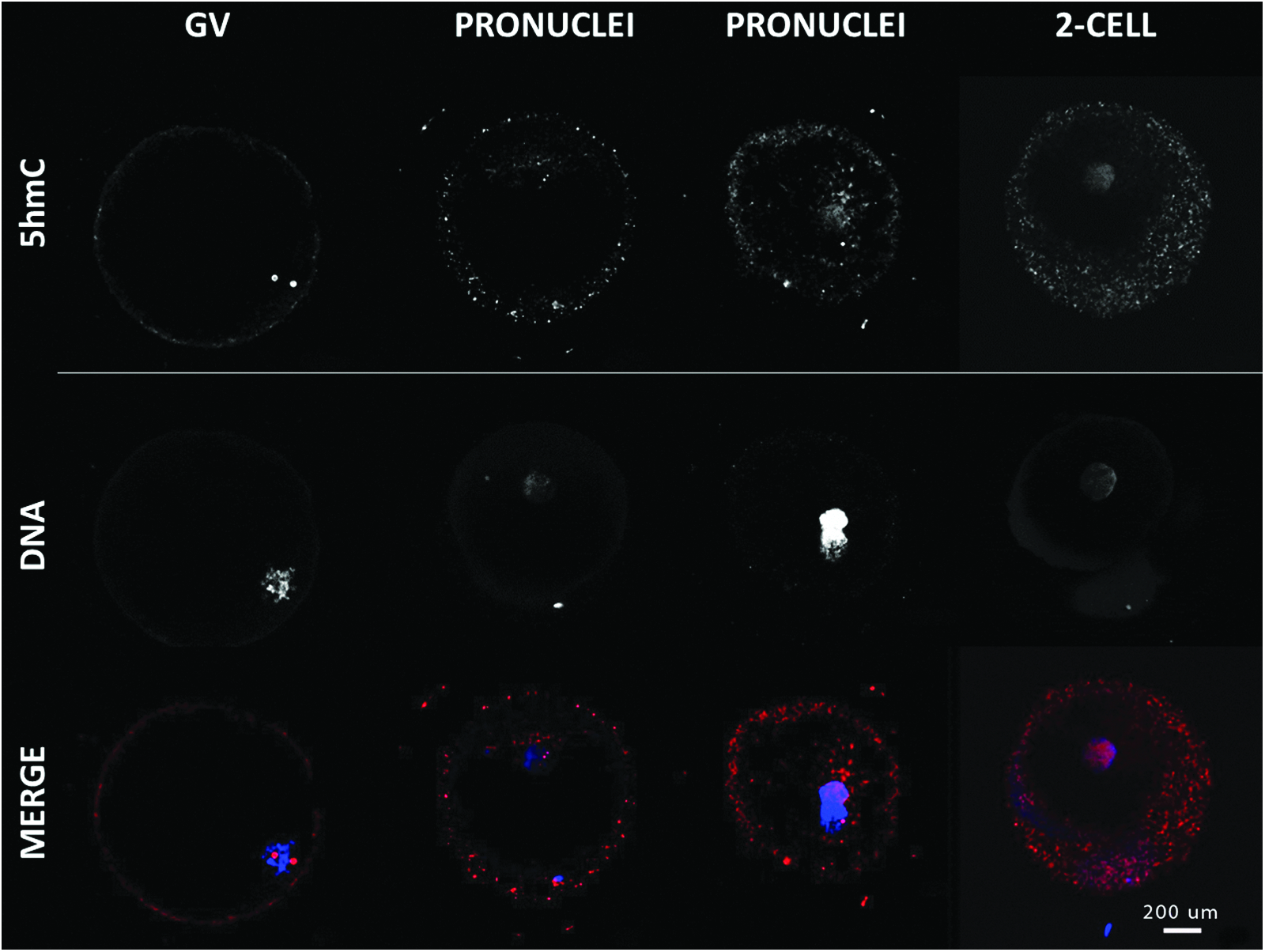
Immunolocalization of 5hmC in oocytes and early embryo development. Confocal representation of oocytes (GV and MII) and embryos (pronucleus, two-cell) stained in red with anti-5hmC antibody and in blue with DAPI to visualize the DNA. Magnification, 400×. Color images available online at www.liebertpub.com/cell
KDM3A is stored in the oocyte
As revealed by qRT-PCR analyses, KDM3A mRNA was stored in oocytes before final maturation. The maternal stock gradually decreased until MET, which was followed by embryonic expression of the gene (Fig. 5). The decrease during the early stages was validated by a determination coefficient of r2=0.7408. This coefficient was based on GV to eight-cell stages and means that almost 75% of the variation in mRNA abundance may be explained by the progression of embryo in normal development. Post-MET gene expression increased and became similar to GV-stage abundance, which is in agreement with results obtained by McGraw et al. (2007). KDM3A was present at all embryo stages studied and localized into the nucleus, except during GV, where it was also partially present in the cytoplasm (Fig. 6). Soon after fertilization, its presence was restricted to the male pronucleus but became ubiquitous at subsequent stages.

