
Abstract
Abstract
The repairing function and differentiation potency of adipose stem cells (ASCs) transplantation following skeletal muscle injury induced by radiotherapy are still not well defined. In this study, one side of the buttocks of 64 New Zealand white rabbits underwent irradiation and were randomly divided into an ASCs group [5×107 ASCs labeled with CM-Dil and suspended in 1 mL of phosphate-buffered saline (PBS), via intramuscular injection] and a PBS group (1 mL of PBS, via intramuascular injection). ASCs were isolated in New Zealand white rabbits in vitro, and migration of ASCs labeled with CM-Dil was observed after transplantation in vivo. A significant decrease of histological severity scoring was found in irradiated tissue obtained in the ASCs group compared with that in PBS group. Additionally, compensatory hyperplasia was noted after ASCs transplantation in the injured tissues. Moreover, ASCs could upregulate the expression of vascular endothelial growth factor (VEGF) and basic fibroblast growth factor (bFGF) and promote the angiogenesis of the injured tissues. Interestingly, myofilament-like structures were identified in irradiated muscle cells after ASCs transplantation. We concluded that ASCs transplantation could repair the radiation-induced skeletal muscle injury. Its mechanism may be, at least partly, associated with the upregulation of VEGF and bFGF, angiogenesis, promoting the compensatory hyperplasia of muscle satellite cells, as well as the myogenic differentiation.
Introduction
R
Adipose stem cells (ASCs) are multipotent, differentiating along the adipocyte, chondrocyte, myocyte, and osteoblast lineages (Banas et al., 2007; De Ugarte et al., 2003; Dicker et al., 2005; Gimble and Guilak, 2003; Wagner et al., 2005). Currently, studies have been carried out to investigate the treatment efficiency of ASCs on radiation-induced skin injury, and the treatment efficiencies have been satisfactory to some extent (Akita et al., 2010a, b; Ebrahimian et al., 2009; Hadad et al., 2010; Rigotti et al., 2007). Unfortunately, few studies have been carried out to investigate the treatment efficiency of stem cell therapy on muscular injury induced by irradiation. Agay et al. (2010) reported that angiogenesis was observed around the muscle fibers injured by irradiation during the treatment of cutaneous radiation syndrome with autologous bone marrow mesenchymal stem cells (BMSCs) and ASCs in the minipig.
Blood supply to injury sites is of prime importance for healing, and vascular endothelial growth factor (VEGF) and basic fibroblast growth factor (bFGF) are crucial for the angiogenesis during the injury healing (Flamme et al., 1995; Nissen et al., 1998). To our knowledge, no study has been carried out to investigate the effects of local ASC transplantation on growth factors, and thereby promoting the repair of the radiation-induced skeletal muscle injury. Additionally, there are still disputes about whether ASC transplantation could repair skeletal muscle injury induced by radiotherapy repair or ASCs could transdifferentiate into myocytes. In this study, ASCs were isolated from New Zealand rabbit in vitro. The allogeneic ASCs were transplanted into the rabbits to investigate the mechanisms of how ASCs may function in the angiogenesis and repair of radiation-induced skeletal muscle injury.
Materials and Methods
Healthy New Zealand white rabbits [approval no. SCXK(SU) 2007–2008 and certificate no. 0004282] were purchased from the Qinglongshan Animal Breeding Center (Nanjing, China). The animals were raised in standard cages with a room temperature of 21±1°C, a relative humidity of 55%, and a 12 h/12 h light/dark cycle with food and water. All the experiments were approved by the Ethics and Research Committee of the Affiliated Changzhou No. 2 People's Hospital, Nanjing Medical University. All animal procedures were in accordance with the Principles of Laboratory Animal Care formulated by the National Society for Medical Research and the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health (NIH publication 85-23, revised 1996).
Isolation and culture of ASCs
Adipose tissue was harvested from 26-week-old (weighting 3000±500 grams) New Zealand white rabbits in the subcutaneous fatty tissues of the back region. The tissues were digested with collagenase type I (0.075% wt/vol; Sigma, Santa Clara, CA, USA) at 37°C for 60 min and centrifuged at 201×g for 10 min to obtain high-density stromal vascular pellets. The pellets were resuspended with Dulbecco's Modified Eagle's Medium (DMEM) containing 10% fetal bovine serum (FBS). The cell suspension was cultured in an incubator at 37°C in a humidified atmosphere (100%) containing 5% CO2. After 24 h, the cells were washed using phosphate-buffered saline (PBS) to remove the nonadherent cells and obtain the primary cell (P0). The cells were cultured in the DMEM containing 10% FBS, which was replaced twice every week until 80% confluence was reached. ASCs were observed using an inverted phase-contrast microscope (Olympus, CKX41-A32PH).
