
Abstract
Abstract
Previously, we found that oocyte-secreted factors (OSFs) secreted by denuded oocytes during in vitro maturation (IVM) enhance subsequent development of bovine somatic cell nuclear transfer (SCNT) embryos. This treatment requires many oocytes during IVM. Hence, the aim of this study was to investigate whether supplementing with recombinant growth differentiation factor-9 (GDF9), one of crucial OFSs, in oocyte maturation medium could improve developmental competence of bovine oocytes and subsequent development of cloned embryos. Cumulus–oocyte complexes (COCs) from antral follicles of bovine ovaries collected from an abattoir were cultured with (SCNT+GDF9 group) or without (SCNT group) 200 ng/mL recombinant human GDF9 in oocyte maturation medium. After 22 h, metaphase II (MII) oocytes were used for SCNT. The presence of 200 ng/mL GDF9 significantly increased oocyte maturation rates, the cleavage rate, and blastocyst formation rates of bovine cloned embryos. The blastocyst total, inner cell mass (ICM) cell numbers, and ratio of ICM:TE were higher, whereas the rate of apoptosis in bovine cloned blastocysts was lower in the SCNT+GDF9 group than in the SCNT group. The histone modifications at various sites were also different between each group. These results suggest that COCs cultured with recombinant GDF9 in oocyte maturation medium improve oocyte developmental competence and subsequent developmental competence of cloned embryo in cattle.
Introduction
A
During oocyte development, the oocyte and its surrounding cumulus cells (CCs) maintain bidirectional intercellular communication, an association that is essential for both oocyte and CC development (Buccione et al., 1990). CCs provide essential nutrients and transmit molecules via gap junctions for oocytes (Gilchrist et al., 2008), and these factors play a crucial role in oocyte nuclear and cytoplasmic maturation and the acquisition of oocyte developmental competence (Chian and Sirard, 1995; Ka and et al., 1997). In turn, the oocyte secretes paracrine growth factors called OSFs that act on granulosa cells (GCs) and perform multiple functions on GCs. This bidirectional communication is important for oocyte maturation and subsequent embryogenesis. Yeo et al. suggest that insufficient levels of OSFs and disrupted oocyte–cumulus communication may the cause of compromised developmental competence of oocytes matured in vitro (Yeo et al., 2008).
Several recent studies have demonstrated that native OSFs secreted by denuded oocytes in oocyte maturation medium improve oocyte developmental competence and enhance subsequent embryo development in goat (Romaguera et al., 2010), mice (Glister et al., 2004), pig (Gomez et al., 2012), and cattle (Hussein et al., 2006; Hussein et al., 2011). Growth differentiation factor-9 (GDF9), one of crucial OSFs, regulates graulosa and cumulus cell functions in several animal species (Erickson and Shimasaki, 2003; Glister et al., 2004; Juengel et al., 2002; McGrath et al., 1995; Silva et al., 2005). COCs cultured with exogenous recombinant mouse GDF9 in oocyte maturation medium could significantly improves oocyte developmental competence and subsequent embryo development (Hussein et al., 2006; Hussein et al., 2011; Yeo et al., 2008). Previously, we also found that OSFs secreted by denuded oocytes during in vitro maturation (IVM) enhance subsequent development of bovine cloned embryos (Su et al., 2014). This treatment requires many oocytes during IVM. Hence, the aim of this study was to investigate whether supplementing with recombinant GDF9 in oocyte maturation medium could improve developmental competence of bovine oocytes and subsequent development of cloned embryos.
Materials and Methods
All chemicals, unless otherwise indicated, were obtained from Sigma (St. Louis, MO, USA). Disposable, sterile plasticware was purchased from Corning Inc. (Corning, NY, USA).
IVM of bovine oocytes
COCs collection and IVM were performed as described previously (Su et al., 2011a; Su et al., 2012). Briefly, bovine ovaries were collected from the local slaughterhouse, stored temporarily in a thermos bottle with sterile saline at 15–25°C, and transported to the laboratory within 4 h. After three washes with sterile saline, COCs were aspirated from 3- to 8-mm antral follicles using a disposable syringe (12-gauge needle). Oocytes with a compact cumulus investment and with evenly granulated cytoplasm were selected for further culture.