Relative abundance of KDM3A transcripts through early developmental stages as shown by qRT-PCR analysis. Statistical analysis shows a Pearson determination coefficient (r2) of 0.7408 and a p value (two-tailed) of 0.0003.
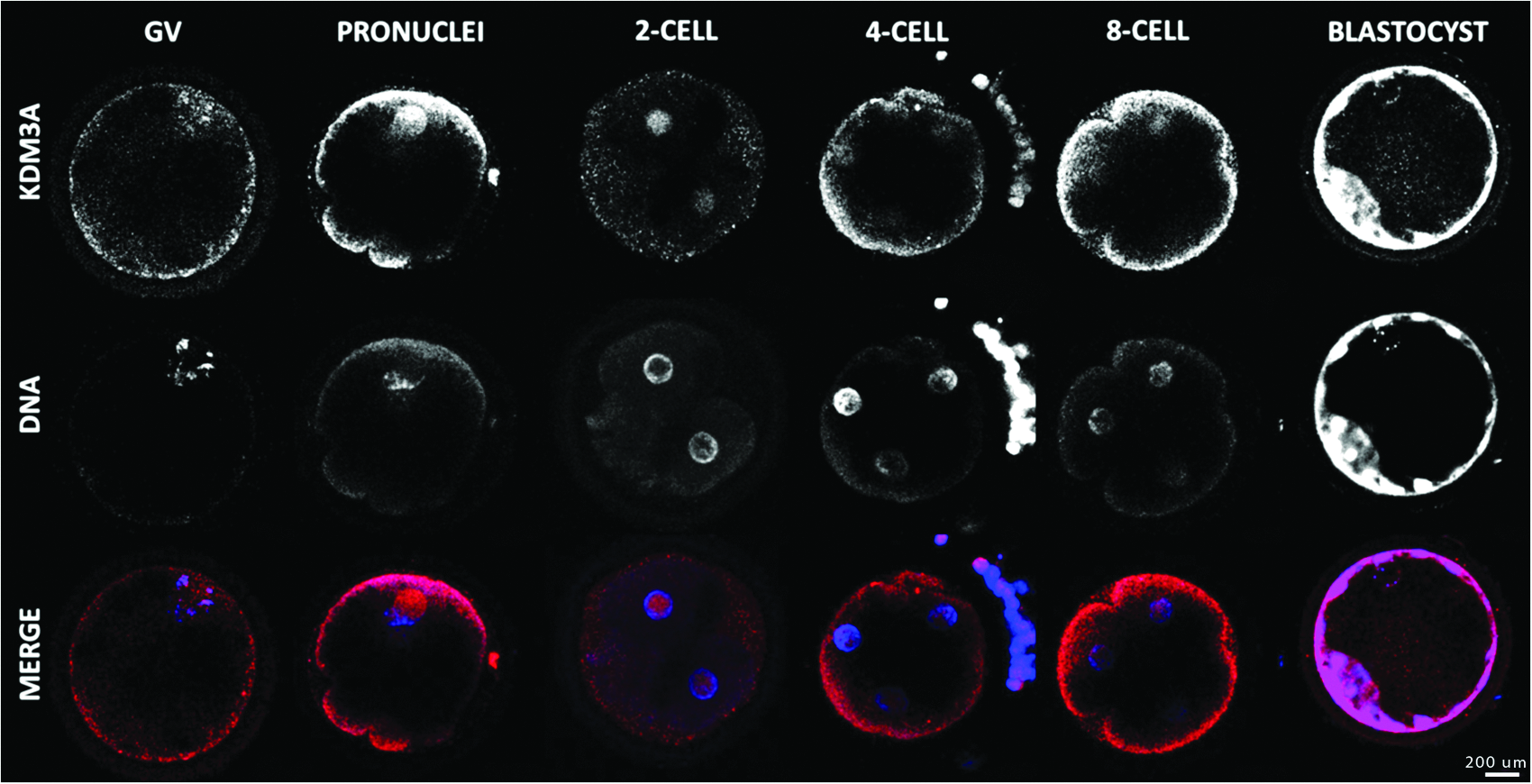
Immunolocalization of KDM3A in oocyte and early embryo development. Confocal representation of oocytes (GV) and embryos (pronucleus, two-, four-, eight-cell, and blastocyst) stained in red with anti-KDM3A antibody and in blue with DAPI to visualize the DNA. Magnification, 400×. Color images available online at www.liebertpub.com/cell
KDM4A has both maternal and embryonic origin
Similar to KDM3A, the KDM4A mRNA pattern corresponded to genes of both maternal and embryonic origin. KDM4A mRNA transcripts accumulated in GV oocytes and were for the most part lost during later cleavage stages (Fig. 7). However, transcription was reinitiated in the embryo after MET and allowed mRNA to reach levels similar to those observed at the GV stage. The mRNA profile from GV to eight-cell embryo was described by a Pearson determination coefficient of 0.6231 (double-tailed p value of 0.0023). Immunofluorescence experiments of KDM4A showed its exclusively nuclear presence throughout the bovine preimplantation stages. At GV and MII stages, the protein strictly co-localized with DAPI. In subsequent stages, the labeling is diffused in the nucleus (Fig. 8).