Phenotype of cultured ASCs identification
Flow cytometry (Becton-Dickinson, Becton, NJ, USA) was carried out to determine the cell-surface markers of the ASCs. ASCs (P3) were fixed for 30 min in ice-cold 1% paraformaldehyde. Cells were then stained with primary antibodies at room temperature for 30 min in the dark. Because few immunostaining antibodies were available, the following primary antibodies and isotype-matched immunoglobulin G (IgG) were used: Fluorescein isothiocyanate (FITC)-conjugated mouse monoclonal (FITC.MRC OX-7) to CD90/Thy1 (FITC) antibodies and mouse monoclonal (MOPC-21) mouse IgG (FITC) (Abcam, New Territories, HK), rabbit anti-CD 31 FITC-conjugated and rabbit IgG (FITC)-conjugated (Biosynthesis Biotechnology, Beijing), rabbit anti-CD34 RBITC-conjugated, and rabbit IgG [heavy and light chains (H+L)] rhodamine B isothiocyanate (RBITC)-conjugated (Biosynthesis Biotechnology, Beijing). Data analysis was carried out with Cell Quest software (Becton-Dickinson, Becton, NJ, USA).
CM-Dil labeling of ASCs
For the labeling of ASCs, 5×106 ASCs (P3) were inoculated in a 75-mL culture flask. The cells were digested by trypsinase once the exponential growth phase was achieved. After that, 1 mL of PBS was added to the mixture. Subsequently, CM-Dil labeling was carried out with strict adhesion to the manufacturer's instructions (Maibio Biotech, Shanghai, China). In brief, 2.5 μL of CM-Dil (1 μg/μL) was inoculated with the cell suspension at 37°C for 5 min, and finally for 15 min at 4°C. Then the mixture was centrifuged at 201×g for 5 min. The cells were resuspended using PBS to wash the free CM-Dil three times. A portion of the cells labeled with CM-Dil was inoculated in the flask and observed using a fluorescence microscope (Olympus, IX71-22FL/P) with an excitation wavelength of 553 nm.
Protocol I: In vivo study
Animal study
Sixty-four New Zealand white rabbits (8 weeks old, weighing 1500±250 grams) were anesthetized using 1.5% pentobarbital sodium via intraperitoneal injection. The animals were fixed in a prone position. The skin distance focus was 100 cm, and the field size was 5×5 cm2. The dose rate was 300 cGy/min, and the maximum dose depth was 2 cm subcutaneously. Using these parameters, monodose radiation (80 Gy) from a linear accelerator (Primus-Plus, Siemens) producing electron rays of 9 MeV energy was randomly delivered to one side of the buttocks of each animal. The animals were then randomly divided into an ASCs group (n=32, 5×107 allogeneic ASCs labeled with CM-Dil suspended in 1 mL of PBS via local intramuscular injection on the buttock underwent irradiation) and a PBS group (n=32, 1 mL of PBS, via local intramuscular injection on the buttock underwent irradiation). ASCs and PBS were injected 24 h after irradiation. Eight animals were randomly sacrificed using an air embolism through the auricular vein at 1 week, 4 weeks, 8 weeks, and 26 weeks after the treatment. The muscular tissues were obtained from the central area subjected to irradiation (about 2 cm subcutaneously). The body side that had not undergone irradiation was set as the normal control of each group.
Biodistribution of CM-Dil–labeled ASCs in vivo
The distribution of CM-Dil–labeled ASCs was determined at 1 week, 4 weeks, 8 weeks, and 26 weeks after the ASC transplantation. Once the animals were sacrificed, the tissues (0.5 cm3, obtained from the area of ASCs transplantation) were embedded in O.C.T. compound, shock frozen at −80°C, and cut into 4-μm slices, which were frozen. The sections were analyzed using an inverted fluorescence microscope (IX71-22FL/PH, Olympus) to determine the distribution of CM-Dil–labeled ASCs in vivo.
Histopathological studies
The muscle samples (1 cm3) were fixed using 10% formaldehyde. Then each sample was embedded in paraffin, cut into 5-mm sections, and stained with Hematoxylin & Eosin (H&E). All microscope images (BX-4, Olympus) were analyzed with image analyzing software (Langjia Biological Medical Engineering Co., Wuxi, China).
