
Abstract
Abstract
Partially reprogrammed cells [preinduced pluripotent stem cells (pre-iPSCs)] commonly stall at epigenetic barriers, and this is one of the major failures in the reprogramming process. These cells can be converted to the fully reprogrammed state by reducing epigenetic blocks. In this study, we established three iPSC lines and two pre-iPSC lines induced by the doxycycline (dox)-inducible lentiviral system. In the pre-iPSC lines maintained under dox treatment (dox+), a small portion of embryonic stem cell (ESC)-like colonies spontaneously emerged after dox withdrawal (dox−), and major differentiation into fibroblast-like cells occurred. The spontaneous conversions based on the number of stage-specific embryonic antigen-1–positive (SSEA-1+) colonies were 0.006±0.004% [mean±standard deviation (SD)] for the #89-7D line and 0.016±0.004% for the #102-2D line. The SSEA-1+ colonies did not express the Nanog protein. However, the colonies showed characteristics typical of fully reprogrammed iPSCs after further expansion. To determine whether spontaneous conversion could be improved by epigenetic modification, we applied four small molecules—valproic acid (VPA), sodium butyrate (SB), trichostatin (TSA), and 5-aza-2′-deoxycytidine (5-Aza)—in both pre-iPSC lines. SB was the most effective molecule in enhancing the number of SSEA-1+ colonies (32- to 39-fold) at day 5 of dox− treatment. In addition, the expression of pluripotent genes (sox2 and nanog) was increased by SB. After exposure to SB, we found that the expression of four reprogramming factors and cell cycle–related genes was relatively increased compared to their expression following dox+ of pre-iPSCs. These changes caused by SB might play an important role in the spontaneous conversion of partially reprogrammed cells.
Introduction
I
Here, we generated mouse pre-iPSCs during iPSC generation with the doxycycline (dox)-inducible lentiviral system, including Oct4, Sox2, Klf4, and c-Myc. In the pre-iPSCs, spontaneous conversion to fully reprogrammed iPSCs occurred at a low ratio. Thus, we evaluated whether epigenetic modulators are able to enhance the conversion of incompletely reprogrammed cells to fully reprogrammed iPSCs.
Materials and Methods
Cell preparation
Mouse embryonic fibroblasts (MEFs) were derived from the CD1 strain [12.5 days postcoitum (dpc)]. MEFs were maintained in MEF medium [Dulbecco's Modified Eagle Medium (DMEM) with 10% fetal bovine serum (FBS), nonessential amino acids,
Reprogramming and grouping of reprogrammed cells
For somatic reprogramming, 1×106 MEFs (passage 3) were seeded on a 0.1% gelatin-coated 100-mm culture plate. After overnight incubation, a concentrated lentiviral soup of mouse Oct4, Sox2, Klf4, and c-Myc (OSKM, Stemgent, MA, USA) as well as reverse tetracycline transactivator (rtTA) were diluted in MEF medium at 1:100 (100 μL of each vector into 9.5 mL of MEF medium). The viral mixture was spread on MEFs when approximately 70% confluency was achieved. After 20 h, the mixture was completely removed, and fresh MEF medium was added. One day later, the infected MEFs were collected, and one-third of the cells were reseeded on a 0.1% gelatin-coated plate with 10 mL of MEF medium. After 1 day, the MEF medium was completely removed, and mouse iPSC medium (DMEM with ES-FBS, nonessential amino acids,
AP staining was performed on the picked colonies using the Alkaline Phosphatase Detection kit (Millipore) according to the manufacturer's instructions.
Immunocytochemistry
For immunostaining, the pre-iPSCs were fixed with phosphate-buffered saline (PBS) containing 4% paraformaldehyde (PFA; Sigma-Aldrich, MO, USA) for 5 min at room temperature. Then, the cells were subjected to immunostaining using the primary antibodies anti-Nanog (1:100, Millipore), anti-Oct4 (1:100, Millipore), and anti-SSEA-1 (1:100, Millipore). After 1×PBS washes, antibody binding was visualized using immunoglobulin G (IgG)/IgM conjugated with Alexa Fluor 488 (1:100, Millipore), and the nuclei were stained with Hochest 33258 (1:10,000, Invitrogen, MD, USA). Images were obtained using a fluorescence microscope (Axiovert, Carl Zeiss, Germany).
Quantitative reverse transcription PCR
Total RNA was extracted with TRIzol® regent (Invitrogen). cDNA was synthesized with Accupower RT Premix (Bioneer, Republic of Korea). Quantitative real-time PCR (qRT-PCR) analysis was performed using the iCycler iQ™ Real-Time PCR Detection System and the iQ™ SYBR Green Supermix (BioRad, CA, USA). The primer sets nanog, stella, rex1, oct4, sox2, kf4, and c-myc were used for detection (Table 1). To detect expressions of transgenes (OSKM) in pre-iPSCs, RNA was extracted from #89-7D and #102-2D lines cultured with dox+ and dox−, respectively. The samples of #89-7D and #102-2D under dox− were maintained for 5 days after dox withdrawal and then RNA was extracted. The primer sets of Tg-OKSM are shown in Table 1. All reactions were carried out in duplicate, and gene expression was normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as an internal standard. The relative expression of each gene was quantified as threshold cycles for amplification using the 2ΔΔCt method.
Bisulfide sequencing
Genomic DNA was extracted from each set of cells using a QIAamp DNA Micro Kit (Qiagen, Germany). Bisulfite conversion was performed with 500 ng of genomic DNA using the reagents provided in the EZ DNA Metylation-Gold™ Kit (Zymo Research, CA, USA). The promoter region of mouse oct3/4 and nanog was amplified using primers as in the study by Wernig et al. (2008) (Table 1). The purified PCR products were inserted into a plasmid vector using a TOPO TA Cloning® Kit (Invitrogen) according to the manufacturer's instructions and transformed into One Shot® Top10 competent cells (Invitrogen). Clones of each sample were verified by sequencing with the T7 universal primer.
