
Abstract
Abstract
On the basis of their self-renewal capacity and their ability to differentiate into derivatives of all three germ layers, germ line–derived multipotent adult stem cells (maGSCs) from mouse testis might serve as one of preferable sources for pluripotent stem cells in regenerative medicine. In our study, we aimed for an efficient hepatic differentiation protocol that is applicable for both maGSCs and embryonic stem cells (ESCs). We attempted to accomplish this goal by using a new established co-culture system with OP9 stroma cells for direct differentiation of maGSCs and ESCs into hepatic cells. We found that the hepatic differentiation of maGSCs was induced by the OP9 co-culture system in comparison to the gelatin culture. Furthermore, we showed that the combination of OP9 co-culture with activin A resulted in the increased expression of endodermal and early hepatic markers Gata4, Sox17, Foxa2, Hnf4, Afp, and Ttr compared to differentiated cells on gelatin or on OP9 alone. Moreover, the hepatic progenitors were capable of differentiating further into mature hepatic cells, demonstrated by the expression of liver-specific markers Aat, Alb, Tdo2, Krt18, Krt8, Krt19, Cps1, Sek, Cyp7a1, Otc, and Pah. A high percentage of maGSC-derived hepatic progenitors (51% AFP- and 61% DLK1-positive) and mature hepatic-like cells (26% ALB-positive) were achieved using this OP9 co-culture system. These generated hepatic cells successfully demonstrated in vitro functions associated with mature hepatocytes, including albumin and urea secretion, glycogen storage, and uptake of low-density lipoprotein. The established co-culture system for maGSCs into functional hepatic cells might serve as a suitable model to delineate the differentiation process for the generation of high numbers of mature hepatocytes in humans without genetic manipulations and make germ line–derived stem cells a potential autologous and alternative cell source for hepatic transplants in metabolic liver disorders.
Introduction
S
Adult stem cells are found in nearly all postnatal organs and tissues and have the capacity for renewal after disease. Adult stem cells overcome the limits regarding ethical and immunocompatible problems, but their differentiation potential is limited. Nevertheless, some adult stem cells have already shown their ability for hepatocyte differentiation, including hematopoietic stem cells (Alison et al., 2000; Petersen et al., 1999) and mesenchymal stem cells from bone marrow (Sato et al., 2005; Schwartz et al., 2002), umbilical cord blood (Lee et al., 2004; Wang et al., 2005), or placenta (Chien et al., 2006).
In the past years, pluripotent germ cells became much more interesting because of their developmental potential. Spermatogonial stem cells (SSCs) were shown to be isolated from both human and murine testis (Conrad et al., 2008; Dym et al., 2009; Golestaneh et al., 2009; Kossack et al., 2009; Seandel et al., 2007). We showed that multipotent adult germ-line stem cells (maGSCs) could be established from isolated SSCs from adult mouse testis. They revealed ESC properties in vitro and were able to differentiate spontaneously into various cell types of all three germ layers, including the endodermal epithelium and hepatic-like cells (Guan et al., 2006). Furthermore, maGSCs can differentiate into functional cardiomyocytes, neuronal cells, and endothelial cells (Cheng et al., 2012; Guan et al., 2007; Streckfuss-Bömeke et al., 2009). maGSCs have the common features of both ESCs and adult stem cells and are therefore of prime importance in tissue regeneration. And most importantly, no genetic manipulations are required for reprogramming these adult stem cells into pluripotent cells.
Two groups reported the generation of functional hepatocytes from germ line cell–derived pluripotent stem cells in vitro (Fagoonee et al., 2010; Loya et al., 2009). However, in these studies, the differentiation of pluripotent stem cells toward the hepatic phenotype was achieved spontaneously by embryoid bodies, resulting in a low efficiency of spontaneous endodermal specification and differentiated hepatic progenitors.
One important step in the establishment of the hepatic lineage in differentiation cultures is the recapitulation of the signaling pathways of the early embryo for endoderm induction and specification to the liver. Previous studies reported that activin A (AA) efficiently induced pluripotent cells to form definitive endoderm in vitro, mimicking the role of Nodal, a related factor that signals through the same receptor (Hay et al., 2008; Kubo et al., 2004; Tam et al., 2003). Liver specification in the mouse embryo is controlled by inductive signals from subpopulations of mesodermal cells (Lemaigre and Zaret, 2004). It is known that signals from the cardiac mesoderm (Jung et al., 1999) and from the septum transversum mesenchyme (Rossi et al., 2001) initiate liver bud formation in the anterior definitive endoderm. The OP9 co-culture was shown to induce the mesodermal lineage in ESCs and maGSCs (Cheng et al., 2012; Nakano et al., 1994). In addition, OP9 cells secrete high amounts of stem cell factor (SCF), which is important in liver and myocardial regeneration (Meng et al., 2012). OP9 cells do not express functional macrophage colony-stimulating factor (M-CSF). The presence of M-CSF had inhibitory effects on the differentiation of ESCs to blood cells other than macrophages (Nakano et al., 1994). We hypothesized that the OP9-induced mesodermal formation of maGSCs is required for specification of the endoderm to hepatic cells and enhances the amount of differentiated hepatic progenitors and mature hepatocytes.
In this study, we show for the first time the successful establishment of a co-culture differentiation system of maGSCs with OP9 cells to generate high numbers of hepatic progenitors and mature hepatocytes in vitro. These differentiated cells are characterized by an expression pattern of genes and proteins specific for endodermal-derived hepatic progenitors and mature hepatocytes that are functional in vitro.
Materials and Methods
Cell culture of maGSCs
Mouse maGSCs used in this study were derived from SV129/Ola mice as described previously (Guan et al., 2006). Expansion of undifferentiated maGSCs was carried out by cultivation on mitomycin C–inactivated mouse embryonic fibroblasts (MEFs) isolated from 13.5- to 16.5-day-old embryos (Wobus et al., 2002) in ESC medium: Dulbecco's modified Eagle medium (DMEM; Invitrogen, Darmstadt, Germany) supplemented with 15% fetal calf serum (FCS; selected batch, Invitrogen), 1×minimum essential medium (MEM)-nonessential amino acids (NEAA, Invitrogen), 50 μM β-mercaptoethanol (β-ME; Promega, Mannheim, Germany), and 103 units/mL leukemia inhibitory factor (LIF; ESGRO, Millipore, Schwalbach, Germany).