Assessment of the histological severity of lesions
The assessment of the histological severity of lesions was performed on H&E-stained slides, as previously described (Gallet et al., 2011) and centered mainly on the presence of dystrophic nuclei and necrosis. In brief, a score of severity was set as follows: Grade 0, no alteration; grade 1, some dystrophy of nucleus and swelling of muscle cells; grade 2, dystrophy of nucleus and rare aspects of necrosis; and grade 3, dystrophy of nucleus and necrosis. The comparisons among the scores were carried out by two personnel blinded to the conditions.
Quantification of centronucleated regenerating myofibers
Twenty-six weeks after irradiation, the assessment of muscle regeneration from muscle satellite cells in the PBS group and ASCs group was performed on H&E-stained slides. Because regenerating myofibers were sparse in the areas of fibrosis, the areas rich in regenerating myofibers were chosen by scanning the sections at low power (40×and 100×). After the area rich in regenerating myofibers was identified, individual regenerating myofibers were randomly counted on five fields (400×). Any myofiber with a larger centronucleus and increased cytoplasm was considered a single countable regenerating myofiber. Morphometric analyses were carried out by two researchers blinded to the conditions.
Ultrastructural imaging of the muscular tissues
Sections (1 mm3) obtained at 4 weeks and 26 weeks were fixed with 5% glutaraldehyde for 2 h at 4°C. The samples were washed using PBS buffer and fixed using 1% osmic acid for 2 h at 4°C. These samples were then washed and soaked in 2% uranyl acetate solution for 2 h. After dehydration, immersion, and embedding, the sections were prepared with a depth of 50–80 nm and observed using a JEM-1010 transmission electron microscope (JEOL).
Immunohistochemical staining of VEGF, bFGF, and CD31
Immunohistochemical staining was performed to evaluate the expression of VEGF, bFGF, and CD31 in the muscular tissues using an enhanced labeled polymer system. The following primary antibodies were used in immunohistochemistry studies: VEGF antibody (1:300; Abbiotec, San Diego, CA, USA), rabbit anti-bFGF antibody (1:300; PL Laboratories, British Columbia, Canada), and mouse monoclonal anti-CD31 antibody (Abcam, New Territories, HK). Goat anti-rabbit IgG (H+L), FITC-conjugated (PL Laboratories, British Columbia, Canada), was the second antibody.
Quantification of the immunostaining of VEGF and bFGF was performed by digital image analysis with Image-Pro Plus 6.0 software (Media Cybernetics, Silver Spring, MD, USA) according to previous reports (Guo et al., 2013). Because brown-staining myofibers were sparse in fibrosis areas, five microscopic fields (200×) rich in brown-staining myofibers were randomly chosen for the assessment of the integrated optimal density (IOD) on each section. To determine capillary density, as specified by Weidner et al. (1991), any brown-staining endothelial cells or endothelial cell clusters that were clearly separate from adjacent microvessels and other connective tissue elements were considered a single countable microvessel. Three microscopic fields (200×) with the highest neovascularization were selected and microvessels were counted. Morphometric analyses were carried out by two researchers blinded to the conditions.
Western blot analysis
Western blotting analysis was performed routinely, as described (Nie et al., 2012). Briefly, muscular tissues (200 mg) were homogenized in radio immunoprecipitation assay (RIPA) lysis buffer containing protease and phosphatase inhibitors. Proteins were separated by electrophoresis on a 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gel and transferred to a polyvinylidene difluoride (PVDF) membrane. Subsequently, the membranes were blocked with 5% nonfat milk and incubated with primary antibodies against VEGF (1:200; Abbiotec, San Diego, CA, USA), anti-bFGF (1:200; PL Laboratories, British Columbia, Canada) overnight at 4°C, followed by incubation with the horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG secondary antibody (1:5000 Keygen Biotech, Nanjing, China), for 1 h at room temperature. The same membrane was probed for β-actin for loading control. The immune complex was quantified by photodetection. The relative density of VEGF and bFGF to β-actin was analyzed with the Gel-Pro Analyzer 3.2 (Media Cybernetics, Rockville, MD, USA)
Protocol II: In vitro study
Collection of conditioned medium for ASCs
The induction of skeletal muscle injury was performed as mentioned above. One week later, 4.5 grams (wet weight) of muscle tissues obtained from the irradiated and nonirradiated buttock, respectively, were added to 30 mL of DMEM. The tissues were homogenized and centrifuged at 560×g for 5 min. Then the supernatants were filtered using a 0.22-μm syringe filter. The supernatants were then added to four separate wells of a six-well plate as follows: 5 mL of normal muscle supernatant, 5 mL of normal muscle supernatant plus 2×105 cells ASCs (P3), 5 mL of irradiated muscle supernatant, and 5 mL of irradiated muscle supernatant plus 2×105 cells ASCs (P3), respectively. The six-well plates were cultured in a humidified atmosphere (100%) containing 5% CO2 at 37°C. The conditioned medium was collected at 24, 48, 72, and 92 h, respectively.