Relative abundance of KDM4A transcript through embryo development as shown by qRT-PCR analysis. Statistical analysis shows a Pearson determination coefficient (r2) of 0.6231 and a p value (two-tailed) of 0.0023.
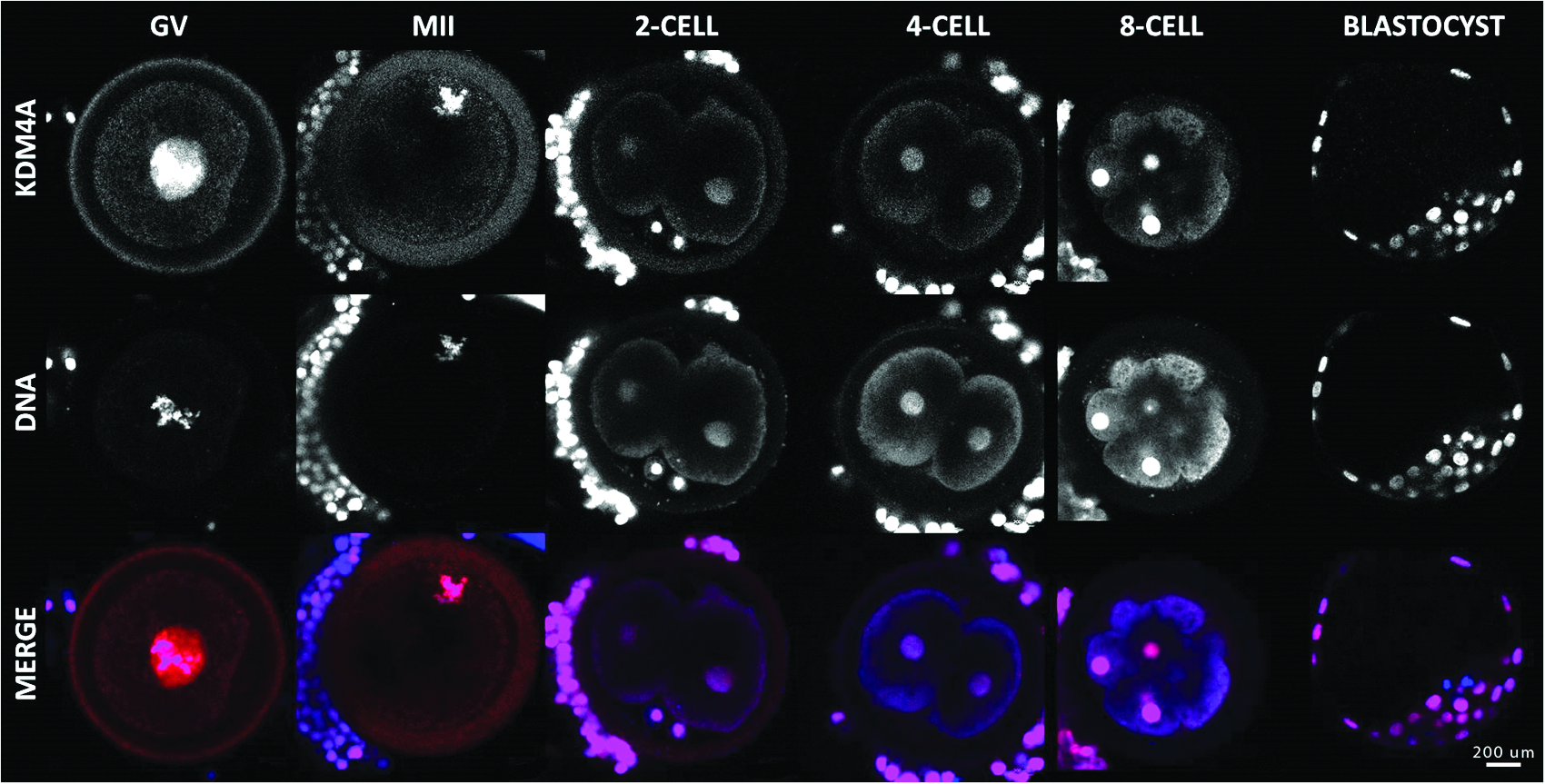
Immunolocalization of KDM4A in oocyte and early embryo development. Confocal representation of oocytes (GV and MII) and embryos (two-, four-, eight-cell, and blastocyst) stained in red with anti-KDM4A antibody and in blue with DAPI to visualize the DNA. Magnification, 400×. Color images available online at www.liebertpub.com/cell
KDM4C is present during MET
Both KDM4C protein localization and transcripts pattern in early embryo stages were very different from the KDM3A and KDM4A patterns. First, as opposed to KDM3A and KDM4A, KDM4C was weakly present before MET. Second, faint nuclear labeling of KDM4C protein was detected in oocytes, followed by a very specific punctuate labeling in both pronuclei (Fig. 9). The particularity of this labeling localization was enhanced by the presence of two distinct dots in each of the pronuclei. Because maternal and paternal DNA are replicated before fusion, the two dots in each pronucleus suggest binding to the two alleles of a same locus. During this pre-MET period, the mRNA abundance was characterized by a Pearson coefficient of correlation of 0.5915. After MET, both transcripts and protein levels were upregulated (Fig. 10).
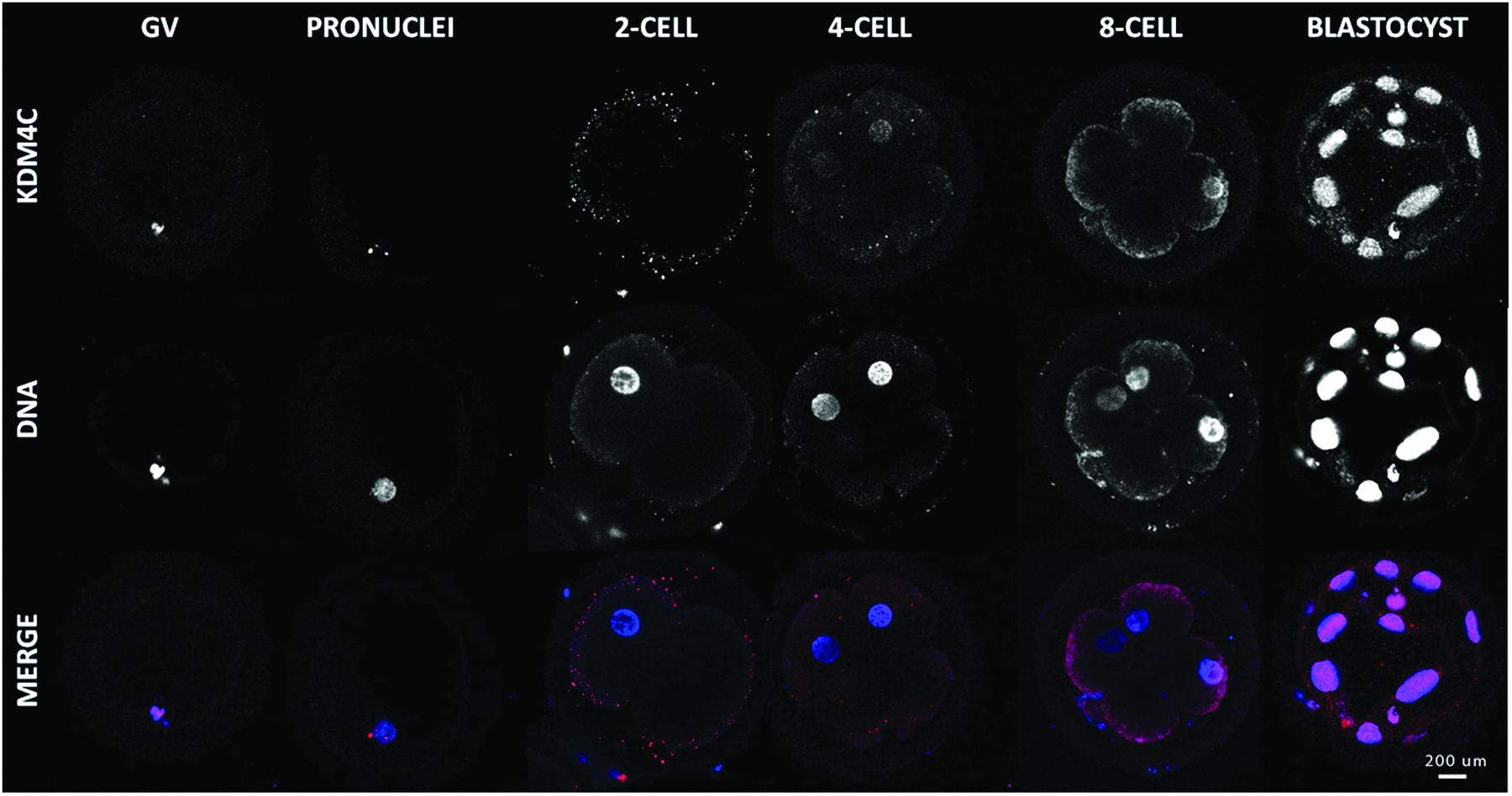