In vitro differentiation
To confirm the differentiation potential in vitro, iPSCs were dissociated with 0.05% trypsin-EDTA and resuspended in EB medium (DMEM with 15% FBS, nonessential amino acids,
Teratoma formation
For teratoma assays, 2×106 cells were prepared in 1×PBS solution and mixed with Matrigel (1:1) on ice. The mixture was subcutaneously injected into the dorsal hind limbs of 5- to 6-week-old severe combined immunodeficient (SCID) mice (C.B-17/IcrHsd-Prkdcscid; Harlan Lab., Indianapolis, IN, USA). Three to 4 weeks later, the teratomas were dissected. Tumor masses were fixed with 1×PBS containing 4% PFA. Paraffin-embedded tissues were sectioned and stained with Hematoxylin & Eosin (H&E) and analyzed in the Cell Imaging-Histology core facility at the Quarantine & Inspection Agency (QIA). All animal experiments were performed according to the Code of Laboratory Animal Welfare Ethics, Animal, Plant and Fisheries Quarantine & Inspection Agency (QIA), Republic of Korea.
Small molecules
The epigenetic modulators valproic acid (VPA; Sigma-Aldrich) and sodium butyrate (SB; Sigma-Aldrich) and trichostatin A (TSA; Sigma-Aldrich), a HDAC inhibitor, and 5-aza-2′-deoxycytidine (5-Aza; Sigma-Aldrich), a DNMT inhibitor, were used in this study. Two pre-iPSC lines were exposed to each chemical for 48 h, and then the mouse iPSC medium of dox− was changed. After posttreatment, we evaluated the spontaneous reprogramming efficiency at day 5 of dox−.
Statistics
The data are expressed as the mean±standard error (SE). The statistical significance of differences was determined using Statistica software (Version 5.5) by performing a one-way analysis of variance (ANOVA) with post hoc comparisons between the control group and each treatment group using Duncan's multiple comparison tests. A p value<0.05 was considered a statistically significant difference between two groups.
Results
Selection of two different populations during iPSC induction