After three washes in phosphate-buffered saline (PBS) containing 5% (vol/vol) fetal bovine serum (FBS), some COCs were matured in vitro and were used for in IVF. Other COCs were chosen for SCNT and were cultured with 0, 20, 100, 200, and 400 ng/mL recombinant human GDF9 in oocyte maturation medium [tissue culture medium-199 (TCM-199) containing 10% FBS, 20 ng/mL epidermal growth factor (EGF), 1 μg/mL 17β-estradiol, and 0.075 IU/mL human menopausal gonadotropin] for 22 h in 95% humidified air with 5% CO2 at 38.5°C. Approximately 30 COCs were cultured in droplets of 0.4 mL of maturation medium in a 35-mm Petri dish under mineral oil. After IVM for 22 h, the cumulus cells of COCs were dispersed by vortexing. Oocytes with an extruded first polar body and uniform ooplasm were selected for SCNT.
SCNT and embryo culture
Nuclear donor cells cultures (the ear skin of a newborn female Holstein calf), SCNT, activation of reconstructed embryos, and SCNT embryo cultures were carried out in accordance with the methods of our previous studies (Su et al., 2011a; Su et al., 2011b). Briefly, after dislodgement of the cumulus cells of COCs by vortexing for 3 min in PBS containing 0.1% bovine testicular hyaluronidase in 1.5-mL centrifuge tubes, oocytes having an extruded first polar body and with uniform ooplasm were selected and stained with 10 μg/mL Hoechst 33342 for 10 min. Then, the first polar body and a small amount of surrounding cytoplasm were aspirated using a 20-μm inner diameter glass pipette in PBS microdrops containing 7.5 μg/mL cytochalasin B (CB) and 10% FBS under mineral oil. The expelled cytoplasm was surveyed under ultraviolet radiation to verify that the nuclear material had been removed. After enucleation, a single donor cell was injected into the perivitelline space of an enucleated oocyte using the same glass pipette as used for enucleation. A pair of platinum electrodes connected to a micromanipulator were used for oocyte–cell fusion, which was performed using a double electrical pulse of 35 V for 10 μsec in microdrops of Zimmermann's fusion medium covered with mineral oil. After fusion, the reconstructed embryos were kept for 2 h in G1.5 medium containing 5 μg/mL CB under mineral oil in 95% humidified air with 5% CO2 at 38.5°C. Activation of reconstructed embryos was carried out using ionomycin (5 μM, 3 min) and dimethynopyridine (1.9 mM, 3 h) in 95% humidified air with 5% CO2 at 38.5°C. After activation, cloned embryos were cultured in G1.5/G2.5 sequential media (Vitrolife AB, Gothenburg, Sweden). Droplets of 150 μL G1.5 covered with mineral oil were prepared in a 35-mm cell culture dish and equilibrated for 2 h in 95% humidified air with 5% CO2 at 38.5°C before loading of embryos (20 embryos/microdrop). Embryos were transferred into G2.5 droplets covered with mineral oil at 72 h of culture. Subsequent in vitro development to two-cell and blastocyst stages was monitored at 48 h and 168 h of culture, respectively (0 h being the time embryos were transferred into G1.5).
In vitro fertilization
IVF was carried out as described previously (Wang et al., 2011). Briefly, frozen–thawed sperm were washed in Brackett and Oliphant (BO) medium containing 6 mg/mL bovine serum albumin (BSA) and 20 μg/mL heparin and centrifuged twice at 500×g for 5 min. A 50-μL sperm suspension (a concentration of 2×106 sperm /mL) was added to 20–25 COCs in a 50-μL microdrop of the BO medium containing 6 mg/mL BSA and 20 μg/mL heparin under mineral oil. After 20 h of IVF, cumulus cells, and redundant sperm were dislodged from the oocytes with PBS supplemented with 0.1% bovine testicular hyaluronidase. Oocytes were washed twice with G1.5 medium and cultured in drops of 50 μL G1.5 in 95% humidified air with 5% CO2 at 38.5°C. Droplets of 150 μL G1.5 covered with mineral oil were prepared in a 35-mm cell culture dish and equilibrated for 2 h before loading of embryos (20 embryos/microdrop). Embryos were transferred into droplets of G2.5 medium at 72 h of culture. Subsequent in vitro development to two-cell and blastocyst stages was monitored at 48 h and 168 h of culture, respectively (0 h being the time embryos were transferred into G1.5).