Cell culture of OP9 cells
OP9 stromal cells were cultured in a MEM α modification (α-MEM; Invitrogen) supplemented with 20% FCS (Lonza, Verviers, Belgium), 1×NEAA, and 50 μM β-ME.
In vitro differentiation of maGSCs
Prior to endodermal differentiation of maGSCs, MEFs were eliminated by using the preplating method. Briefly, cells were trypsinized with 0.1% trypsin and 0.01% EDTA in phosphate-buffered saline (PBS), replated on gelatin-coated (0.1%; Fluka, Taufkirchen, Germany) dishes in culture medium, and incubated at 37°C and 5% CO2. One hour after incubation, MEFs attached the culture dishes, and the floating maGSCs were then collected and used for differentiation.
For spontaneous differentiation, maGSCs (400 cells/20 μL) were used to form embryoid bodies (EBs) using the hanging drop method for 3 days, followed by suspension culture for 2 days. Subsequently the EBs were plated on gelatin-coated dishes (partly containing four cover slips for immunofluorescence staining) in Iscove's modified Dulbecco's medium (IMDM; Invitrogen) supplemented with 20% FCS,
For directed differentiation into the endodermal lineage, maGSCs (typically 3×104 cells/dish) were allowed to grow on mitomycin C–inactivated OP9 stromal cells (typically 1.5×106 cells/dish) in 10-cm cell culture dishes (Nunc, Langenselbold, Germany) in IMDM with supplements. For induction of directed endodermal differentiation, human AA (100 ng/mL; PeproTech, USA) was added for the first 2 days. After 5 days of differentiation, OP9 cells were eliminated, and predifferentiated maGSCs (typically 3×104 cells/dish) were transferred onto fresh OP9 cells.
Differentiation of hepatic progenitors and mature hepatocytes was achieved by culturing on OP9 cells for 5+25 days. The medium was changed every 2 days. For late hepatic differentiation, the following growth factors were added: Basic fibroblast growth factor (bFGF; 100 ng/mL, PeproTech) from day 2 to day 5 and hepatocyte growth factor (HGF; 50 ng/mL, PeproTech) from day 5+5 to day 5+15. Thereafter, the IMDM was changed to hepatocyte growth medium (HGM) with supplements (PromoCell, Germany) to the end of the experiment.
Characterization of generated hepatic progenitors and mature hepatocytes
For characterization of the generated hepatic progenitors and mature hepatocytes, RT-PCR analysis, immunocytochemical staining, flow cytometry, and functional assays [albumin secretion, urea secretion, Periodic Acid Schiff (PAS) staining for glycogen, Indocyanine Green (ICG) uptake, and release and uptake of low-density protein] were performed at various differentiation stages. The detailed procedures are described in Supplementary Data (Supplementary Data are available at www.liebertpub.com/cell/).
Fetal hepatoblast and adult hepatocyte preparation and culture
For fetal hepatoblast isolation, day-14 fetal livers were dissected from timed pregnant NMRI mice, and cells were isolated as previously described (Gouon-Evans et al., 2006). Adult hepatocytes were harvested from the liver of 11- to 12-week-old C57BL/6 mice using a two-step collagenase perfusion protocol. The detailed procedure is included in Supplementary Data.
Animal care
NMRI and C57BL/6 mice were maintained under conventional conditions in the local animal facility. All animal studies (T-30.08; T-28.08) were approved by the local animal care committee at the University Clinic in Göttingen.
Results
maGSCs differentiate spontaneously into the endodermal and hepatic lineages
To analyze the endodermal differentiation potential of maGSCs, we induced these cells to differentiate spontaneously into the hepatic lineage by applying the hanging drop method. The expression of the early endodermal markers Foxa2 [hepatocyte nuclear factor 3β (Hnf3β) (Sund et al., 2000)] and SRY-sex determining region Y-17 [(Sox17) (Sinner et al., 2004)] was increased from day 5 after the initiation of differentiation compared to undifferentiated maGSCs (day 0) (Fig. 1A). The earliest markers of hepatic differentiation, α-fetoprotein (Afp) and transthyretin (Ttr), were expressed at day 5+2. Expression of the late-stage hepatocyte-specific marker α-anti-trypsin (Aat) was observed from day 5+5, whereas albumin (Alb) was expressed later on during differentiation with a peak at day 5+14 (Fig. 1A). These mRNA data were supported by immunofluorescence staining and flow cytometry using antibodies against AFP and ALB. The number of differentiated cells positive for AFP increased from 5.5% at day 5+8 to 27% at day 5+10, measured by flow cytometry. Immunofluorescence staining confirmed the cytoplasmic localization of AFP (Fig. 1B). In contrast, only a few cells showed a positive staining for ALB at day 5+14 (Fig. 1B), which was confirmed by flow cytometry, with 12.7% of the cells positive for ALB at day 5+25. These data indicate that maGSCs can be spontaneously induced to differentiate into the endodermal lineage.