The dox-inducible lentiviral transgene delivery system was applied to reprogram mouse MEF cells (Fig. 1A). After the introduction of four reprogramming factors to MEF cells, morphological changes were observed 3 days after transgene activation via dox treatment (dox+). ESC-like colonies emerged approximately 2 weeks later, and these colonies were positive for AP staining (Fig. 1B). To obtain iPSCs, ESC-like colonies were picked by glass needle under a stereomicroscope. Out of the 104 colonies selected, 25 colonies were strongly AP+ whereas the other colonies appeared entirely AP− or partially AP+ (data not shown). Twenty-five AP+ clones showed a different growth pattern during extended subculture under dox+. Two groups were categorized by distinct morphological differences (Fig. 1C). Five clones showed a typical ESC-like morphology (Group #1), whereas the other 20 colonies were composed of a heterogenic cell population (Group #2). Interestingly, after dox withdrawal (dox−), five clones maintained an ESC-like shape, whereas 20 colonies were differentiated into mainly fibroblast-like cells (Fig. 1D). The data showed that Group #1 maintained a fully reprogrammed state regardless of dox treatment and Group #2 represented pre-iPSCs generated by transgene activation with dox treatment.
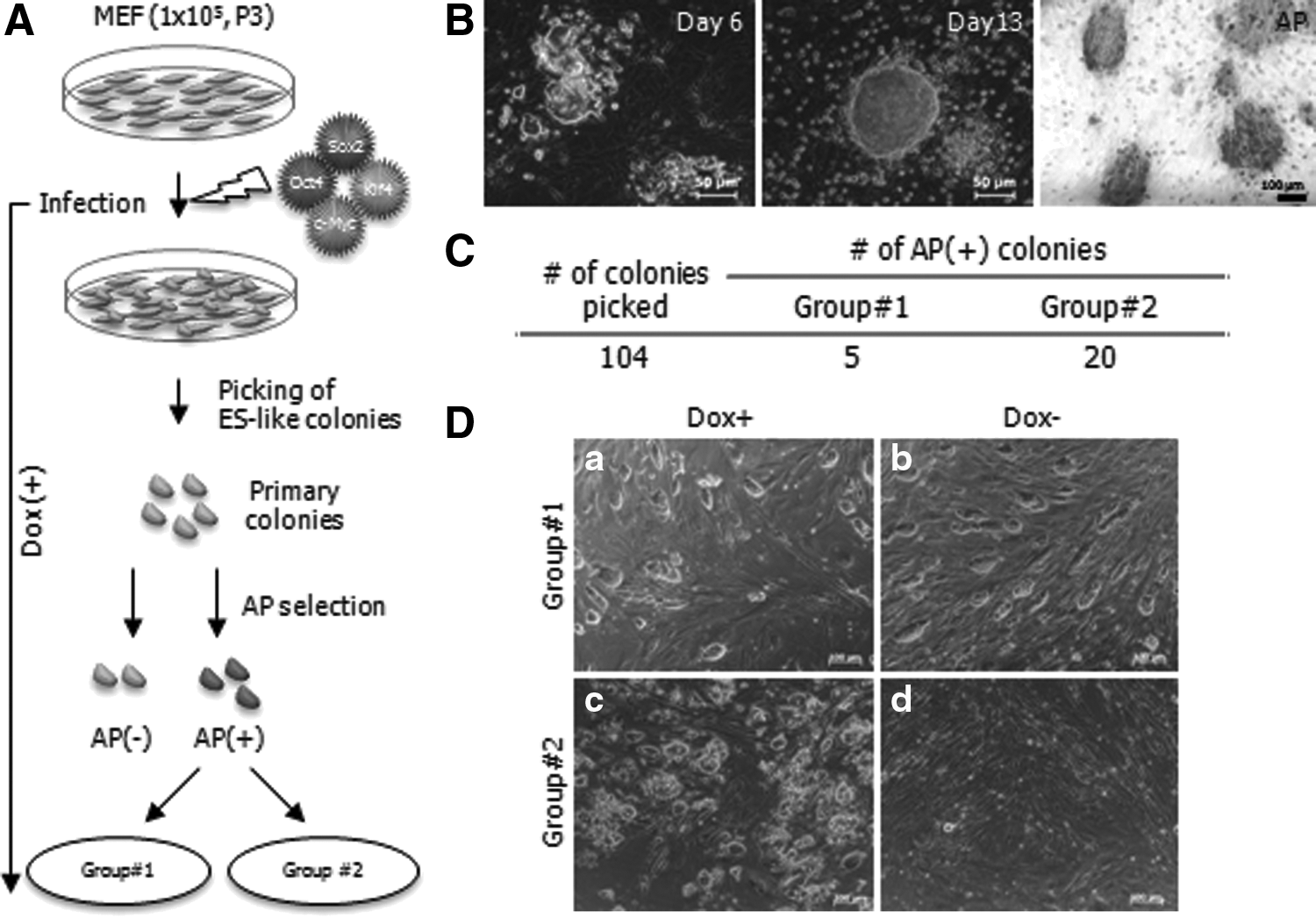
Induction of mouse iPSCs via a dox-inducible lentiviral system. (
Characterization of the reprogrammed cells of Group #1
To characterize the reprogrammed status in Group #1, three cell lines (#15, #60, and #91) were selected. They showed strong AP+ (Fig. 2A) and reacted with pluripotent markers such as SSEA-1, Oct4, and Nanog antibodies (Fig. 2B). Gene expression of pluripotent markers was confirmed by qRT-PCR. The expression of nanog, stella, rex1, oct4, and sox2 was elevated, but klf4 and c-myc was decreased compared with that in MEFs. The expression patterns of the above genes in Group #1 were very similar to ESCs (Fig. 2C). In addition, the CpG sites of oct3/4 and nanog promoter regions were demethylated, like ESCs (Fig. 2D). To identify the differentiation potential, formation of EBs and teratomas was assessed. Subsequently, the three lines differentiated into endoderm, mesoderm, and ectoderm in vitro (Fig. 2E) and in vivo (Fig. 2F). On the basis of the present data, three lines of Group #1 demonstrated fully reprogrammed iPSCs having genetic status and differentiation potential similar to ESCs.