Determination of VEGF and bFGF in conditioned medium
VEGF and bFGF in conditioned medium were tested using the enzyme-linked immunosorbent assay (ELISA) kits (Hushang Biotech, Shanghai, China). Tests were performed according to the manufacturer's instructions. The expression levels of VEGF and bFGF secreted by ASCs in conditioned medium were described as “ΔVEGF” and “ΔbFGF,” respectively, and were calculated as follows:
Statistical analysis
The data were presented as mean±standard deviation. SPSS 16.0 software was used for the data analysis. Student's t-test was used for the intergroup comparison. The Mann–Whitney test was used for the analysis of each histological severity score. p<0.05 was considered as a significant difference.
Results
Morphology and cell-surface markers of cultured ASCs
The morphology of the cultured ASCs was nearly the same from P0 to P3. Fibroblast-like cells were spindle shaped with clear cellular boundaries. The nucleus was well stacked in the center of each cell. After CM-Dil labeling, the membrane and cytoplasm were labeled in red. No significant change was noted in the morphology of the cells after CM-Dil labeling. The cell-surface marker analysis indicated ASCs represented a homogeneous population of cells with low expression of CD34 and CD31 and high expression of CD90 (Fig. 1A–C).

The cell-surface markers of cultured ASCs. (
Biodistribution of CD-Dil–labeled ASCs in muscle tissues
No fluorescence was noted in the tissues obtained from PBS group. For the ASCs group, migration of ASCs along the needle passage was observed in the tissues with irradiation injury. Gradually, the fluorescence attenuated and the labeled ASCs were integrated into the surrounding muscle cells (Fig. 2a–c).
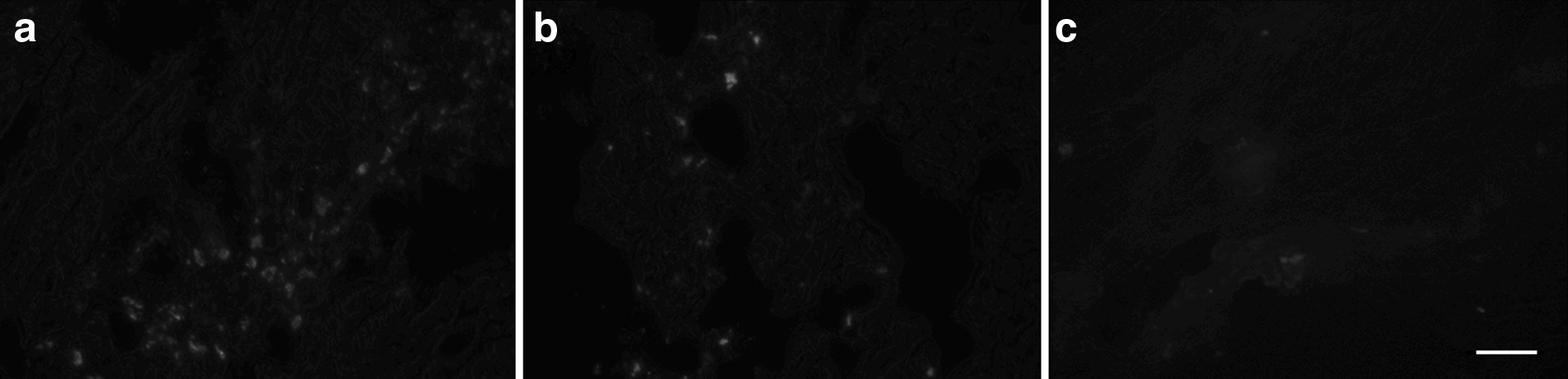
Distribution of CM-Dil–labeled ASCs in the ASCs group after irradiation at a monodose of 80Gy. (
Histopathological results
Observation by light microscopy
After irradiation at a monodose of 80 Gy, the histological severity scores obtained in the ASCs group and the PBS group were higher than those of the normal control group. For the ASCs group, a remarkable decrease of the histological severity scores was noted at 4, 8, and 26 weeks after irradiation compared with those obtained in the PBS group at the same time point, respectively (p<0.05) (Table 1). The number of centronucleated regenerating myofibers in the ASCs group was significantly larger than that in PBS group 26 weeks after irradiation (27.01±9.36 vs. 5.23±4.23, p<0.05) (Fig. 3a–b).