Immunofluorescence staining
After fixation, permeabilization, and blocking, embryos were incubated with the anti-CDX2 (1:200, BioGenex), anti-H3K918ac (1:500, Abcam), anti-H3K9ac (1:500, Abcam), anti-H3K9me3 (1:500, Abcam), or anti-H3K9me2 (1:700, Beyotime) for 12 h at 4°C. Secondary antibodies were Alexa Fluor 555-labeled goat anti-mouse immunoglobulin G (IgG; Beyotime) for CDX2, Alexa Fluor 488-labeled goat anti-rabbit IgG (Beyotime) for H3K9ac, H3K9me3, and Alexa Fluor 555-labeled goat anti-rabbit IgG (Beyotime) for H3K18ac. 4,6-Diamidino-2-phenylindole (DAPI; Beyotime) was used to stain DNA. A Nikon Eclipse Ti-S microscope equipped with a 198 Nikon DS-Ri1 digital camera (Nikon, Tokyo, Japan) was used to analyze immunofluorescence.
Quantification of signal intensity of H3K18ac, H3K9ac, H3K9me3, and H3K9me2 (green fluorescence) was performed using Image-Pro plus software (v6.0, Media Cybernetics, Silver Spring, MD, USA) in accordance with the methods of our previous study (Su et al., 2012). The relative level of histone acetylations and histone methylation of embryos was represented by mean value±standard error of the mean (SEM). To quantify fluorescence intensity, the intensity levels of the SCNT+GDF9 and the SCNT groups were presented relative to the mean intensity level of IVF embryos.
Apoptosis assays
Apoptosis assays were performed using a DeadEnd Fluorometric TUNEL System (Promega, Madison, WI, USA) in accordance with the methods of our previous study (Su et al., 2011a). After the embryos were washed, fixed, permeabilized, and equilibrated, they were incubated with rTdT incubation buffer in the dark for 1 h at 37°C; 2×standard saline citrate was used to terminate the tailing reaction. Finally, DAPI was used to stain the DNA.
Statistical analysis
Statistical analyses were carried out by one-way analysis of variance (ANOVA) and least significant difference (LSD) tests using SPSS 18.0 software (SPSS Inc., Chicago, IL, USA). Significance was established at the p<0.05 level. Data were presented as mean±standard error of the mean (SEM).
Experimental design
Experiment 1. Effect of recombinant GDF9 on subsequent developmental competence of bovine embryos
To determine the effect of recombinant human GDF9 on subsequent developmental competence of cloned bovine embryos, COCs were cultured with 0, 20, 100, 200, and 400 ng/mL recombinant human GDF9 in the presence of 17β-estradiol, EGF, human menopausal gonadotropin, and FBS in the oocyte maturation medium. Oocyte nuclear maturation was indicated by first polar body extrusion. Oocytes maturation rates were evaluated after IVM for 22 h. Oocytes with an extruded first polar body and uniform ooplasm were selected for SCNT, and subsequent developmental competence of cloned bovine embryos was evaluated. In vitro development to two-cell and blastocyst stages was monitored at 48 h and 168 h of culture, respectively (0 h being the time embryos were transferred to G1.5 medium). The effect of recombinant human GDF9 (200 ng/mL) on subsequent developmental competence of bovine IVF embryos was also analyzed. Five replicates were performed; about 60 COCs per replicate were used for each group, respectively.