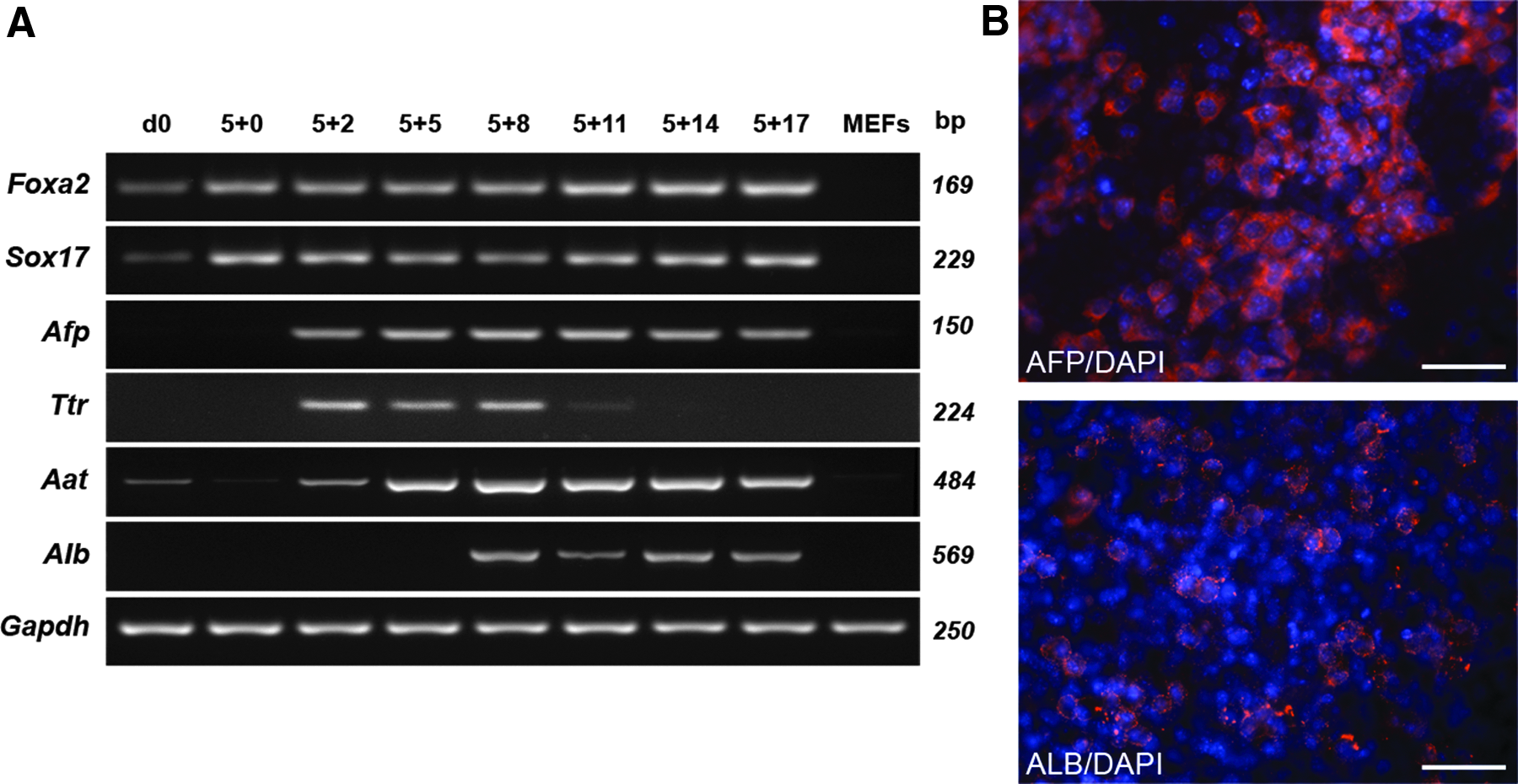
Spontaneous endodermal differentiation of maGSCs using EB formation. (
Standardized protocol for efficient differentiation of maGSCs into hepatic lineages using OP9 co-culture
To enhance the amount of hepatic progenitors and mature hepatocytes, we established a co-culture differentiation protocol of maGSCs with OP9 stromal cells. MaGSCs co-cultured with OP9 stromal cells were directly compared to a monolayer culture on gelatin for endodermal differentiation (Fig. 2). MEF-eliminated undifferentiated maGSCs were cultured on OP9 cells or gelatin for 5 days in differentiation medium. These predifferentiated maGSCs were passaged at day 5 and seeded back on fresh OP9 cells or on gelatin, respectively, for an additional 25-day differentiation (Fig. 2A).

Differentiation of maGSCs into hepatic-like cells. (
RT-PCR analysis of markers and transcription factors specific for the endoderm and hepatic progenitors showed a significantly increased expression of the endodermal gene Sox17 in maGSCs co-cultured with OP9 cells in comparison with cells differentiated on gelatin (Figs. 2B and 3). Furthermore, the early hepatic progenitor markers Hnf4, Afp, and Ttr were also enhanced in co-culture than on gelatin. Thus, co-cultivation of maGSCs with OP9 cells resulted in the efficient induction of hepatic progenitors in contrast to the differentiation on gelatin (Figs. 2B and 3).
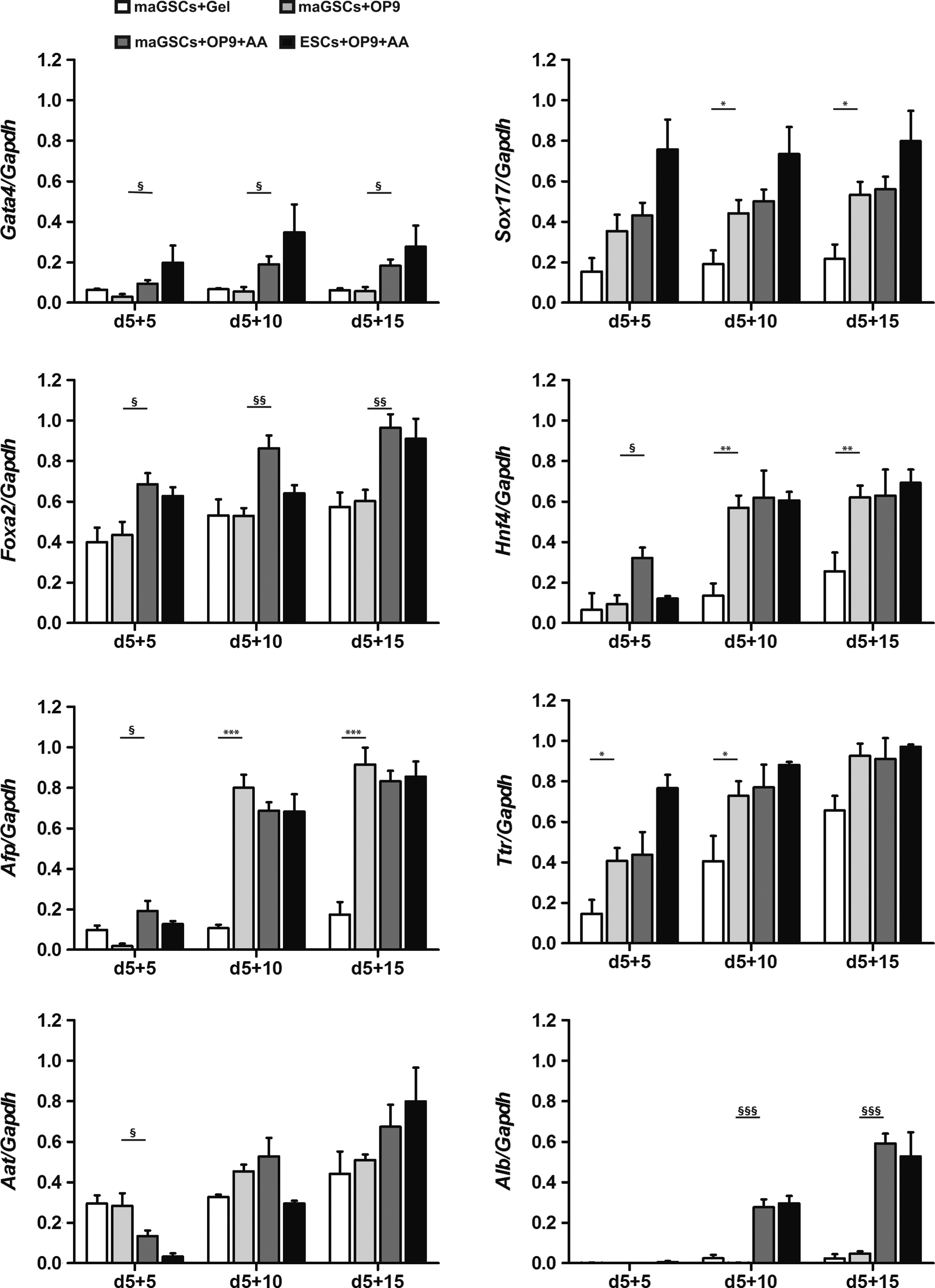
Comparison of expression of hepatic-specific genes during differentiation of maGSCs and ESCs. Semiquantitative RT-PCR was performed to analyze hepatic gene expression profiles of differentiated maGSCs on gelatin (white bars) or on OP9 cells (light grey bars) and with AA treatment (dark grey bars) for various endodermal and hepatic genes. Cells indicated with black bars resemble ESC-derivatives with AA treatment. An increase of expression of the endodermal markers Foxa2 and Gata4 and of the hepatoblast markers Hnf4 and Afp were observed after AA treatment at early time points (Hnf4 and Afp) or all time points of differentiation. At day 5+10 most prominently, an increase of mature hepatic markers like Alb was detected by AA treatment. AA-treated and OP9 co-cultured ESC-derivatives show a similar endodermal and hepatic-like expression pattern as maGSC-derivatives. The graphs depict the means of at least three independent experiments±standard error of the mean (SEM). (* or §) p<0.05; (** or §§) p<0.01; (*** or §§§) p<0.001.
AA enhances the amount of maGSC-derived hepatic progenitors and mature hepatic-like cells
To analyze the influence of AA on the endodermal differentiation capacity of maGSCs in the OP9 co-culture system, high concentrations of AA (100 ng/mL) were added for the first 2 days of in vitro differentiation (day 0 to day 2) in the maGSC-OP9 co-culture (Fig. 2A). When maGSCs were cultivated on OP9 cells and treated with AA, their morphology changed from compact colonies to outgrowing cells at day 5 and sharped-edged, elongated cells at day 5+15 and resulted in a homogeneous cell population with polygonal shape at day 5+25 (Fig. 2A). Semiquantitative analysis of endodermal and hepatic markers revealed a significantly enhanced expression of Gata4 and Foxa2 by the AA treatment at all analyzed time points during maGSC differentiation (Fig. 3). The hepatocyte progenitor markers Hnf4 and Afp were expressed significantly higher in maGSCs treated with AA at day 5+5 compared to the differentiation culture without AA (Fig. 3). Furthermore, a significant increase in expression of the mature hepatic gene Alb was detected by AA treatment at day 5+10 and day 5+15 (Fig. 3). These data indicate that the AA treatment from day 0 to day 2 enhances the endodermal, hepatic progenitor, and mature hepatocyte differentiation of maGSCs using the OP9 co-culture system. To investigate whether the hepatic induction by the AA treatment from day 0 to day 2 (treatment once) can be further enhanced, we applied an addition of AA to the co-culture from day 5 to day 5+2 (treatment twice) and found no major difference in cell morphology and in hepatic gene expression between once and twice AA treatments (Fig. S1) (Supplementary Data are available at www.liebertpub.com/cell/).