Immunolocalization of KDM4C in oocyte and early embryo development. Confocal representation of oocytes (GV) and embryos (pronucleus, two-, four-, eight-cell, and blastocyst) stained in red with anti-KDM4C antibody and in blue with DAPI to visualize the DNA. Magnification, 400×. Color images available online at www.liebertpub.com/cell

Relative abundance of KDM4C transcripts through early developmental stages as shown by qRT-PCR analysis. Statistical analysis shows a Pearson determination coefficient (r2) of 0,5915 and a p value (two-tailed) of 0.0035.
KDM5B has embryonic origin
No KDM5B mRNA transcript was detected at any of the stages studied before MET (results not shown). Later during development, the relative transcript level was significantly higher in blastocysts than in the eight-cell embryos. This profile indicates that KDM5B could be more important in the embryo during or after MET than in the first cells' cleavage. Immunofluorescences analyses showed absence or weak presence of the protein from the pronucleus stage to the four-cell embryo that is markedly increased at the eight-cell stage (Fig. 11). The pronounced staining in oocytes despite weak mRNA abundance could be explained by the residual protein presence from earlier maturation stages or the translation of the residual mRNA.
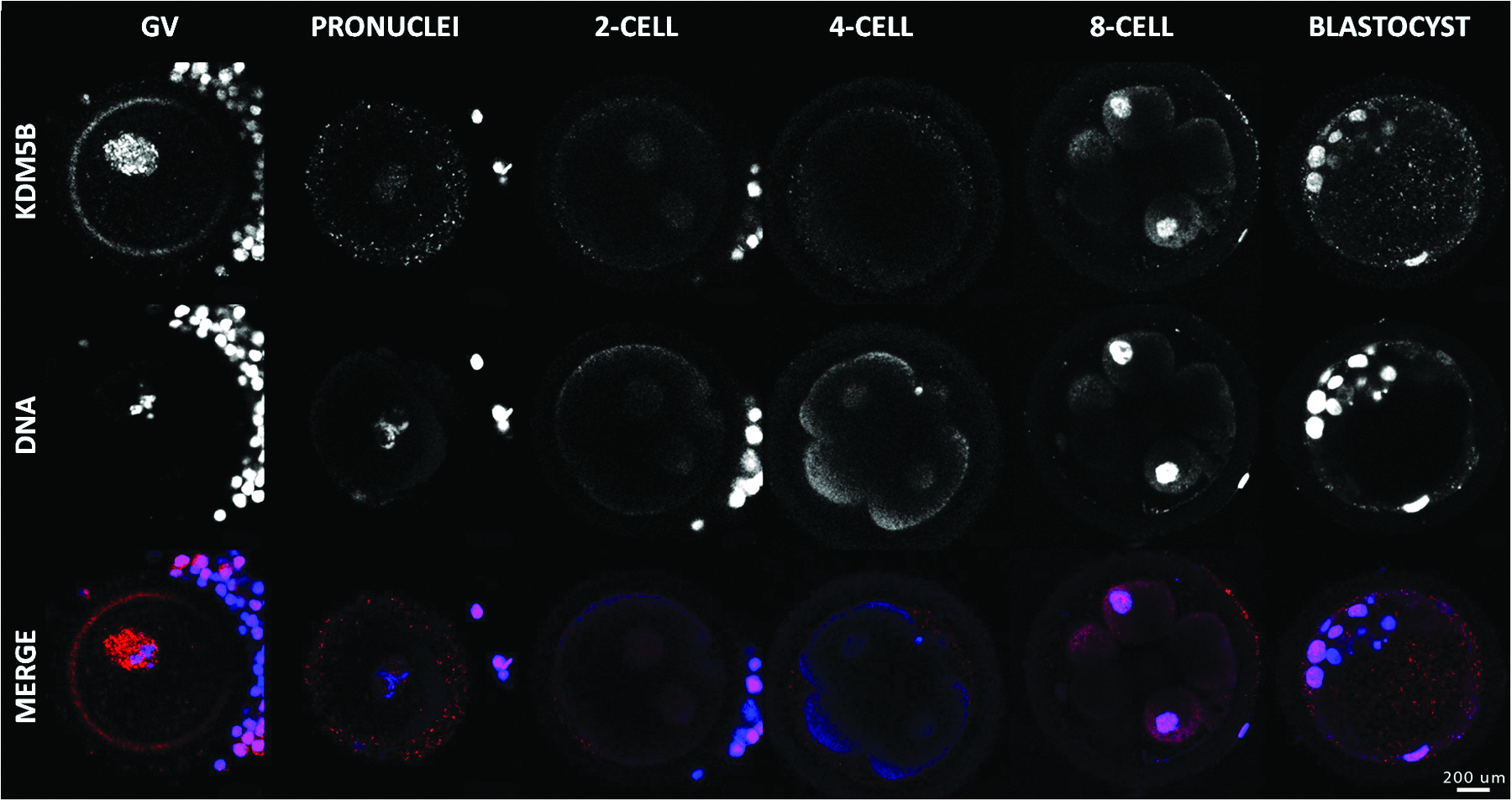
Immunolocalization of KDM5B in oocyte and early embryo development. Confocal representation of oocytes (GV) and embryos (pronucleus, two-, four-, eight-cell, and blastocyst) stained in red with anti-KDM4C antibody and in blue with DAPI to visualize the DNA. Magnification, 400×. Color images available online at www.liebertpub.com/cell