Experiment 2. Effect of recombinant GDF9 on embryo quality of cloned bovine embryo: Total cell numbers, number of ICM cells and trophectoderm cells, and ratio of ICM:TE in day-7 blastocysts
The total cells, trophectoderm (TE) cells, and inner cell mass (ICM) cell numbers in day-7 embryos of three groups were counted to assess the quality of embryos: SCNT group, SCNT+GDF9 group, and IVF group. The SCNT and SCNT+GDF9 groups were bovine cloned embryos developed from oocytes treated with (200 ng/mL) or without recombinant human GDF9. The IVF group was bovine IVF embryos developed from untreated oocytes. The total cell numbers of day-7 blastocysts were analyzed by counting the total number of cell nuclei using DAPI. The TE cell number was assessed using immunostaining for CDX2, a marker expressed in TE cells. The ICM cell number was estimated as the total cell number minus the TE cell number (Ono et al., 2010; Su et al., 2011a). The number of analyzed embryos was 24, 33, and 26 in the SCNT, SCNT+GDF9, and IVF groups, respectively.
Experiment 3. Effect of recombinant GDF9 on embryo quality of cloned bovine embryo: Apoptotic index in blastocysts
The apoptotic index of day-7 blastocysts from the three groups was assessed by terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay. The number of analyzed embryos was 23, 29, and 26 in the SCNT, SCNT+GDF9, and IVF groups, respectively.
Experiment 4. Effect of recombinant GDF9 on epigenetic modifications of cloned bovine embryo
Embryos of the three groups were collected at the blastocyst stage for detecting the levels of H3K18ac, H3K9ac, H3K9me3, and H3K9me2. The experiments were replicated three times. In each replication, n=7–9 per group.
Results
Effect of recombinant GDF9 on subsequent developmental competence of bovine embryos
To assess whether addition of recombinant human GDF9 during IVM could benefit development of oocyte and subsequent development of early SCNT bovine embryos, we treated oocytes with different concentrations of recombinant human GDF9 during IVM and calculated the maturation rates of oocytes and the in vitro developmental rates at the two-cell embryo and the blastocyst stage of cloned bovine embryos (Table 1). The effect of recombinant human GDF9 was observed from 100 ng/mL onward. The 200- and 400-ng/mL GDF9 amounts significantly improved oocyte developmental competence and subsequent development of cloned bovine embryos (Table 1).
Five replicates were performed; about 60 COCs per replicate were used for each group, respectively. Numbers in parentheses were development rates (mean±SEM%), whereas other numbers were total oocytes or embryos number of five replicates among groups. Development rates of two-cell embryos and blastocysts were the percent of total MII oocytes and were monitored at 48 and 168 h of culture respectively (0 h being the time embryos were transferred to G1.5 medium.
Values with different superscripts within columns are significantly different from each other (p<0.05).
GDF9, growth differentiation factor-9; COCs, cumulus–ooctye complexes; SEM, standard error of the mean; MII, metaphase II.
Addition of recombinant human GDF9 during IVM also significantly improved subsequent development of bovine IVF embryos (Table 2).
Five replicates were performed; about 60 oocytes per replicate were used for each group, respectively. Numbers in parentheses were development rates (mean±SEM%), whereas other numbers were total oocytes or embryos number of five replicates among groups. Development rates of two-cell embryos and blastocysts were the percent of total MII oocytes and were monitored at 48 and 168 h of culture respectively (0 h being the time embryos were transferred to G1.5 medium.
Values with different superscripts within columns are significantly different from each other (p<0.05).
GDF9, growth differentiation factor-9; IVF, in vitro fertilization; SEM, standard error of the mean; MII, metaphase II.
Effect of recombinant GDF9 on embryo quality of cloned bovine embryo: Total cell numbers, number of ICM cells and TE cells, and ratio of ICM:TE in blastocysts
As shown in Figure 1 and Table 3, total cell number, ICM cell number, and the ICM:TE ratio of blastocysts were significantly higher in the GDF9 supplementation group (SCNT+GDF9 group) compared with the nonsupplementation group (SCNT group) (p<0.05), reaching a similar level as IVF embryos.

Immunostaining of CDX2. Day-7 cloned and IVF blastocysts of the SCNT, SCNT+GDF9, and IVF groups were stained with DAPI and CDX2, a marker for trophectoderm. Bar, 100 μm. n=24, 33, and 26 in the SCNT, SCNT+GDF9, and IVF groups, respectively.
The cell numbers in blastocysts were estimated by counting the total number of nuclei using DAPI, and the number of TE nuclei was assessed using immunostaining for CDX2. The cell number of the ICM was estimated as the total number of nuclei minus the number of TE nuclei. The data were shown as mean±SEM.
