
Abstract
Abstract
Faulty reprogramming of the donor somatic nucleus to a totipotent embryonic state by the recipient oocyte is a major obstacle for cloning success. Accordingly, treatment of cloned embryos with epigenetic modifiers, such as histone deacetylase inhibitors (HDACi), enhances cloning efficiency. The purpose of our study was to further explore the potential effect of valproic acid (VPA), used in previous studies, and to investigate the effect of psammaplin A (PsA), a novel HDACi, on the development and quality of cloned mouse embryos. To this aim, cloned embryos were treated with 5, 10, and 20 μM PsA or 2 and 4 mM VPA for 8–9 h (before and during activation) or 16 h or 24 h (during and after activation), and their in vitro developmental potential and blastocyst quality were evaluated. Treatments with 10 μM PsA and 2 mM VPA for 16 h were selected as the most optimal, showing higher blastocyst rates and quality. These treatments had no significant effects on the expression of Nanog, Oct4, and Cdx2 or on global histone and DNA methylation levels at the blastocyst stage, but both increased global levels of histone acetylation at early developmental stages. This was correlated with a two-fold (for VPA) and four-fold (for PsA) increase in full-term development, and a 11.5-fold increase when PsA was combined with the use of latrunculin A instead of cytochalasin B. In conclusion, PsA improves mouse cloning efficiency to a higher extent than VPA.
Introduction
U
Cloning of a variety of mammalian species through somatic cell nuclear transfer (SCNT) has proved that mature oocytes have the ability to reprogram somatic genomes in addition to gametic genomes. It has been suggested that reprogramming factors in the oocyte may be involved in loosening the highly compacted somatic chromatin through epigenetic modifications, allowing additional remodeling and transcription factors to access the genome and establish the embryo developmental program (Gurdon and Wilmut, 2011). This situation is similar, in fact, to the acquisition of a hyperacetylated and hypomethylated chromatin state in the gametic genomes, which increases their accessibility during normal development (Cantone and Fisher, 2013). However, as manifested by the low developmental capacity of SCNT embryos and the many abnormalities reported in cloned animals (Van Thuan et al., 2010; Yang et al., 2007), the oocyte ability to reprogram a somatic nucleus is limited. It is possible that, due to the different nature or the higher presence of epigenetic marks in a somatic genome in comparison with a gametic genome, the reprogramming machinery that effectively sets the developmental program after fertilization encounters major difficulties when acting upon somatic chromatin, resulting in its incomplete or incorrect reprogramming (Ogura et al., 2013). Indeed, abnormal epigenomes have been reported in SCNT embryos (Chan et al., 2012; Dean et al., 2001; Kang et al., 2001, 2002; Maalouf et al., 2008), leading to aberrant gene expression profiles (Bortvin et al., 2003; Humpherys et al., 2002; Mann et al., 2003), which in turn cause developmental arrest or the birth of animals with abnormalities, such as fetal and placental overweight, abnormal organ size, adult obesity, respiratory problems, or immunological deficiencies (Nieman et al., 2008; Van Thuan et al., 2010; Yang et al., 2007).
Current strategies to improve SCNT efficiency are focused on helping the oocyte in its reprogramming task, mainly through chemical treatment of the donor cells or the reconstructed oocytes with epigenetic modifiers. In particular, inhibitors of histone deacetylases (HDACi) and DNA methyltransferases (DNMTi), which result in increased levels of histone acetylation and reduced levels of DNA methylation, respectively, could help to open up the chromatin of the transferred nucleus and make it more accessible to the reprogramming factors present in the ooplasm. Trichostatin A (TSA), a potent HDACi, was the first epigenetic modifier successfully used in SCNT protocols. Treatment of either donor bovine somatic cells or reconstructed mouse oocytes with TSA resulted in improved embryonic development in vitro and, in the latter case, in a significant increase in the success rate of mouse cloning and of derivation of embryonic stem cells (ESCs) from the cloned embryos (Enright et al., 2003; Kishigami et al., 2006a; Rybouchkin et al., 2006). Since then, many other HDACis, such as Scriptaid, valproic acid (VPA), or oxamflatin, have been applied to SCNT protocols to enhance reprogramming and increase cloning rates (Van Thuan et al., 2010; Wakayama and Wakayama, 2010). VPA, for instance, is a short-chain fatty acid with HDACi activity that has proved to increase development and quality of mouse, bovine, and porcine SCNT embryos to a similar or even higher extent than TSA (Costa-Borges et al., 2010; Huang et al., 2011; Isaji et al., 2013; Kim et al., 2011; Miyoshi et al., 2010; Xu et al., 2012). Although the effects of each particular HDACi may vary among strains and species and some of the tested HDACis, such as aroyl pyrrolyl hydroxamide, have shown no beneficial effects (Van Thuan et al., 2009), it is clear from numerous reports that most HDACis improve nuclear reprogramming and cloning efficiency. However, whether the same applies for other epigenetic modifiers, such as DNMTis or inhibitors of histone methyltransferases (HMTi), is still uncertain.