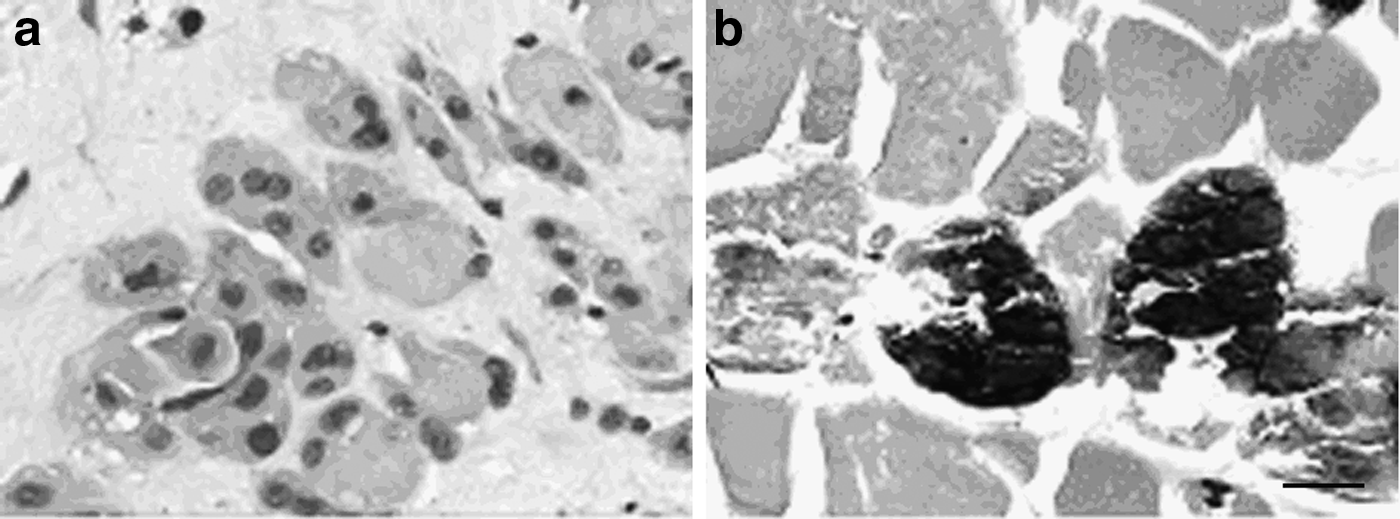
Compensatory hyperplasia of muscle satellite cells at 26 weeks after irradiation at a monodose of 80Gy. (
After irradiation at a monodose of 80 Gy, the histological severity scores in the ASCs group was significantly lower than those in the PBS group at 4, 8, and 26 weeks, respectively.
PBS, phosphate-buffered saline; ASCs, adipose stem cells.
Observation by electron microscopy
Uneven sarcomere swelling was identified in the ASCs group and the PBS group 4 weeks after irradiation at a monodose of 80 Gy. The light band and dark band were not distinct. Multiple focal necrosis was noted in the myofibrils, which formed an amorphous structure of uneven electron density. The size of the nucleus showed a decrease in partial cells together with attenuation of chromatin and margination. Additionally, mitochondrial swelling and rupture and vanishing of the cristae were observed. The density of the mitochondrial matrix was lower, with vacuole-like lesions compared to that of normal mitochondrial matrix. At 26 weeks after the irradiation, the severity of the injury aggregated in the PBS group, as indicated by enhanced necrosis. Nevertheless, for the ASCs group, the lesion showed improvement together with reduced necrotic area. Moreover, differing from normal adult myofilaments with mature sarcomeres (Fig. 4a) and amorphous area of necrosis sarcomeres (Fig. 4b), short and clustered myofilament-like structures in irregular alignments were observed in the sarcomere (Fig. 4c), the spatium intermusculare (Fig. 4d), and the subsarolemma (Fig. 4e–f′). These structures showed obviously higher density than those of inherent ultrastructures of muscle cells. Because these structures were not noted in the PBS group, we speculated that ASCs may be involved in myogenic differentiation.