Values with different superscripts within columns are significantly different from each other (p<0.05).
DAPI, 4′,6-diamidino-2-phenylindole; ICM, inner cell mass; TE, trophectoderm; SCNT, somatic cell nuclear transfer; GDF9, growth differentiation factor-9; IVF, in vitro fertilization.
Effect of recombinant GDF9 on embryo quality of cloned bovine embryo: Apoptotic index in blastocysts
The apoptotic index was significantly lower in the SCNT+GDF9 group than in the SCNT and IVF groups (p<0.05, Fig. 2 and Table 4).
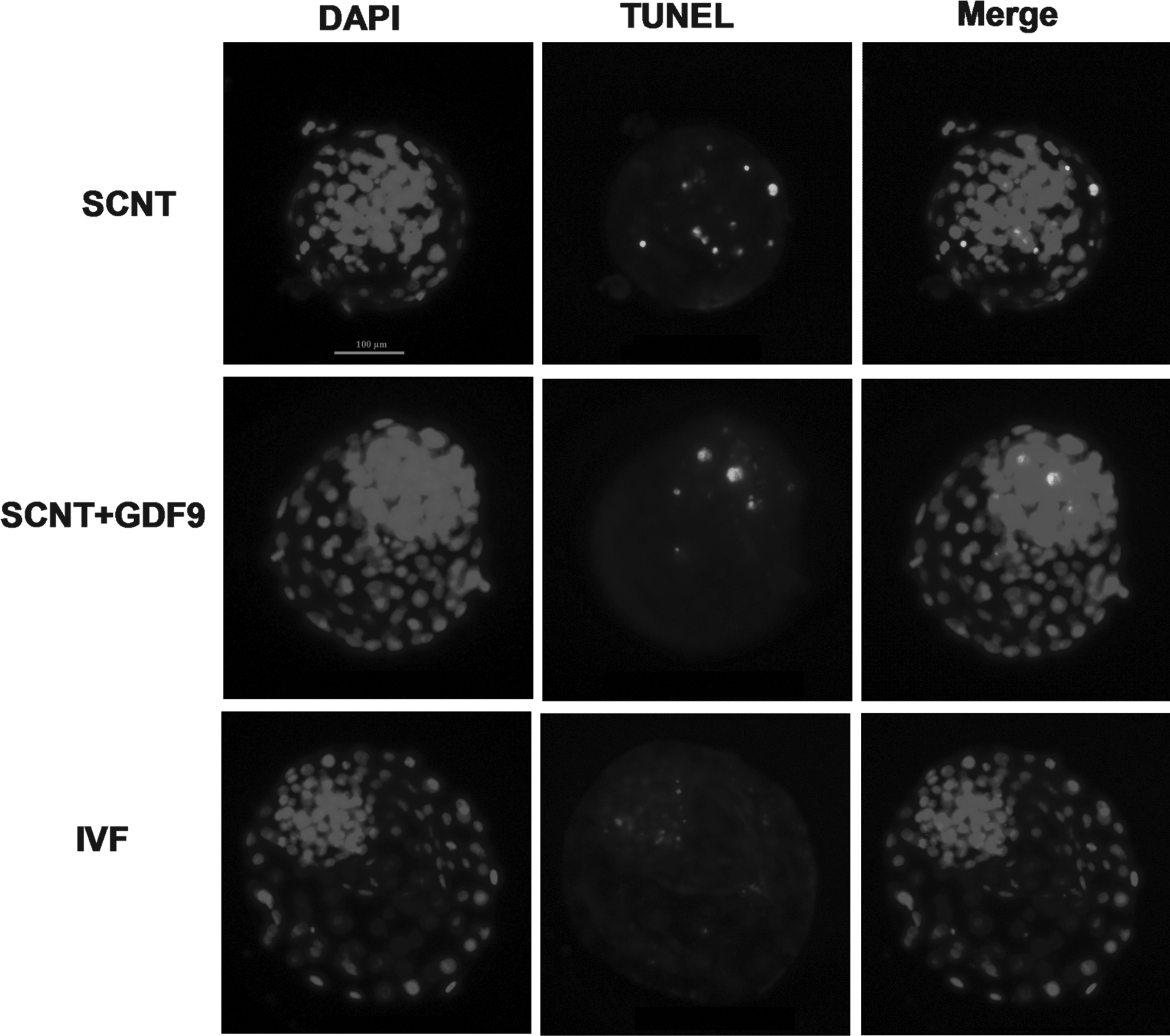
Apoptotic index of day-7 blastocysts. Representative photographs of TUNEL assay of blastocysts from the three groups. Each sample was counterstained with DAPI to visualize DNA. Bar, 100 μm.