To confirm the gene expression data, we further analyzed the expression of SOX17, AFP, DLK1 (hepatic stellate cell-derived delta-like homolog 1, a member of the epidermal growth factor-like family; Laborda, 2000), AAT, ALB, and CK18 (cytokeratin 18) in AA-treated maGSC-OP9 co-culture at protein levels by immunofluorescence staining and flow cytometry (Fig. 4). As shown in Figure 4, most of the day 5+5 differentiated cells were positive for SOX17 with a clear cytoplasmic localization. This confirms that the majority of the cells at this stage were differentiated into definitive endoderm, which occurred before hepatic lineage commitment. A clear cytoplasmic localization of AFP was observed at day 5+10 and a cell-surface staining of DLK1 at day 5+15, indicating the differentiation of maGSCs toward the hepatic progenitors (Fig. 4A). A significant increase was detected in AFP-positive cells from 11% at day 5+5 up to 51% at day 5+15. Furthermore, 35% and 61% of hepatic progenitors at day 5+10 and day 5+15 were positively stained for DLK1, respectively (Fig. 4B). These experiments confirm the effective generation of hepatic progenitors from definitive endoderm. We also observed a significant increase of ALB-positive cells from 13% at day 5+10 to 24% at day 5+15 of differentiation. This observation was confirmed by a cytoplasmic localization of ALB at the same time point. Furthermore, expression of the mature hepatic marker AAT and the intermediate filament protein CK18 were confirmed by immunofluorescence at day 5+10 and day 5+15 (Fig. 4). We further studied expression of various late hepatic markers in maGSC-derived mature hepatic-like cells at day 5+15 compared to cells isolated from adult mouse liver (Fig. S1, Fig. 5). The expression level of Aat and Alb in differentiated cells at day 5+15 was similar to cells isolated from adult mouse liver (Fig. S1). Cytokeratin 18 (Krt18) and 8 (Krt8) were strongly expressed at day 5+15 at a higher level than in adult hepatocytes. Furthermore, the bile duct epithelial marker Krt19 was expressed at differentiation day 5+15, but not in adult hepatocytes (Fig. 5). The expression of markers specific for late mature hepatocytes, such as carbamyl-phosphate-synthase-1 (Cps1), tryptophan-2,3-dioxygenase (Tdo2), SAPK/ERK-kinase-1 (Sek), and cytochrome-P450-dependent monooxygenase 7A1 (Cyp7a1) were detected at day 5+15, but at lower levels compared to adult hepatocytes (Fig. 5). Both ornithine transcarbamylase (Otc), which is involved in the urea cycle (Murakami et al., 1990), and phenylalanine hydroxylase (Pah), which is mainly expressed in hepatocytes (McGee et al., 1972), were expressed at day 5+15, but at a lower level than in adult hepatocytes (Fig. 5). Furthermore, we did not see any difference in the mature hepatic gene expression pattern between one and two times AA treatment (Fig. S1). These data demonstrate that maGSC-derived hepatic-like cells express mature hepatic markers, but at a lower level than in the adult liver cells. The prolongation of the differentiation time period did not lead to further maturation of maGSC-derived hepatic-like cells. We found no significant increase in ALB-positive cells (26%) at day 5+25 (Fig. 4). At late differentiation stage day 5+25, about 35% and 60% of the cells still expressed the hepatic progenitor markers AFP and DLK1, respectively, indicating that differentiation to the mature hepatocyte phenotype was not uniform and not accomplished.