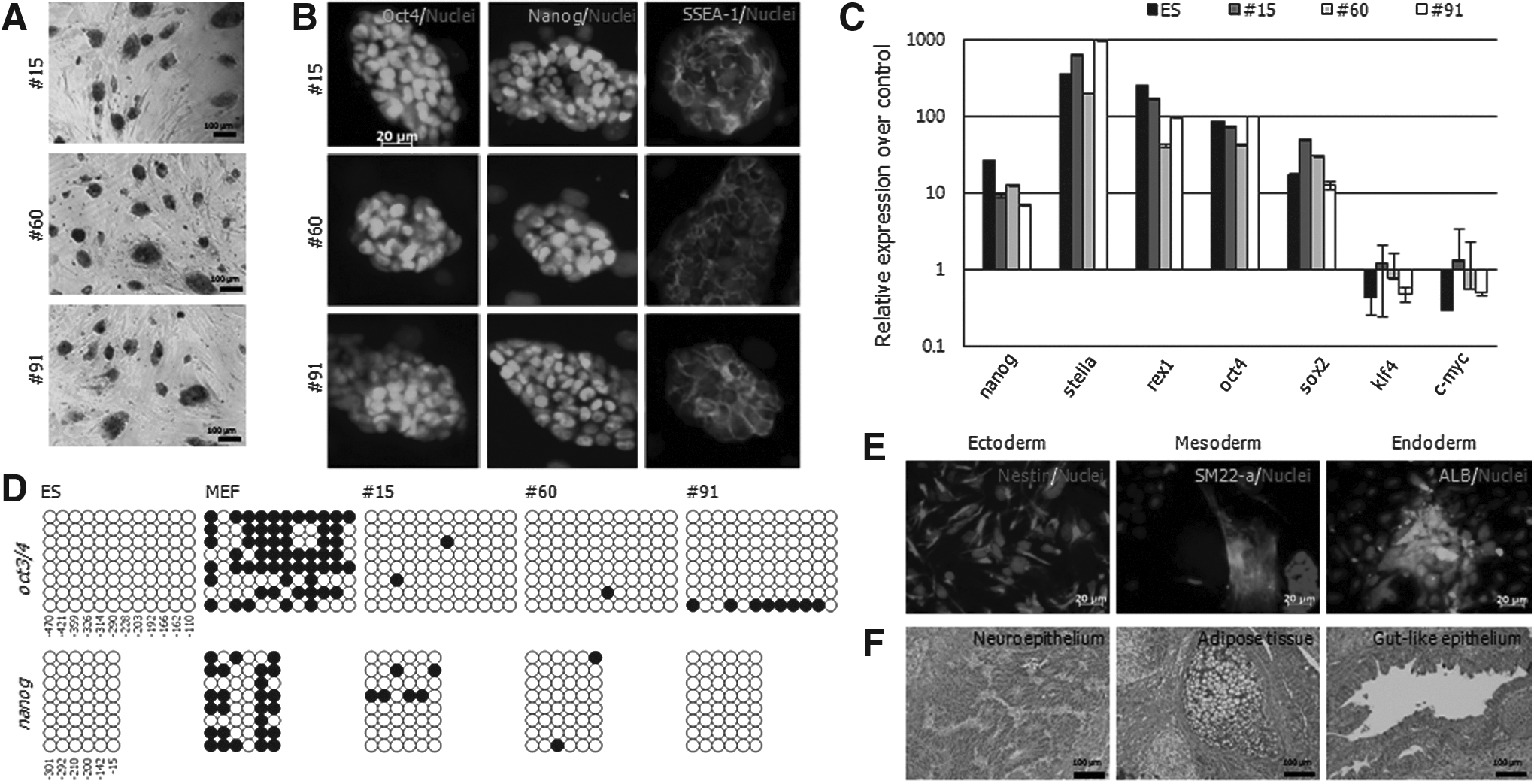
Characterization of putative iPSCs in Group #1. Out of five, three putative-iPSC lines (#15, #60, and #91) were characterized by pluripotency assay: (
Establishment of fully reprogrammed cells from pre-iPSCs
Most clones in Group #2 exhibited morphological heterogeneity and maintained overexpression of the exogenous factors with dox treatment. The morphological heterogeneity was thought to originate from cell populations with different genetic backgrounds. To exclude colony heterogeneity, we cloned two lines (#89-7D and #102-2D) originating from single cells of #89 and #102 under dox+, respectively. Despite the identical genetic backgrounds, both cloned cell lines exhibited a morphologically mixed population (Fig. 3A), strongly expressed Oct4, Sox2, Klf4, and c-Myc (Fig. 3B), and were maintained stably during continuous subpassages. In addition, both cell lines displayed hypermethylation states on the promoter regions of oct3/4 and nanog (Fig. 3C) and reacted strongly with SSEA-1 (52.4–64.9%) and Oct4 (72.9–92.8%) but weakly with Nanog (0.09–0.21%) antibodies (Fig. 3D). The above data proved that the two lines were intermediate states, not fully reprogrammed states. Interestingly, a small portion of colonies emerged from both intermediate lines after dox−, even though most cells differentiated to fibroblast-like cells. These colonies developed into ES-like cells (Fig. 4A, middle panel). The colonies expanded stably in vitro without dox (Fig. 4A, right panel). To confirm whether the colonies that emerged at day 5 of dox− were derived from activation of OKSM, the expression of OSKM was compared with dox+ and dox− conditions. The expression of exogenous genes in dox− disappeared entirely in #89-7D and #102-2D (Fig. 4B). Therefore, these findings confirm that the pre-iPSCs might be spontaneously converted to ESC-like colonies.