Within the same column, values with different superscripts were different (p<0.05).
SEM, standard error of the mean; SCNT, somatic cell nuclear transfer; GDF, growth and differentiation factor-9; IVF, in vitro fertilization.
Epigenetic modifications were analyzed in embryos from the three groups
We analyzed the global acetylation levels of histone H3 at lysine 18 (H3K18ac) and 9 (H3K9ac) and global levels of histone H3 trimethylated at lysine 9 (H3K9me3) and dimethylated at lysine 9 (H3K9me2) in blastocyst stage embryos (Figs. 3 and 4). The levels of H3K18ac in cloned embryos of the SCNT+GDF9 group were higher than both SCNT and IVF groups (p<0.05) (Figs. 3A, C). The levels of H3K9ac in the SCNT+GDF9 group were also higher than the SCNT group, but no different from the IVF group (Fig. 3B, C). The cloned embryos in both the SCNT+GDF9 and SCNT groups showed a higher level of H3K9me3 than the IVF group (p<0.05, Fig. 4A, C). The levels of H3K9me3 in the SCNT group were also higher than in the SCNT+GDF9 group. There was a similar level of H3K9me2 (Fig. 4B, C) among the three groups.
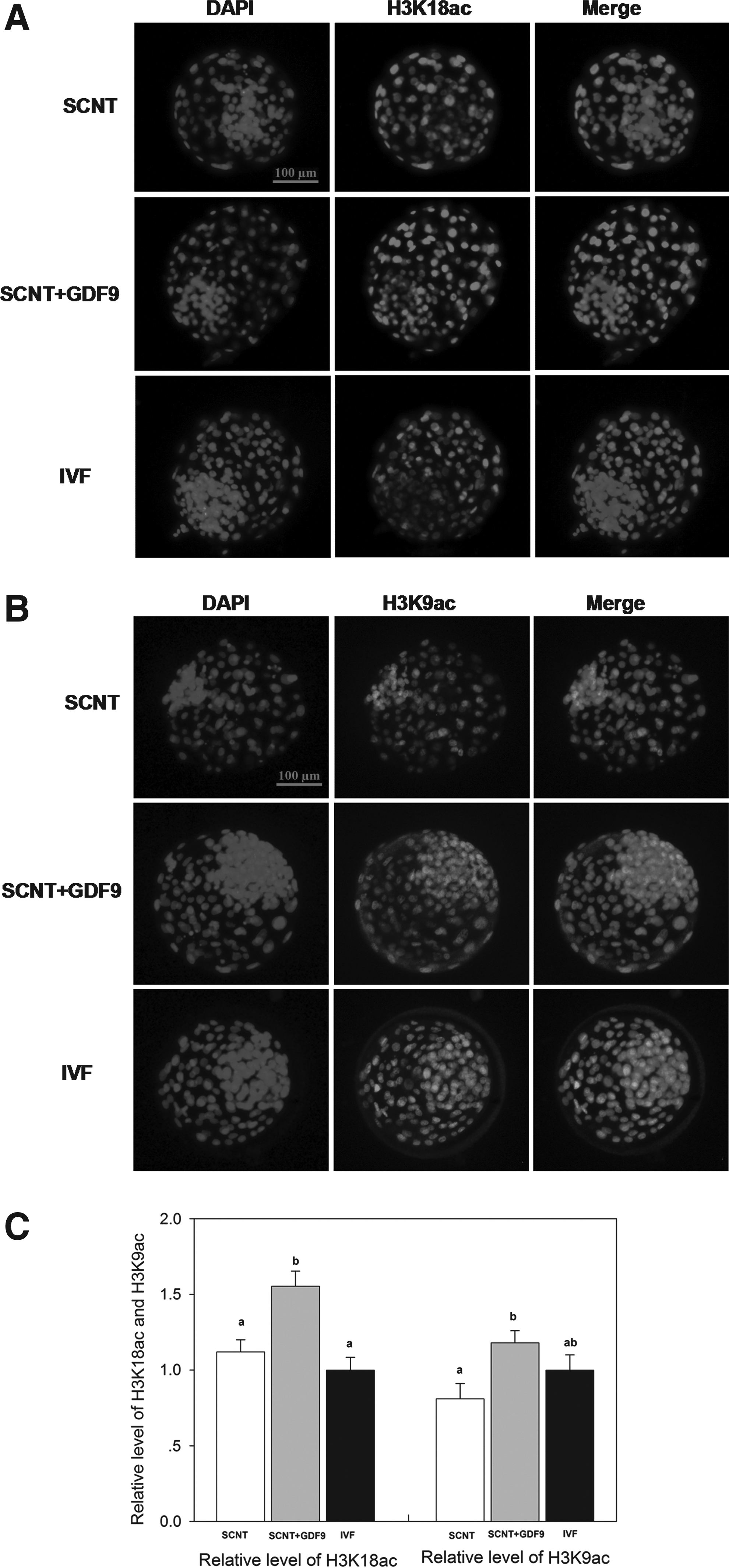
Global acetylation levels of H3K18 and H3K9 in day-7 blastocysts. Staining of H3K18ac (

Global levels of H3K9me3 and H3K9me2 in day-7 blastocysts. Staining of H3K9me3 (
Discussion
The oocyte-secreted factors regulate many of the distinctive functions of CCs, including differentiation, DNA synthesis, cellular proliferation, steroidogenesis, cumulus expansion, and cell apoptosis (Gilchrist et al., 2003; Gilchrist et al., 2008; Hickey et al., 2005; Hussein et al., 2005; Li et al., 2000; Procházka et al., 1998; Varnosfaderani Sh et al., 2013; Wang et al., 2013; Zhang et al., 2008). GDF9 and bone morphogenetic protein 15 (BMP15), as key OSFs, play crucial roles in these regulatory effects. GDF9 and BMP15 are members of the largest