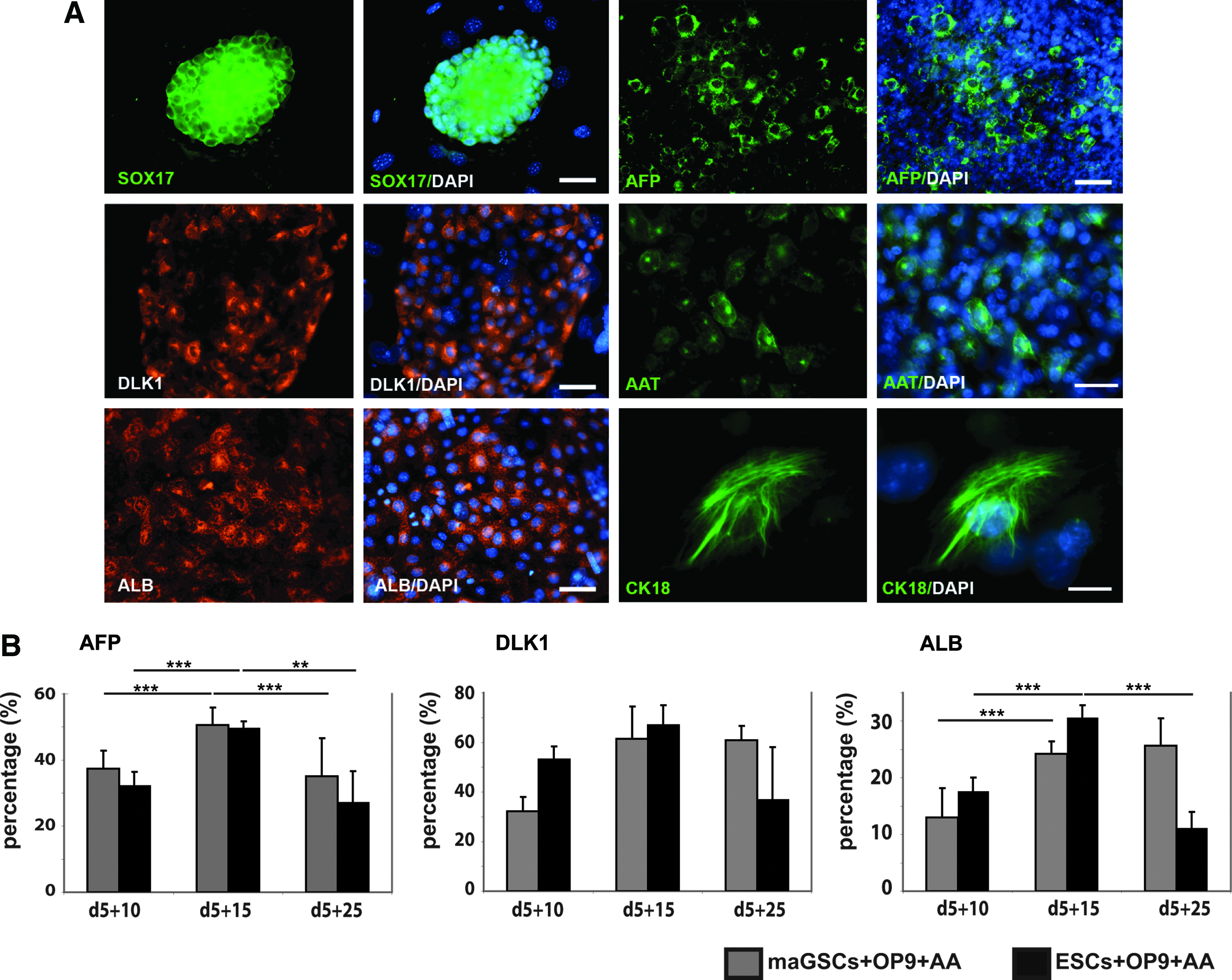
(

mRNA expression of late hepatic markers in maGSCs after co-culture with OP9 and AA at time point day 0 (undifferentiated maGSCs) and day 5+15. Visible expression of late hepatocyte markers Cps1, Tdo, Cyp7a1, Otc, Pah, Sek, Krt18, and Krt8. Krt19 was used as a marker for bile duct cells. Adult liver (AL) was used as positive control.
MaGSC-derived hepatic-like cells are functional in vitro
To test the in vitro functionality of maGSC-derived hepatocytes, we first measured the ALB-secreting potential of the differentiated cells using an enzyme-linked immunosorbent assay (ELISA). The maGSC-derived hepatic-like cell supernatants showed albumin secretion from day 5+10 (12.5 ng/mL) to day 5+25 (25 ng/mL), consistent with the flow cytometry data showing the ALB-positive cells from day 5+15 (13%) to day 5+25 (26%) (Fig. 6). Furthermore, a significant increase of albumin secretion was observed by the AA treatment at day 5+15 (Fig. 6). In addition, urea secretion was measured and resulted in a significant increase by the AA treatment at day 5+15 of differentiation (Fig. 6).