The ultrastructures of the skeletal muscular tissues were observed under a transmission electron microscope. (
Effects of ASCs implantation on the expression of VEGF, bFGF, and CD31
VEGF and bFGF were stained brown and distributed in the cytoplasm of the muscle cells, as indicated by the immunohistochemisty analysis of skeletal muscle tissues. There was no statistical difference at each time point (p>0.05) in the expression levels of VEGF and bFGF in the normal control group. Nevertheless, the expression levels of VEGF and bFGF in the ASCs group were all significantly higher than those in the PBS group and normal group at 1, 4, 8, and 26 weeks (p<0.05; Fig. 5A–C). After irradiation, the density of microvessels positively stained with CD31 showed a remarkable increase after ASCs implantation at 4, 8, and 26 weeks compared with that of the PBS group (p<0.05; Fig. 5A,D). Moreover, the expression levels of VEGF and bFGF proteins detected using western blotting showed an increase at each time point in the ASCs groups compared with those of the PBS group (Fig. 5E–F).

The changes of growth factors and angiogenesis treated with ASCs or PBS in irradiated skeletal muscular tissues. (
Growth factors released by ASCs
No statistical difference was noted in the expression levels of VEGF and bFGF in normal muscle supernatant, irradiated muscle supernatant, and conditioned medium obtained from the normal skeletal muscle with cultured ASCs at 24, 48, 72 h, and 96 h, respectively (p>0.05). The expression levels of VEGF obtained from the irradiated skeletal muscle supernatant with cultured ASCs were higher than those in irradiated muscle supernatant at 48 h, 72 h, and 96 h (p<0.05). The expression levels of VEGF secreted by ASCs in irradiated muscle supernatant (ΔVEGFirradiated muscle) were higher than those secreted by ASCs in normal muscle supernatant (ΔVEGFnormal muscle) at 48 h, 72 h, and 96 h, respectively (p<0.05; Fig. 6A). The expression levels of bFGF obtained from the irradiated skeletal muscle supernatant with cultured ASCs were higher than those in irradiated muscle supernatant at 72 h and 96 h, respectively (p<0.05). The expression levels of bFGF secreted by ASCs in irradiated muscle supernatant (ΔbFGFirradiated muscle) were higher than those secreted by ASCs in normal muscle supernatant (ΔbFGFnormal muscle) at 72 h and 96 h, respectively (p<0.05; Fig. 6B).
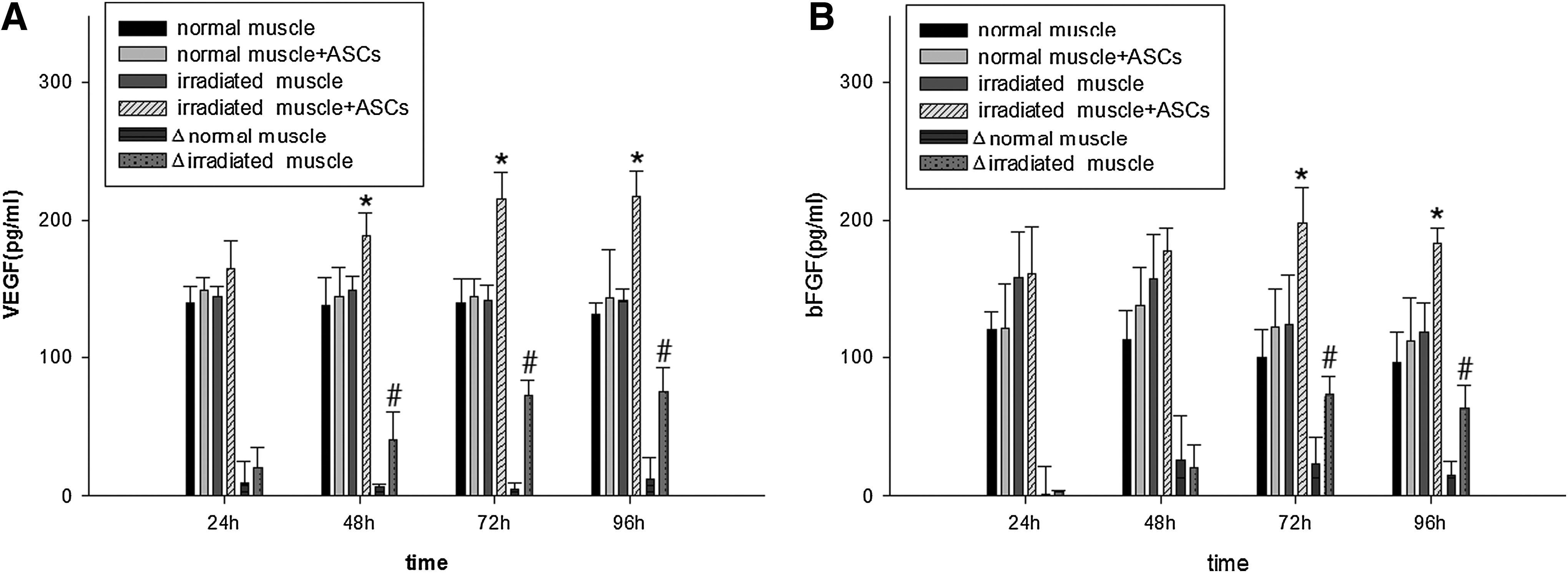
Growth factors released by ASCs. (
Discussion
The maximum tolerated dose of adult skeletal muscle to irradiation is 80 Gy (TD50/5); however, the irradiation dose for inducing skeletal muscle injury in animal models is still disputed. In a previous study, cobalt-60 irradiation of rats was carried out on the hind limb at a monodose of 30 Gy. The results indicated that irradiation induced a marked increase of histological scoring compared to that of unirradiated skeletal muscles 6 months after irradiation (Gallet et al., 2011). Rabbits exposed