family of secreted proteins in mammals, transforming growth factor-β (TGF-β) superfamily. GDF9 and BMP15 proteins are essential for ovarian follicular development in sheep (Juengel et al., 2002). GDF9 has been shown genetically to control ovarian physiology. GDF9-null female mice and sheep were infertile (Dong et al., 1996; Elvin et al., 1999; Hanrahan et al., 2004), and GDF9+/−BMP15−/− double-mutant mice had more severe fertility defects than subfertile BMP15−/− mice (Yan et al., 2001; Su et al., 2004). GDF9 and BMP15 mutations were related to premature ovarian failure (Dixit et al., 2006; Laissue et al., 2006). Peng et al. (2013) showed that GDF9 and BMP15 have synergistic functions and species-specific differences in the bioactivity, and they found that mouse and human GDF9:BMP15 heterodimers are the most biopotent regulators in regulating cumulus granulosa cell functions. The GDF9:BMP15 heterodimers bind to a unique signaling complex (BMP receptor type 2-ALK4/5/7-ALK6 receptor complex) to stimulate the phosphorylation of SMAD2/3 in human and mouse granulosa cells (Peng et al., 2013).
Insufficient levels of OSFs caused disrupted oocyte–cumulus communication, and oocytes matured in vitro showed compromised developmental competence compared to oocytes developed in vivo (Yeo et al., 2008). Several recent studies have shown that OSFs secreted by denuded oocytes or exogenous recombinant OSFs (GDF9 and/or BMP15) in oocyte maturation medium enhance oocyte developmental competence, oocyte quality, and subsequent embryo development in cattle (Hussein et al., 2006; Hussein et al., 2011), goat (Romaguera et al., 2010), pig (Gomez et al., 2012), and mice (Yeo et al., 2008). However, the effects of exogenous recombinant OSFs during oocyte IVM on subsequent nuclear reprogramming and developmental competence of cloned embryos have not yet been investigated.