Discussion
Our results demonstrate that neither AICDA nor TET1 seem to be involved in DNA demethylation following fertilization. This function would, at least in part, be initiated by hydroxylation activity of TET3 dioxygenase on methylated cytosine. This study also demonstrates that many histone demethylases act at different key moments during bovine development. Those key moments are GV, pronucleus (before maternal and paternal genome fusion), MET transition at the eight-cell stage, and the first differentiation step at the blastocyst stage.
TET3 may be the major DNA demethylation enzyme
For paternal DNA, fertilization is the starting point of a myriad of rapid and coordinated modifications, of which protamine replacement by maternal histones is the very first step. After DNA decondensation, cytosine demethylation is activated. In mammals, thymine DNA glycosylase (TDG) and methyl-binding domain 4 (MBD4) are the main glycosylases. The essential role of TDG in embryo development has been shown in mice, whereas its absence (TDG null mice) was associated with epigenetic aberrations on development genes (Cortazar et al., 2011). Nevertheless, neither TDG nor other mammal glycosylase presented affinity for 5mC.
AICDA is one of these enzymes that modifies methylated cytosine and enhances their affinity for glycosylase. AICDA is a well-known deaminase that was first identified in immature B lymphocytes playing a central role in immunoglobulin class changes and somatic hypermutation (Neuberger et al., 2003). It is now accepted that AICDA is present in oocytes, embryonic tissues, and primordial germ cells (PGC) in mouse as well as in zebrafish oocytes (Rai et al., 2008). This gene is part of a pluripotency genes cluster that includes STELLA, NANOG, and CpG-restricted genes (Morgan et al., 2004). It was expected that AICDA could contribute to epigenome reprogramming, but null mice for AICDA do not present any particular phenotype except for immunoglobulin production and are still fertile (Muramatsu et al., 2000). Thus, an AICDA contribution would not be essential to embryonic and fetal development in mice (Muramatsu et al., 2000). In bovines, the very low expression level of AICDA at each embryonic stage combined with the absence of immunofluorescence signal support the inactivity of AICDA in the epigenetic reprogramming process following fertilization in this species.
Another enzyme family that acts on methylated cytosine is TET. This family is composed of three dioxygenases (TET1–3) that hydroxylate the methyl group to form 5hmC. A second and third hydroxylation is then possible to form 5fC (Pfaffeneder et al., 2011) and 5caC, respectively (He et al., 2011). Even though mammalian glycosylase recognizes 5hmC, 5fC and 5caC have higher turnover rates. 5-Hydroxymethyluridine, formed by the combined activity of deaminase and dioxygenase, is also rapidly metabolized by glycosylase.