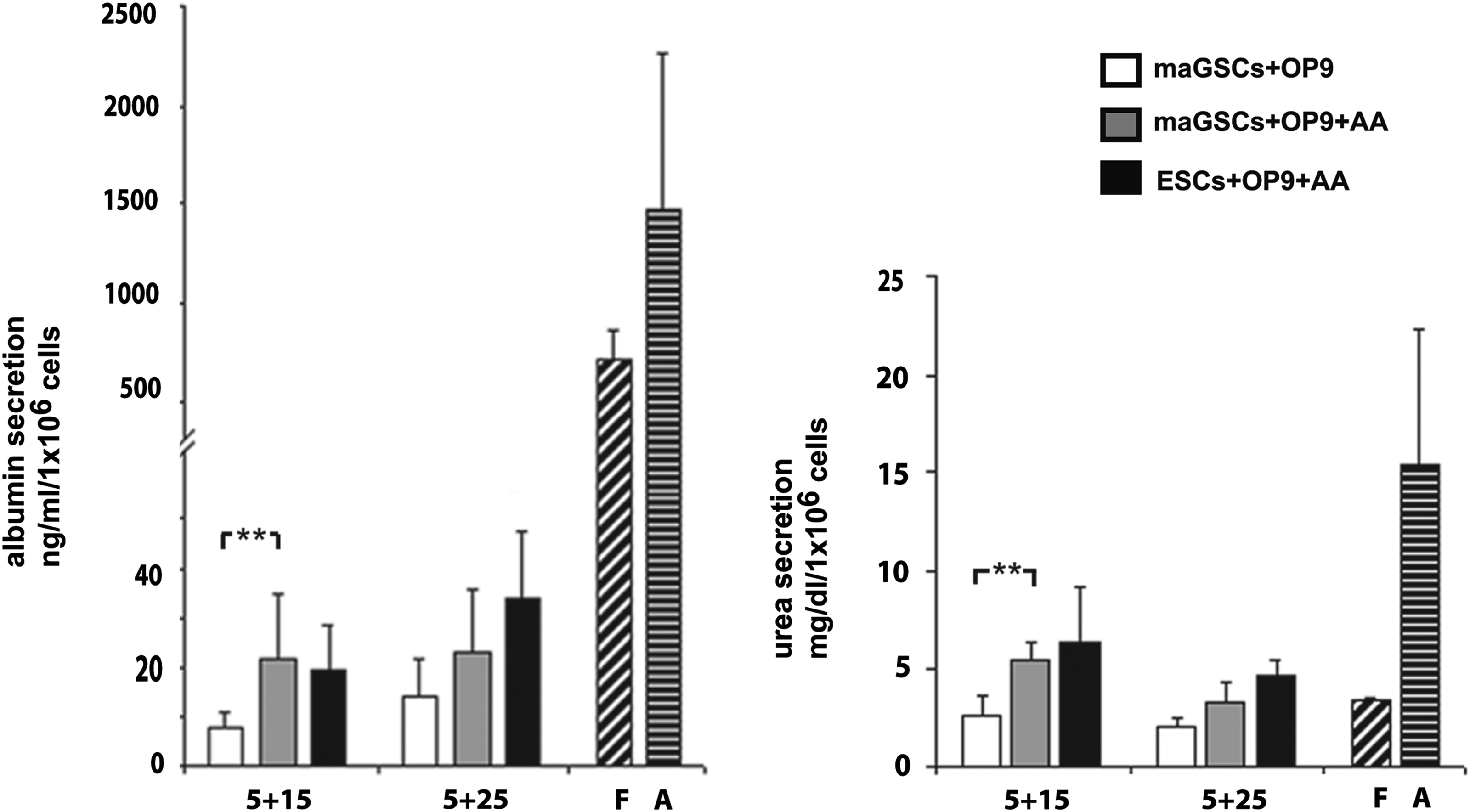
Functional characterization of maGSC- and ESC-derived hepatocytes. Detection of albumin and urea secretion measured by ELISA of 2-day supernatants of maGSC-derived (white, without AA; grey, with AA) and ESC-derived hepatocytes (black, with AA) during differentiation. The levels of albumin and urea secretion were enhanced by AA treatment at day 5+15. Albumin levels are indicated in ng/mL per 1×106 cells and urea secretion is indicated as mg/dL per 1×106 cells. Two-day supernatants from fetal (F) and adult (A) hepatocytes (diluted 1:10) were used as positive controls for the ELISA evaluation. The graph depicts the means of three independent experiments±standard error of the mean (SEM). (**) p<0.01.
Additional assays were performed to characterize endoderm-derived hepatic cell function in vitro (Fig. 7A–D). PAS staining of the maGSC-derived hepatocytes was performed at day 5+5, day 5+15, and day 5+25 to look at typical red cytoplasmic staining indicative of glycogen storage, a characteristic of mature hepatocytes (Lillie et al., 1976). At day 5+25, the majority of the cells showed strong red glycogen staining, in contrast to early differentiated cells at day 5+5 (Fig. 7A). The test for uptake, conjugation, and elimination of ICG is used in the clinic to determine the function of mature hepatocytes and also in ESC-derived hepatocyte cultures to identify their function in vitro (Yamada et al., 2002). We investigated this function in maGSC-derived hepatic cells and compared the capacity of differentiated cells at day 5+15 and day 5+25 to uptake and release ICG. We found that the maGSC-derived hepatocytes showed slightly and strong ICG uptake at day 5+15 and day 5+25, respectively (Fig. 7C). After 4 h, the ICG in the cytoplasm was completely released and not detectable in the hepatocytes (Fig. 7C).