In this study, we found that the addition of recombinant human GDF9 during oocyte IVM improved oocyte maturation and subsequent developmental competence of bovine cloned embryos. Hussein et al. also found that the addition of 175 ng/mL recombinant mouse GDF9 or 10% vol/vol recombinant ovine BMP15 during oocyte IVM significantly increased blastocyst developmental rate postinsemination in cattle (Hussein et al., 2006; Hussein et al., 2011). In mouse, the addition of 200 ng/mL recombinant mouse GDF9 in oocyte maturation medium significantly enhances subsequent embryo development postinsemination and fetal viability (Yeo et al., 2008). We (Su et al., 2014) and several others have found that OSFs secreted by denuded oocytes (DOs) improve oocyte developmental competence and subsequent embryo development. Those results suggest that GDF9, as one of important OSFs, significantly affects oocyte developmental competence and the addition of recombinant GDF9 in oocyte maturation medium significantly enhances subsequent embryo development.
To investigate whether recombinant human GDF9 during oocyte IVM could improve embryo quality of cloned bovine embryos, we analyzed the numbers of total cells, ICM cells, TE cells in blastocysts, and ICM:TE ratio, which are usually used to assess blastocyst quality (Van Soom et al., 1996). We found that the addition of recombinant human GDF9 during IVM increased total cell number, ICM cell number, and even the ICM:TE ratio of bovine cloned blastocysts. Hussein et al. found native OSFs secreted by DOs also improved total and TE cell number of IVF blastocysts (Hussein et al. 2011; Hussein et al. 2006). Apoptotic index is another criterion for assessment of blastocyst quality (Hardy, 1997). Here, the apoptotic index was significantly lower in the SCNT+GDF9 group than in the SCNT and IVF groups. Both experiments suggest that these COCs treated with recombinant human GDF9 during IVM could generate high-quality cloned embryos.
The aberrant epigenetic nuclear reprogramming of the donor nucleus by oocytes may be the main cause of low cloning efficiency. The nuclear reprogramming process involves various epigenetic modifications, including DNA modifications (e.g., 5-methylcytosine and 5-hydroxymethylcytosine) and histone modifications (histone acetylation and histone methylation). Both histone acetylation and histone methylation play an important role in the process of nuclear reprogramming, and ultimately affects the development of cloned embryos (Dai et al., 2010; Das et al., 2010; Li et al., 2008; Wee et al., 2007; Yamanaka et al., 2009). In this study, we examined the global acetylation levels of H3K18 and H3K9 and the global levels of global levels of histone H3 trimethylated and dimethylated at lysine 9 in bovine blastocyst-stage embryos. The results showed the acetylation levels of H3K18 and H3K9 in cloned embryos of the SCNT+GDF9 group were higher than those of the SCNT group. Histone hyperacetylation could alleviate transcriptional repression, facilitate the access of various factors to nucleosomes, and facilitate chromatin remodeling (Jones et al., 1998; Nan et al., 1998). The addition of recombinant human GDF9 during IVM may improve oocyte developmental competence and oocyte reprogramming competence and subsequent improved subsequent nuclear reprogramming of cloned bovine embryo, which may associated with the improvements of cloned embryos development.
Histone methylation, another key histone modification, can be monomethylated (me1), dimethylated (me2), or trimethylated (me3) (Greer and Shi, 2012). H3K9me2 and H3K9me3 are associated with repressed chromatin and heterochromatin formation (Shi, 2007). We found the cloned embryos in both the SCNT+GDF9 and SCNT groups showed a higher level of H3K9me3 than the IVF group, which indicated that cloned embryos generated aberrant reprogramming of H3K9me3 in cattle.
In conclusion, the evidence presented in this study demonstrates that recombinant human GDF9 improves oocyte nuclear reprogramming competence and enhances subsequent development of bovine SCNT embryos.
Footnotes
Acknowledgments
The authors thank Dr. Lei Zhang for his technical assistance. We are also thankful to Mr. Younan Wang for transportation of the cow ovaries used in this study. This work was supported the National High Technology Research and Development Program of China (863 Program) (No. 2011AA100303) and China Postdoctoral Science Foundation (2013M530433).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.