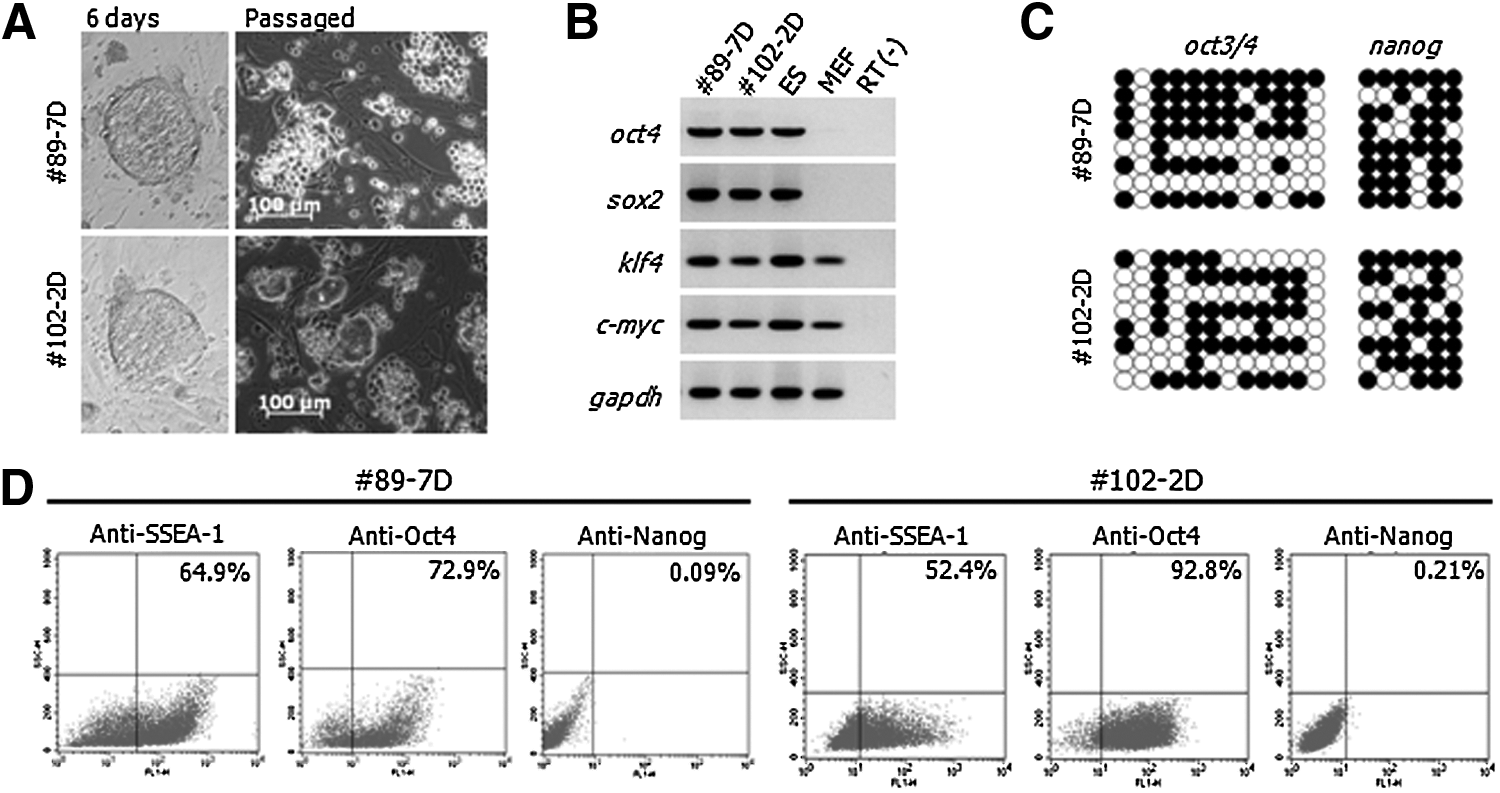
Characteristics of single cell–derived pre-iPSC lines. #89-7D and #102-2D were generated clonally from single cells of #89 and #102 maintained on a MEF feeder under dox+ and composed of a heterogenic population (mixed ESC-like colonies and cobblestone-like cells). (
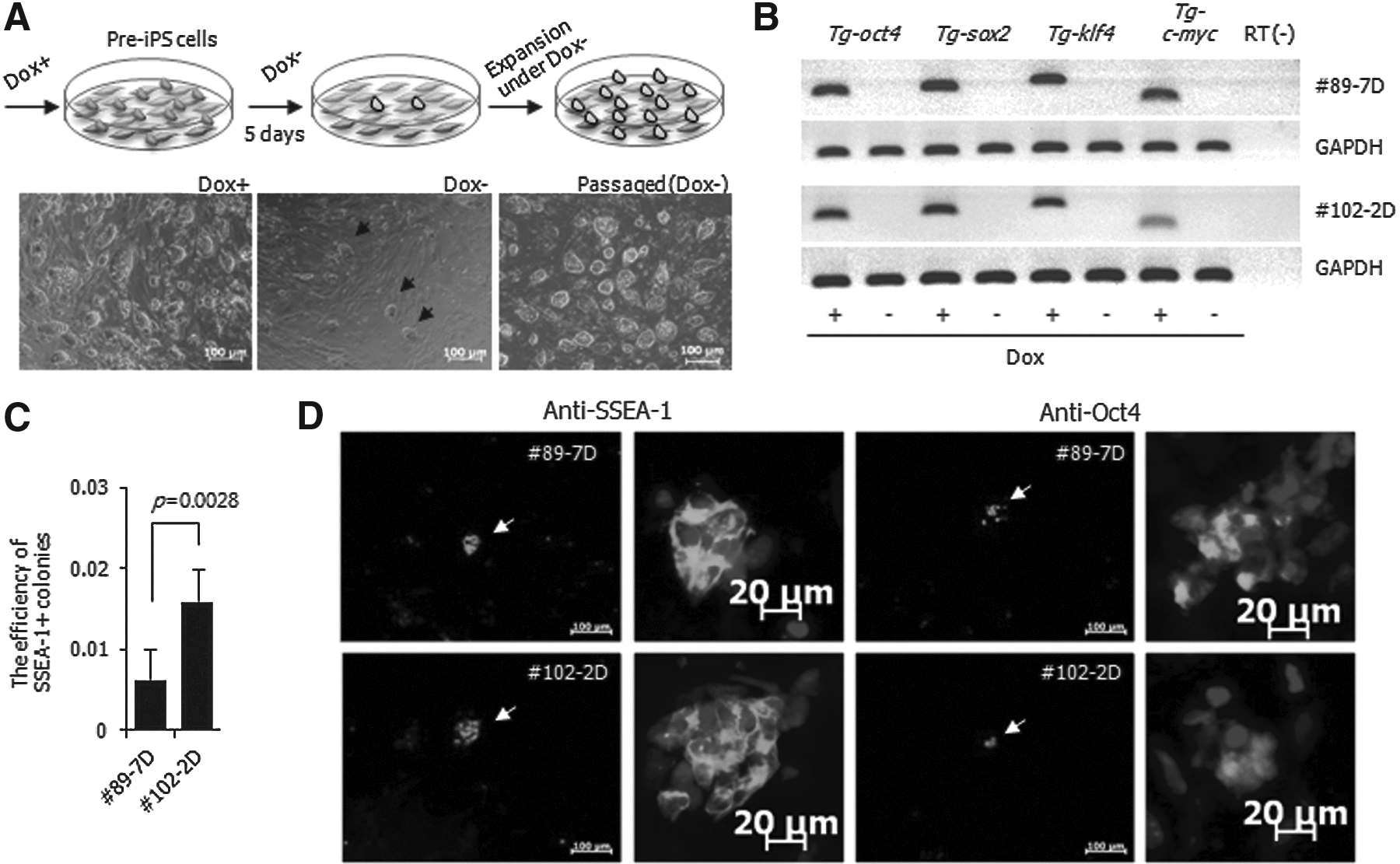
Spontaneous conversion of pre-iPSCs to fully reprogrammed iPSCs. (
To measure the efficiency of spontaneous conversion of pre-iPSCs to fully reprogrammed cells, the expression of the SSEA-1 marker protein within the colonies was determined. The expression ratio was expressed as a percentage of positive colonies out of the 50,000 cells seeded. The ratio for SSEA-1+ colonies was 0.006±0.004% [mean±standard deviation (SD)] for #89-7D and 0.016±0.004% for #102-2D 5 days after dox− (Fig. 4C). At this time point, the expression of SSEA-1 and Oct4 was remarkably decreased in the pre-iPSC lines, and subsequently the expression of these proteins was restricted in the recolonized regions (Fig. 4D). However, the expression of Nanog was not observed at recolonization. On the basis of the above data, small portions of colonies were not completely converted to a fully reprogrammed status at this time point. However, the colonies were maintained with ESC-like morphology through subpassages (Fig. 4A, right panel). Therefore, we analyzed the characteristics of the spontaneously converted colonies named as #89-7 and #102-2, respectively. Both cell lines were AP+ (Fig. 5A), reacted with SSEA-1, Oct4, and Nanog antibodies (Fig. 5B), and expressed nanog, stella, rex1, oct4, sox2, klf4, and c-myc (Fig. 5C). The CpG sites on the promoters of oct3/4 and nanog were demethylated in both sublines (Fig. 5D), whereas those sites were hypermethylated in #89-7D and #102-2D (Fig. 3C). In addition, #89-7 and #102-2 differentiated into three germ layers in vitro, which was confirmed by immunostaining with nestin, SM22-α, and albumin markers (Fig. 5E) as well as by having the capability to form a teratoma with three germ layer tissues (Fig. 5F).