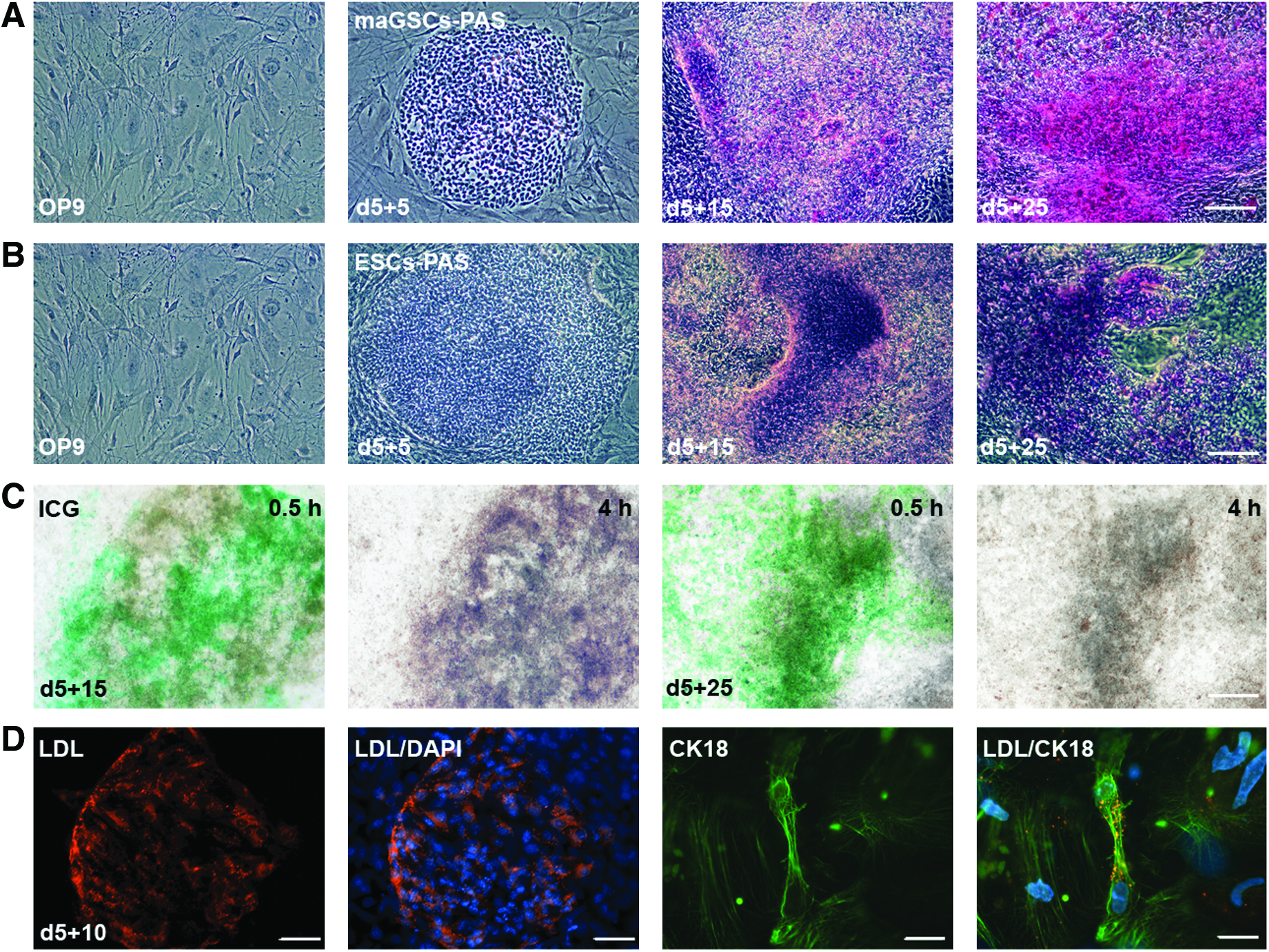
In vitro functionality of maGSC-derived hepatic-like cells. (
Another important function for maturated hepatocytes is their ability to take up low-density lipoprotein (LDL) (Zhao et al., 2009). We were able to show that significant levels of LDL were detected in the cytoplasm of maGSC-derived hepatocytes at day 5+10. In addition, we found that cells that were taking up LDL were also positive for the hepatic marker CK18 (Fig. 7D). For all functional assays, we included OP9 cells alone as control and did not observe cells positive for PAS and any uptake of ICG and LDL by OP9 cells. All of these functional assays indicated that the maGSC-derived hepatocytes are functional.
MaGSCs have similar differentiation capabilities like ESCs
Previous studies showed inherent differences in the gene expression profile of undifferentiated mouse ESCs and germ line–derived maGSCs (Assou et al., 2009; Silva et al., 2009). To investigate whether these differences in gene expression may impact on endodermal and hepatic differentiation capabilities, we differentiated mouse ESCs using our established OP9 co-culture system with 2-day AA treatment (100 ng/mL; day 0 to day 2) and compared them to maGSC-derived hepatic-like cells. We observed a similar gene expression pattern of endodermal and hepatic markers in ESCs during differentiation as we described above for maGSCs (Fig. 3). Flow cytometry analysis for quantification of AFP-, DLKA-and ALB-positive cells at different stages also showed similar results to those observed in maGSC-derived differentiation (Fig. 4B). Furthermore, the ESC-derived hepatic-like cells were metabolically active, similar to maGSC-derived cells, as shown by albumin secretion and glycogen storage (Fig. 6). PAS staining of the ESC-derived hepatocyte cultures revealed that most cells within the colonies showed the typical red cytoplasmic glycogen storage at day 5+25 in comparison to day 5+5, when no red staining was observed (Fig. 7). These data indicate that our OP9 co-culture system can be used for both pluripotent cell types, maGSCs and ESCs, giving rise to cells with a fetal hepatoblast and mature hepatocyte morphology and showing expression of genes representing definitive endoderm-derived cells, early hepatoblasts, or even mature hepatic-like cells.
Discussion
In this study, we show for the first time the successful differentiation of maGSCs into endodermal lineages and mature hepatocytes using an OP9 co-culture system. During differentiation, the cells morphologically resemble mouse fetal hepatoblasts and primary hepatocytes and express marker genes in a developmentally controlled pattern specific for hepatic precursors and mature hepatocytes. These hepatic-like cells possess functional ability in vitro and may serve as a model to delineate the differentiation process for the generation of functional mature human hepatocytes (Gai et al., 2010).
Recently, two groups reported the generation of functional hepatocytes from germ line cell–derived pluripotent stem cells in vitro (Fagoonee et al., 2010; Loya et al., 2009). In comparison to these studies, where differentiation was achieved by EBs or via a cytokine-based approach on adherent pluripotent monolayer, our OP9-differentiation protocol resulted in much higher amounts of endodermal cells and therefore terminally differentiated hepatocytes, indicated by albumin expression and secretion. In comparison to other differentiation protocols, our OP9 co-culture protocol mimics the signaling pathways of the early embryo for endoderm induction and the specification from the endoderm to the liver. Previous studies showed that cardiac mesodermal cells could induce liver specification in the mouse embryo (Lemaigre and Zaret, 2004). Co-culturing of ESCs or maGSCs with OP9 cells resulted in differentiation into hematopoietic cells, fetal liver kinase (Flk)-1–positive cardiac progenitor cells, and later into cardiomyocytes and endothelial cells (Cheng et al., 2012; Nakano et al., 1994; Narazaki et al., 2008). Our data suggest that the OP9-induced mesodermal differentiation may induce endodermal formation and hepatic specification in maGSCs and ESCs.