to single dose of iridium-192 gamma irradiation (120 Gy) on the iliospinal muscle showed muscular necrosis at 5 weeks postexposure (Alberini et al., 2000). A single dose of gamma rays (160 Gy) at the skin surface of pig models could induce femoris muscle radiation fibrosis at 5–6 months postexposure (Geffrotin et al., 1998). In a study carried out by Powers and colleagues (Powers et al., 1991), intraoperative irradiation using 6 MeV electrons combined with external beam irradiation using a 6 MV linear accelerator (80 Gy) could lead to radiation-induced muscle injury in beagle dogs, which was characterized by loss of muscle fibers, decreased fiber size, severe vascular lesions, hemorrhage, inflammation, coagulation necrosis, and fibrosis. On this basis, the authors speculated that the radiation-induced muscle injury was likely to be associated with the injury of supporting vasculature. Hsu et al. (1998) reported that male Wistar rats were exposed to 80 Gy 6 MV X rays from a linear accelerator given in 40 fractions of 2 Gy; the irradiated gastrocnemius muscle displayed injury of vasculature, as well as injury of muscle cells at 6 months after irradiation. Currently, the skeletal muscle injury in clinical practice was mainly caused by a high dose of irradiation. Therefore, a high dose (80 Gy) was selected in our study. Although monodose irradiation was carried out, similar histopathological changes were noticed compared with those obtained in humans who underwent radiotherapy; cellular swelling, multiple focal necrosis, fibrosis, and inflammatory cell infiltration appeared in the muscular tissues in the PBS group. In the chronic phase, thickening of vessel walls, stenosis, and occlusion of lumina as well as fibroplasis were observed. In the electron microscope imaging, shortened sarcomeres and rupture and vanishing of myofilaments were noted together with a large-scale amorphous area in the PBS group. Moreover, attenuation and margination of chromatin were noted in muscle cells together with mitochondrial paraplasm. These indicated that a monodose of 80 Gy electron rays could be efficient for the establishing of animal models with skeletal muscle injury.
Recently, stem cell therapy has been considered as a promising therapeutical approach for improving wound repair of severe radiation burns (Bey et al., 2010). ASCs, a population of stem cells derived from adipose tissues (Rodriguez et al., 2005), have been considered as the most widely distributed adult stem cells (Strem et al., 2005). In addition, ASCs can be easily obtained from patients by a simple minimally invasive method and expanded to the quantity needed for transplantation in vitro due to their proliferation capacity (Nakagami et al., 2005). Moreover, these cells could maintain multidirectional differentiation potential after successive cultures (Zuk et al., 2002). Therefore, ASCs have been considered as a novel source for cells to be used for the repair of wound injury.