Not all members of the TET family participate in demethylation in the embryo, as shown by our mRNA abundance study. Only TET2 and TET3 were detectable in the early embryo with a predominance of TET3 (1:100). In accordance with a previous study in bovines (Wossidlo et al., 2011), we suggest that TET3 is one of the major actors of the reprogramming process, and that TET2 is not. Furthermore, it has been shown that the normal demethylation process of the POU5F1 and NANOG genes is compromised in the male pronucleus of TET3−/− mice, where its methylation level was similar to the female allele of these genes (Gu et al., 2011; Iqbal et al., 2011). This underscores the importance of TET3 in active demethylation following fertilization and suggests its special role in epigenome reprogramming in mice.
On the basis of immunofluorescence labeling, TET3 was present in oocytes in the form of two distinct dots maintained until late pronocleus stage. We found a similar labeling with 5hmC antibody. Consequently, we concluded that some targeted locus might be actively demethylated in the maternal pronucleus. The paternal genome seems to be more sensitive to TET3 hydroxylation because the labeling was diffuse rather than specific. To explain the nonactive demethylation of maternal pronucleus in mice, Nakamura et al. (2012) have proposed a protective effect of STELLA on H3K9me2 (Nakamura et al., 2012). This epigenetic mark is tightly associated with methylated cytosines and must be removed to allow cytosine demethylation. Considering that H3K9me2 is more specifically localized in maternal pronucleus (Nakamura et al., 2012), it has been shown that STELLA protects H3K9me2 from demethylation and, therefore, prevents cytosine demethylation. Nakamura et al. also showed the very low level of H3K9me2 in the male pronucleus except for two specific dots.
Knowing that over 80% of imprinted genes are grouped into DNA clusters (Reik et al., 2001), it is proposed that those two dots could correspond to some of those imprinted genes clusters whose methylation must be conserved. In addition, Wossidlo et al. (2011) recently observed that, in bovines as well as in mice and rabbits, the male pronucleus was globally marked with 5hmC, whereas the maternal pronucleus was largely marked with 5mC. In that perspective, we propose that TET3 would be the very first enzyme of the active demethylation process in the bovine embryo and acts early in the male pronucleus. Female pronucleus demethylation would follow a passive process that implies the exclusion of methylation enzymes from the nuclei to avoid maintenance of DNA methylation on the newly synthesized DNA strand. Considering that the vast majority of maternal genome methylation sites remain stable during early embryo development (Smith et al., 2012), it might be possible that the DNA methylation process is exclusively achieved in a passive way.
KDM3A has different function depending on the embryo stage
The presence of KDM3A in every embryo stage studied corroborates real-time PCR results and suggests a broad involvement of the enzyme in the epigenetic remodeling process. KDM3A demethylates H3K9me2, an epigenetic mark associated with DNA methylation and, thus, gene repression. Therefore, the removal of this epigenetic mark implies DNA demethylation and further gene activation. This idea is supported by the peculiar localization of KDM3A that has been observed in both pronuclei. Indeed, the exclusion of KDM3A from the female pronucleus has been noted, which suggests that this exclusion could be linked to passive rather than active demethylation of the female pronucleus. Unfortunately, it was not observed in all samples and should be considered as a hypothesis. As previously seen in mouse, STELLA was mainly present in the maternal pronucleus and, by binding H3K9me2, could prevent cytosine demethylation. More specifically, binding of STELLA to H3K9me2 could interfere with KDM3A binding and thereby protect this epigenetic mark from demethylation.
In addition to its possible role at the moment of male pronucleus demethylation, KDM3A was still present after MET, suggesting additional functions. Real-time PCR results showed a very low mRNA level between the two- to eight-cell stages, and immunofluorescence data suggested a similar trend at the protein level. Considering the study of Loh et al. (2007) in which KDM3A knockout embryos were unable to differentiate and were stuck at the primordial endoderm stage, it is believed that the demethylase activity of KDM3A on H3K9me2/me1 allows the transcription of genes involved in differentiation and contributes to define cell identity.
KDM4A has multiple functions throughout embryo development
KDM4A has several functions in early development. First, by removing one methyl mark from H3K9me3 and from H1.4K26me3/me2 (Trojer et al., 2009) that are associated with heterochromatin, KDM4A antagonizes heterochromatin expansion. Moreover, when highly expressed, the protein has the potential of making heterochromatin regress due to delocalization of HP1, a protein associated with H3K9me3 (Black et al., 2010). Pursuing this further, KDM4A expression can be associated with embryonic genome activation, the moment when a new epigenetic profile is installed. For this purpose, KDM4A could allow some genes, such as pluripotency genes, to stay in a permissive state and could also prevent heterochromatin expansion. In addition to these two epigenetic targets, the demethylase recognizes H3K36me3 (Cloos et al., 2006). This epigenetic mark is associated with gene transcription, whereas its removal leads to transcriptional downregulation. Nevertheless, genes where the epigenetic state of histones changes from H3K36me3 to H3K36me2 stay in an open chromatin state. KDM4A is consequently associated with active genes and is present throughout embryo development.