For induction of pluripotent stem cells to differentiate into hepatic cells, the second important factor is the induction of the definitive endoderm itself. It was reported that AA could efficiently induce ESCs to form definitive endoderm in vitro, mimicking the related factor Nodal that signals through the same receptor (Hay et al., 2008; Kubo et al., 2004). In our study, we showed that a high concentration of AA could induce maGSCs and ESCs efficiently to differentiate into hepatic progenitors and mature hepatocytes in the OP9 co-culture system. Our data are in line with previous findings from ESC-derived hepatic progenitors (Gouon-Evans et al., 2006; Loya et al., 2009; Soto-Gutierrez et al., 2006). In addition, AA-induced endoderm differentiation depends on the time, duration of application, concentration, culture conditions, and interactions of AA with various signaling pathways and differentiation factors (Sulzbacher et al., 2009). Our data show that the treatment of maGSCs with a high concentration of AA for the first 2 days during differentiation is sufficient to induce pluripotent stem cells committing to the definitive endoderm in the OP9 co-culture system. Previous studies showed that the simultaneous activation of Wnt and activin signaling pathways was essential for primordial streak formation, a requirement for the definitive endoderm development (Gadue et al., 2006; Nakanishi et al., 2007). To enhance the amount of maGSC-derived definitive endoderm differentiation, it would be interesting to determine whether the inductive effect of AA can be enhanced by the factor Wnt3a or by using Wnt3a-secreting cells.
In our study, we showed that maGSCs and ESCs could differentiate into functional hepatocytes in vitro using the combination of the OP9 co-culture system and AA treatment, and this was demonstrated by their expression of hepatic-specific genes and proteins as well as by their functionalities, such as secretion of ALB and urea, uptake and release of ICG, PAS staining, and uptake of LDL. However, our data also indicate that the maGSCs-derived hepatocytes are not terminally mature as adult hepatic cells in the liver. This was shown by their lower expression of late mature hepatic cell markers Alb, Aat, and Cy7a1and by their lower ALB and urea secretion, when compared to adult liver cells. In our culture, only 26% of cells were positive for ALB at day 5+25. As a positive control, we used hepatocytes isolated from adult mouse liver, which showed over 99% of cells positive for ALB with a very high albumin secretion. The levels of albumin secreted by maGSC-derived ALB-positive hepatic cells represented only ∼10% of those secreted by adult hepatocytes. This is similar to the albumin secretion rate of differentiated hepatic cells after promoter-based cell sorting reported previously, showing that levels of albumin secreted by ALB-positive cells represented ∼7% of those secreted by adult hepatocytes (Gouon-Evans et al., 2006). In addition, we found that high amounts of AFP- and DLK1-positive hepatic progenitors were detectable in the culture at very late stage (day 5+25). This phenomenon was observed in several other studies (Gai et al., 2010; Gouon-Evans et al., 2006; Soto-Gutierrez et al., 2006). These findings suggest that differentiation toward a terminally mature hepatocyte phenotype has not been reached in these in vitro differentiation systems and needs to be further investigated.
To enhance the maturation of hepatic-like cells derived from maGSCs in vitro, we used factors like bFGF and HGF and a hepatocyte growth and maturation medium as described before (Snykers et al., 2009). Previous studies showed that the exogenous signal molecules FGF2 (bFGF) and FGF4 had an additive effect on endoderm induction in combination with AA (Niswander and Martin, 1992; Shiraki et al., 2008). A stepwise addition of FGF, HGF, a mixture of insulin–transferrin–sodium selenite, dexamethasone, and oncostatin M during mouse and human ESC differentiation resulted in high amounts of hepatic progenitors positive for AFP and hepatocytes positive for ALB (Agarwal et al., 2008; Shiraki et al., 2008). However, the ALB secretion of ESC-derived hepatic cells is much lower than adult liver cells (Shiraki et al., 2008). A recent study reported that the combination of three-dimensional culture and cyclic adenosine monophosphate (cAMP) signaling could promote the maturation of hepatocytes derived from human pluripotent stem cells, which displayed expression profiles and metabolic enzyme levels comparable to those of primary human hepatocytes (Ogawa et al., 2013). Furthermore, the final maturation step of pluripotent stem cell–derived hepatocytes may need (or can be achieved in) an in vivo environment. Our results showed that the generation of a high percentage of hepatic progenitors could be achieved at day 5+15 using high AA concentration and the OP9 co-culture system, suggesting that this differentiation time point may be used for the isolation of DLK1-positive hepatic progenitor cells by fluorescence-activated cell sorting (FACS). It remains to demonstrate that these cells have the potential to engraft into the damaged host tissue and function as hepatocytes after transplantation and that an in vivo environment is necessary for final hepatocyte maturation. In addition, the tumorigenicity of these hepatic progenitors needs to be studied. Previous studies showed that transplantation of ESC-derived hepatic cells resulted in teratoma formation (Payne et al., 2011). Therefore, terminal differentiation of maGSC/ESC-derived hepatocytes does not only improve their function, but may also reduce the risk of tumor formation after transplantation.
In conclusion, here we report for the first time the efficient generation of high amounts of germ-line stem cell–derived AFP- and DLK1-positive hepatic progenitors and ALB-positive mature hepatocytes using the OP9 co-culture system in combination with AA. The DLK1-positive cells can be isolated by FACS and are ready for transplantation and immunological studies. The differentiated hepatic cells at late differentiation stages are functional under in vitro conditions. We also provide evidence that maGSCs have a similar hepatic differentiation potential compared to ESCs. However, the limitation in functional maturation of maGSC-derived hepatic cells in vitro is similar to that observed with ESCs and iPSCs. Further studies to induce maturation of maGSC-derived hepatic cells need to be performed. The maGSC-derived progenitors and mature hepatocytes can now be tested for their ability to restore the function of damaged liver in animal models.
Footnotes
Acknowledgments
We thank Sandra Georgi, Anke Cierpka, Yvonne Hintz, and Yvonne Wiegräfe for excellent technical assistance. We would further like to thank Dr. Petra Krause (Universitaetsmedizin Goettingen) for useful assistance in hepatocyte isolation. This work was supported by the Bundesministerium für Bildung und Forschung grant 01GN0966 (K.G.), and a DFG grant GU 595/2-1 (K.G.).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.