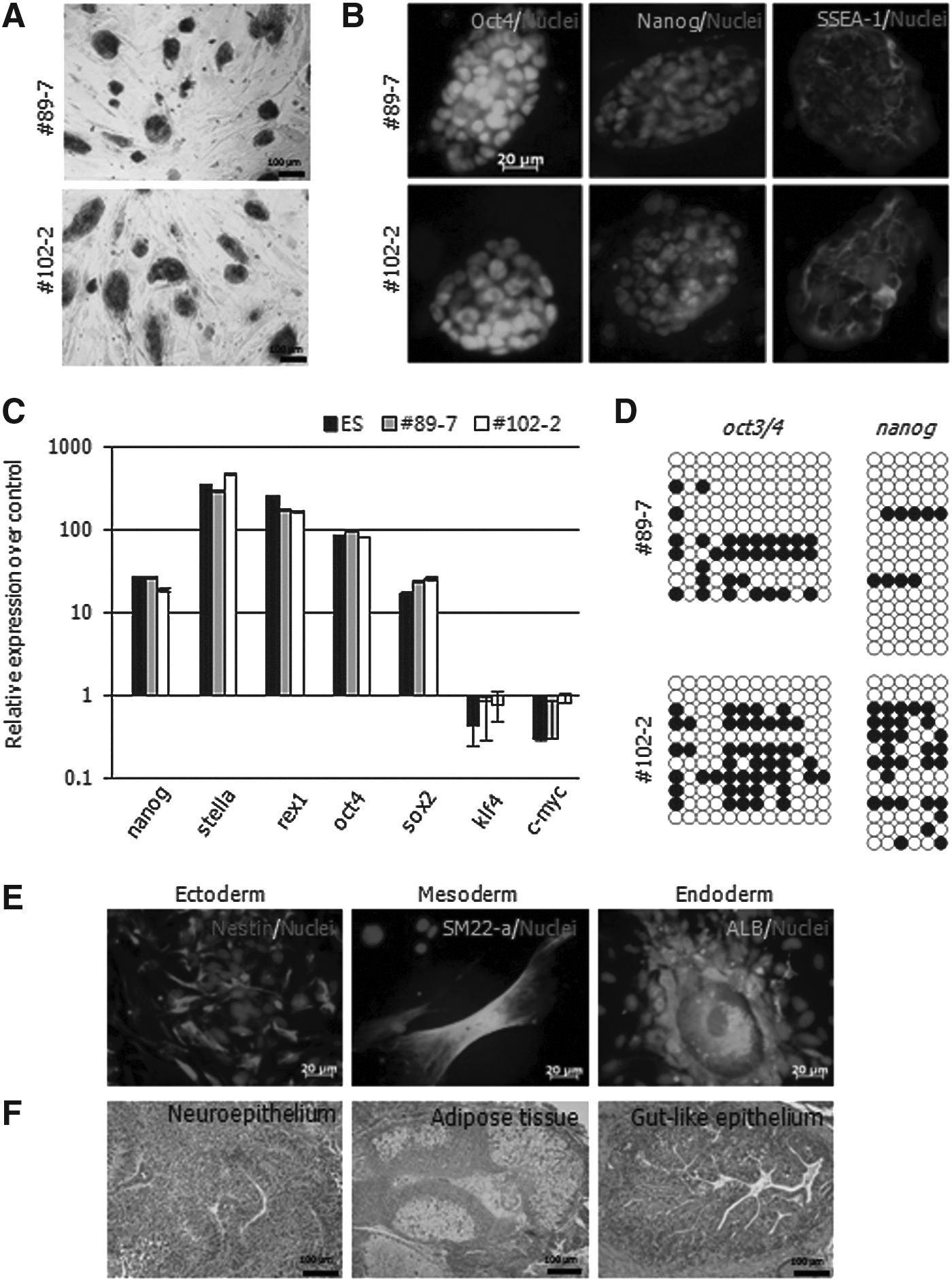
Characterization of putative-iPSCs derived from Group #2 by spontaneous events. #89-7 and #102-2 were derived from #89-7D and #102-2D, respectively, and were characterized by pluripotent assay: (
Improvement of spontaneous conversion with epigenetic modulators
To improve the efficiency of spontaneous conversion, four small molecules (VPA, TSA, SB, and 5-Aza) involved in epigenetic events were applied for 48 h to the #89-7D and #102-2D cell lines, and subsequently dox was withdrawn from the media for 5 days (Fig. 6A). Because Nanog expression was not detected at this time point, the spontaneous conversion efficiency was determined by counting the number of SSEA-1+ colonies. The number of SSEA-1+ colonies was significantly increased by 80 nM TSA (19.9-fold for #89-7D and 26.8-fold for #102-2D) and 1 mM SB (38.7-fold for #89-7D and 31.6-fold for #102-2D) (Fig. 6B). It was slightly increased by VPA (4–8 mM), but not by treatments with 5-Aza. Treatment with 1 mM SB showed the highest increase, and the quantities of sox2 increased 7.9-fold in #89-7D and 9.4-fold in #102-2D (Fig. 6C). SB treatment increased nanog expression 10-fold in #102-2D and 1.3-fold in #89-7D (Fig. 6D). Although expression of nanog gene was increased by SB at day 5 of dox−, Nanog proteins were not detectable. However, most colonies expressed Nanog protein at day 10 of dox− (Fig. 6E).