KDM4C function is different in bovines than in mice
KDM4C is a demethylase specific to H3K9me3 and H3K36me3. In mice, its expression is exclusively embryonic and is first detected at the two-cell stage (Wang et al., 2010). In contrast, bovine expression of KDM4C is both maternal and embryonic. At the pronucleus stage, precise localization of the protein suggests a role for the enzyme in activating some key genes. It has already been demonstrated in mice that KDM4C allows transcriptional regulation of the major pluripotency genes POU5F1, NAGOG, and SOX2, as well as that of the cellular proliferation genes MYC and KLF4. Subsequent embryo stages were characterized by the disappearance of the protein until embryonic genome activation at the late eight-cell stage. Considering the very defined localization of the KDM4C protein in the pronucleus, this enzyme could act at that early stage to activate some of the proliferation and pluripotency genes.
KDM5B is part of MET transition
We have shown that KDM5B was present in the oocyte, even though its mRNA was weakly abundant before MET, and that it was significantly upregulated past the eight-cell embryo. KDM5B demethylates H3K4me3 and H3K36me3 and consequently plays a role in transcription inhibition (Dey et al., 2008). This enzyme has been defined as an inducer of proliferation genes rather than pluripotency genes because of its activity on HOX gene promoters and its stimulation of proliferation genes in embryonic stem cells (Dey et al., 2008). More specifically, studies have shown that KDM5B was directly implicated in cell cycle control by acting on the retinoblastoma protein (pRB), one of the main checkpoints in the G1/S transition of the cell cycle (Chicas et al., 2012). Additionally, it has been shown that KDM5B increases the transcription or proliferative genes in embryonic stem cells and contributes to minimizing cryptic transcription. By inhibiting inappropriate initiation of transcription, KDM5B maximizes the total number of correct transcription events and, consequently, upregulates the transcription of target genes (Xie et al., 2011).
A previous study on bovine embryos has shown that the mRNA level of KDM5B was significantly lower in oocytes and pre-MET embryo stages than at the eight-cell stage and post-MET stages (McGraw et al., 2007). However, our immunofluorescence results clearly show a nuclear localization of KDM5B in the oocyte despite the limited transcriptional activity in fully grown oocytes. This presumed function is in agreement with observations in mice, where a high expression level of KDM5B is measured in testis, more specifically in spermatogonia achieving meiosis (Madsen et al., 2003). In fact, when KDM5B is underexpressed, spermatogonias are unable to correctly process meiosis. In the ovary, KDM5B expression is significantly weaker than in spermatogonia, but could still play a significant function, similar to that in spermatogonia (Madsen et al., 2003). Nevertheless, protein abundance from MII to the eight-cell stage was almost null. The numerous functions of the protein also include inhibition of cellular differentiation. Upregulation of the gene following MET as well as nuclear localization of the protein from that stage to blastocyst support this assumption.
Interestingly, in the blastocyst, KDM5B was almost exclusively observed in the inner cell mass, as revealed by immunofluorescence analyses. Knowing that these cells, in contrast to trophectoblast cells, are still pluripotent, the presence of KDM5B corroborates the studies showing the antidifferentiation and antiproliferation effect of that enzyme. In summary, our results suggest that KDM5B predominance in inner cell mass cells would be the reflection of the proliferative and weakly differentiated state of these cells.
In conclusion, analysis of transcript abundance and protein expression of several factors involved in epigenetic remodeling highlights the fact that factors involved in epigenetic remodeling are present at different levels in oocyte and early embryo stages. For many of the genes studied, namely TET3, KDM3A, and KDM4A, important maternal transcript reserves were observed in the oocyte and were translated throughout embryonic development. Except for TET3, whose expression was limited to the oocyte genome, embryonic genome activation refuels transcript abundance of those genes. For KDM4C, maternal accumulation of transcripts was not clearly correlated to protein presence before MET but, similar to other genes studied, embryonic genome activation upregulated its transcription and increased translation. Finally, KDM5B was absent from pre-MET embryo stages but was upregulated in the blastocyst period, around the time that the embryonic pattern of epigenetic modification is implanted. Although further characterization is required, this study has set firm foundations for the involvement and comprehension of specific chromatin remodeling and regulation genes in oocyte and embryo development.
Footnotes
Acknowledgments
This study was supported by the Natural Sciences and Engineering Research Council of Canada (NSERC). The authors acknowledge Isabelle Laflamme for her assistance in the collection of ovarian tissue and Dr. Isabelle Dufort for the revision of the manuscript.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.