In this study, a significant decrease of histological severity scoring was found in irradiated tissue obtained in the ASCs group compared with that in PBS group. This indicated that allogeneic ASCs transplantation following skeletal muscle injury induced by radiotherapy could repair injured myocytes. However, it is evident to us that histological analysis was not sufficient for precisely evaluating the repair and regeneration of muscle cells. In this context, the analysis of growth factors and observation of ultrastructure appeared to be attractive methods for evaluating tissue changes after ASCs transplantation.
ASCs have the capacity to secrete growth factors, especially the factors associated with angiogenesis (Kinnaird et al., 2004; Rehman et al., 2004). Previous studies indicated that ASCs could upregulate VEGF and bFGF in response to hypoxia (Chung et al., 2009; Lee et al., 2009). Similarly, our results demonstrated that ASCs could increase expression of VEGF and bFGF in the conditioned medium, mimicking the microenvironment of radiation-induced skeletal muscle injury. It has been well acknowledged that ASCs could alternate the expression of growth factors in the lesion area after transplantation and on this basis repair the wound injury accordingly in vivo.
Previous studies have shown that ASCs could enhance angiogenesis in animal models with nonirradiated skin injury (Lee et al., 2009), skeletal muscle injury (Bhang et al., 2009; Nakagami et al., 2005), myocardium injury after myocardial infarction (Miyahara et al., 2006), as well as brain injury after focal cerebral ischemia (Wang et al., 2008) through upregulation of VEGF and bFGF. Nevertheless, the mechanism of how ASCs function in the wound injury (i.e., through autologous differentiation or paracrine section) is still unclear (Kinnaird et al., 2004; Lee et al., 2009; Matsuda et al., 2013).
How ASCs function in irradiated skeletal muscle injury also remains unknown. Agay and his colleagues (Agay et al., 2010) developed a new minipig model of cutaneous radiation syndrome (CRS) to investigate the potential therapeutical benefit of BMSCs and ASCs. The results indicated that BMSCs and ASCs led to improved vascularization in injured muscle. Phulpin et al. (2009) reported that a decrease in irradiated morphologic patterns characterized by an absence of necrotic areas and a vascular network with high-density was noted after transplantation of autologous fat tissues in patients with irradiated head and neck tissues. They speculated that ASCs may be involved in angiogenesis.
In our study, the allogeneic ASCs were labeled with CM-Dil and then injected into irradiated muscular tissues. The biodistribution analysis indicated that the allogeneic ASCs labeled with CM-Dil could migrate in the injured tissues and integrate into the surrounded muscle cells. Additionally, upregulation of VEGF and bFGF was noted in the injured tissues in the ASCs group. Moreover, the density of microvessels positively stained with CD31 showed a remarkable increase after ASCs implantation. Collectively, we speculated that ASCs transplantation may increase angiogenesis in the injured tissues, at least partly, by upregulation of VEGF and bFGF.
In cases of muscle cell injury, the satellite cells were activated and differentiated to repair the injured muscle fibers or regenerate the muscle fibers (Schmalbruch and Lewis, 2000). In our study, significant compensatory hyperplasia of satellite cells was noted in the injured area at 26 weeks after ASCs transplantation compared with that in PBS group. This may lead to the conclusion that ASCs may be involved in the repair of radiation-induced muscle injury through promoting the compensatory hyperplasia of satellite cells.
Because different staining methods were used in our study compared with the study carried by Misao et al. (2006), no stacked Dil granules stained in a dark color were observed in the irradiated area in rabbits. However, in our electron microscope study, at 26 weeks after ASCs transplantation, myofilament-like structures with higher density than that of the inherent structures were identified in irradiated skeletal muscle cells. These myofilament-like structures appeared in the ASCs group and were not noted in PBS group, indicating that ASCs may be involved in myogenic differentiation.
Conclusion
ASCs transplantation could repair the radiation-induced skeletal muscle injury. Its mechanism may be associated with the up-regulation of VEGF and bFGF, angiogenesis, repairing of muscle satellite cells, or promoting the compensatory hyperplasia, as well as myogenic differentiation.
Footnotes
Acknowledgments
This project was sponsored by the Changzhou Municipal Health Bureau Instructional Science and Technology Plan (no. WZ201018), Changzhou Municipal “831” Scientific Research Program, Changzhou Municipal Medical Innovation Talent Program (no. 2–14), and Jiangsu Provincial“333 Talent” Program.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.