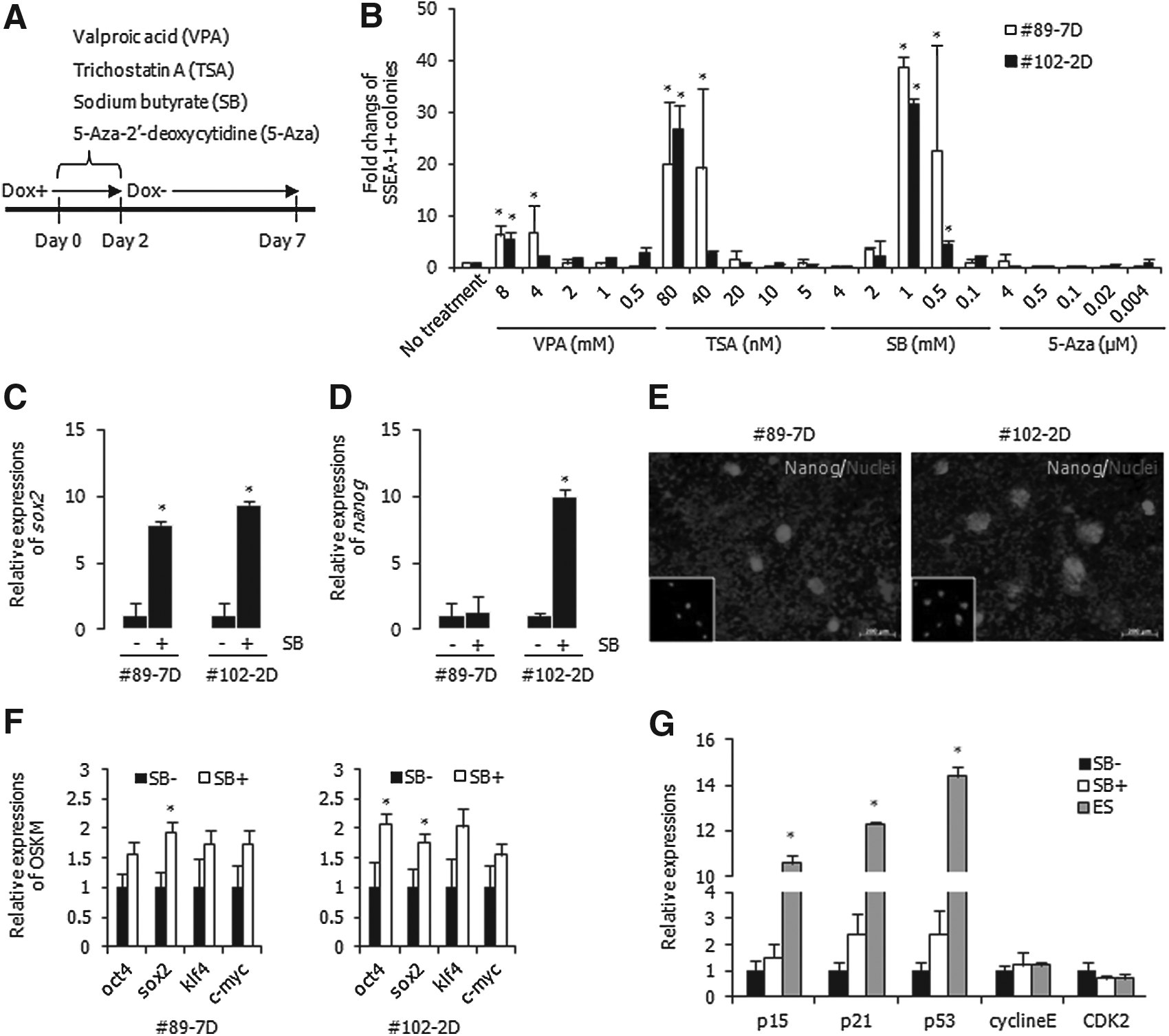
Improvement of spontaneous conversion of pre-iPSCs to full iPSCs by epigenetic modulators. (
To investigate the effects of SB on pre-iPSCs, the changes of four reprogramming factors and cell cycle–related genes were analyzed. After treatment with SB for 48 h, the quantities of the four reprogramming factors were relatively increased in #89-7D and #102-2D (Fig. 6F). The expression of p15, p21, and p53 was relatively suppressed in the pre-iPSCs compared with ESCs. However, the expression of these genes was elevated by SB treatment (Fig. 6G), even though cyclin E and CDK2 were not affected. Increased expression of transgenes and CDK inhibitors due to treatment of SB in the pre-iPSCs might elevate the efficiency of spontaneous conversion.
Discussion
In this study, mouse somatic reprogramming was induced by the four transcription factors OSKM, conditionally activated by dox. Three iPSC and two pre-iPSC lines were established. We found that a few colonies were spontaneously converted into iPSCs at a very low efficiency when exogenous genes were inactivated in pre-iPSCs, and spontaneous conversion was enhanced by HDAC inhibitors, particularly SB. The established iPSCs converted from pre-iPSCs had typical ESC-like characteristics, demonstrated by genetic and epigenetic status as well as pluripotency in vitro and in vivo.
Most somatic cells fail to achieve a fully reprogrammed status during reprogramming by transcriptional activation (up to 0.01–0.05% with the expression of OSKM). Of these, a few can be maintained as stable cell lines, also known as pre-iPSCs. Pre-iPSCs are characterized as intermediate cells with similarities to both ESCs and somatic cells (Mikkelsen et al., 2008). Typically, key pluripotency genes (i.e., Nanog) are not upregulated in pre-iPSCs, whereas lineage-associated genes are efficiently downregulated. In addition, pre-iPSCs show DNA hypermethylation of pluripotency-related genes. Those findings were supported in the present study. In this study, to eliminate heterogeneity in genetic backgrounds, single cell–derived pre-iPSC lines were established (#89-7D and #102-2D). Both lines had typical characteristics of intermediates, e.g., morphological transition from fibroblast to ES-like cells, expression of pluripotent markers (SSEA-1+ and Nanog−) and hypermethylation of oct4 and nanog loci. However, these characteristics of intermediates could be maintained only under dox+. We think that dox−-inducible pre-iPSCs may be a useful model to study key molecules required to achieve full reprogramming.
Even though transgenes were continuously activated by dox+ in the pre-iPSCs, this represented morphological heterogeneity and no Oct4 expression in some portion of #89-7D cells (approximately 73%), as shown in Fig 3D. This might be related to transgene inactivation. Transgene expression via retroviral and adeno-associated viral vectors was dramatically suppressed in culture cells, and transgene suppression was different in the cell types (Chen et al., 1997). From this perspective, it is possible that lower expression in #89-7D might have resulted from transgene silencing regardless of OKSM activation by dox.
Mikkelsen et al. reported that ESC-like colonies occasionally emerged in some pre-iPSC lines (Mikkelsen et al., 2008). They suggested that the spontaneous occurrence resulted from stochastic epigenetic events. In the present study, spontaneous conversion occurred in pre-iPSCs such as #89-7D and #102-2D after dox−. During extended maintenance of both pre-iPSC lines under dox+, however, spontaneous conversion did not occur and Nanog protein was not detected. Moreover, even a small population of spontaneous colonies with ESC-like characteristics after further passages did not express Nanog at day 5 of dox−. There are several reports that transgene silencing is a critical factor in achieving full pluripotency (Mikkelsen et al., 2008; Silva et al., 2008; Takahashi and Yamanaka, 2006). The overexpression of transgenes in pre-iPSCs might interrupt the fully reprogrammed state. Therefore, spontaneous conversion might occur stochastically during the process of transgene inactivation following dox−.
Pre-iPSCs can be efficiently induced to the full iPSC state by activating transcriptional factors and adding epigenetic modifiers. As an example, 5-Aza (500 μM) can greatly improve the transition from a partially to fully reprogrammed state (Mikkelsen et al., 2008). We also applied 5-Aza to both pre-iPSC lines to investigate their conversion to a fully reprogrammed state. Interestingly, we found no enhancement in the conversion efficiency at all 5-Aza concentrations. Moreover, a high rate of cell death was observed with relatively high doses of 5-Aza (0.5–500 μM). This cell death may indicate that apoptosis resulted from genome-wide demethylation following exposure to 5-Aza in differentiated cells (Jackson-Grusby et al., 2001). Another epigenetic modulator, an HDAC inhibitor, can stimulate complete reprogramming via histone remodeling (i.e., H3K9 acetylation). In contrast to 5-Aza, VPA, SB, and TSA increased the number of SSEA-1+ colonies in the pre-iPSCs. The optimal concentrations of SB and TSA increased the efficiency more than VPA. The effects of these chemicals also have been demonstrated in the generation of mouse and human iPSCs (Huangfu et al., 2008; Liang et al., 2010; Mali et al., 2010). In particular, SB is known to promote the generation of mouse iPSCs in early reprogramming (Liang et al., 2010), and it can facilitate human iPSC generation at a relatively later stage (Mali et al., 2010). In the present study, SB enhanced the reprogramming efficiency in mouse pre-iPSCs, which is commonly known to stall at later reprogramming stages.
SB also increased the expression of four reprogramming factors in pre-iPSCs as well as cyclin-dependent kinase (CDK) inhibitors (Fig. 6). There are many reports that SB enhances, prolongs, or reactivates the expression of transgenes. HDAC inhibitors such as SB and TSA can activate transgenes by loosening a compact nucleosome via hyperacetylation on H3 and H4 tails (Chen et al., 1997). In addition, the elevated expression of OKSM might affect the increase of the CDK inhibitors. Indeed, SB is known to inhibit cell proliferation and decrease mRNA levels of c-myc (Philippe et al., 1987). c-Myc is known to fulfill a variety of functions in cellular processes, including cell proliferation, differentiation, and growth (Dang et al., 2006). In particular, it accelerates the cell cycle by activating p15, p21, and Cyclin E (Hooker and Hurlin, 2006). During reprogramming, c-myc can facilitate the generation of iPSCs, even though it has the potential to induce tumorigenesis as an oncogene. In this study, both pre-iPSC clones showed rapid cell growth under dox+, which might be caused by transgene overexpression, in particular, c-myc. Given this data, we presume that SB is able to improve the spontaneous conversion efficiency by increasing transgene expression and modulating the cell cycle either directly or indirectly. It will be interesting to perform further studies to determine whether the efficiency of spontaneous events is affected by cellular proliferation regulated by HDAC inhibitors.
In this study, the expression of transgenes was completely silenced in early stages (Fig. 4B) and pluripotent genes were upregulated (Fig. 6C, D); however, Nanog proteins were not detected yet (Fig. 4D). Thus, the efficiency of spontaneous conversion could not be evaluated by detection of Nanog expression. We think that the Nanog protein might be expressed less in the early stage of reprogramming because we detected it at a later stage of reprogramming. SSEA-1, a pluripotency marker, was used to estimate the spontaneous conversion in this study. However, SSEA-1 is expressed at the early reprogramming stage and may be expressed despite incomplete reprogramming with no complementary activation of Oct4 and Nanog. Therefore, it is difficult to determine whether pre-iPSCs obtained complete pluripotency. Even though all SSEA-1+ cells might not give rise to fully reprogrammed cells, most of SSEA-1+ colonies expressed the Nanog protein at 10 days of dox−. According to the present experiment, SB significantly increased the number of SSEA-1+ colonies (approximately 20- to 40-fold) and the expression of sox2 and nanog. Considering this, it might take more than 10 days to achieve a completed pluripotent state in the pre-iPSCs after dox−.
In conclusion, we established mouse iPSC and pre-iPSC lines using a dox-inducible lentiviral delivery system. Furthermore, spontaneous fully reprogrammed iPSCs were established after dox− treatment and treatment with SB. The present data showed that exposure to SB can increase the expression of four reprogramming factors and cell cycle–related genes. Therefore, we assume that these changes induced by SB can contribute to improving the rate of spontaneous conversion in pre-iPSCs, even though further investigation of the interaction between transgenes and cell cycle–related genes is needed.
Footnotes
Acknowledgments
This project was supported by research funds from Animal, Plant and Fisheries Quarantine & Inspection Agency (QIA), Republic of Korea.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.