In search for new epigenetic modifiers that could potentially improve nuclear reprogramming and increase cloning efficiency to a higher extent than those currently used in SCNT protocols, we selected psammaplin A (PsA), a promising anticancer agent that has never been used in nuclear reprogramming studies. PsA is a natural compound isolated from a marine sponge and originally reported as an inhibitor of mycothiol-S-conjugate amidase and topoisomerase II (Kim et al., 1999a, 1999b; Nicholas et al., 2002). More recently, it has been shown to inhibit proliferation of several cancer lines at nanomolar concentrations and with a low cytotoxicity through selective induction of genes related to cell cycle arrest and apoptosis (Ahn et al., 2008; Kim et al., 2007; Piña et al., 2003). Several studies have found PsA to be a potent inhibitor of HDACs in vitro (Kim et al., 2007; Piña et al., 2003), specifically of HDAC1 (Baud et al., 2012), and to induce the accumulation of highly acetylated H3 in treated HeLa cells (Kim et al., 2007). Piña et al. (2003) also reported a potent DNMTi activity of PsA in vitro in addition to its HDACi activity, but later studies found no evidence for this dual action (Baud et al., 2012; García et al., 2011).
The main aim of the present study was to test the effect of PsA on mouse cloning efficiency. In addition, we aimed to further explore the potential effect of VPA, which in previous studies by our group demonstrated a beneficial effect on both in vitro and in vivo SCNT embryo development when applied at 2 mM concentration for 8–9 h starting immediately after nuclear injection (Costa-Borges et al., 2010). Given the results of more recent studies, higher VPA concentrations and/or longer treatment durations could further increase the beneficial effects of VPA (Huang et al., 2011; Isaji et al., 2013; Kim et al., 2011; Xu et al., 2012). Therefore, in the first part of the study, we sought to determine the optimal concentration and treatment duration for both PsA and VPA in terms of in vitro development of cloned mouse embryos and blastocyst quality. Next, we investigated the effects of PsA and VPA on nuclear reprogramming by assessing the expression of pluripotency (Oct4 and Nanog) and trophectoderm (Cdx2) genes and the presence of epigenetic modifications (H3K14 acetylation, H3K9 dimethylation, and DNA methylation) in the cloned embryos. Finally, full-term development of PsA- or VPA-treated cloned embryos was determined.
Materials and Methods
Unless otherwise indicated, all reagents were purchased from Sigma (Madrid, Spain).
Animals
Mouse care and procedures were conducted according to the protocols approved by the Ethics Committee on Animal and Human Research of the Universitat Autònoma de Barcelona and by the Departament de Medi Ambient i Habitatge of the Generalitat de Catalunya.
Hybrid B6CBAF1 (C57Bl/6×CBA/J) female mice aged 6–12 weeks were used as oocyte and cumulus cell donors and, after mating with males of the same strain, for the collection of in vivo–fertilized embryos used as controls in the embryo transfer and immunostaining procedures. Outbred CD-1 females mated with normal or vasectomized CD-1 males were used as foster or surrogate mothers, respectively. All animals were purchased from Charles River (L'Arbresle, France).
Collection of oocytes, embryos, and cumulus cells
Females were induced to superovulate by intraperitoneal injection of 5 IU of pregnant mare serum gonadotropin (PMSG; Intervet, Barcelona, Spain) followed 48 h later by 5 IU of human chorionic gonadotropin (hCG; Farma-Lepori, Barcelona, Spain). MII oocytes and one-cell embryos were collected from the oviducts 14–15 h or 23–24 h, respectively, after hCG administration in HEPES-buffered CZB medium (HCZB) (Chatot et al., 1989) and treated with 300 U/mL (oocytes) or 150 U/mL (embryos) hyaluronidase in HCZB at 37°C until dispersion of cumulus cells. Denuded oocytes and embryos were then washed and kept in drops of KSOM culture medium (MR-106-D; Millipore, Madrid, Spain) covered with mineral oil at 37°C under 5% CO2. Dispersed cumulus cells recovered from oocytes were removed from hyaluronidase, diluted in HCZB, and centrifuged for 5 min at 250×g. Finally, the pellet was resuspended in a small volume of 3% (vol/vol) polyvinylpyrrolidone in HCZB and kept at 4°C until use.
Production of cloned embryos by SCNT
Oocytes were enucleated mechanically and injected with a cumulus cell nucleus using blunt-end mercury-filled pipettes (outer diameter 8–10 μm and 5–6 μm, respectively) attached to a piezo impact drill (Burleigh, Mississauga, Canada) driven by a micromanipulation device (TransferMan NK 2, Eppendorf, Hamburg, Germany) installed on an Olympus IX71 microscope (Olympus, Hospitalet del Llobregat, Spain). Micromanipulations were performed in groups of 25–30 oocytes in drops of HCZB containing 5 μg/mL cytochalasin B (CB), except in one of the experimental groups in the last set of experiments (embryo transfers) in which CB was replaced with 5 μM latrunculin A (LatA; Santa Cruz Biotechnology, Heidelberg, Germany). Afterward, the reconstructed oocytes were washed extensively and cultured for 1–3 h in KSOM supplemented or not with PsA or VPA before activation.
Parthenogenetic activation of oocytes
Chemical activation of reconstructed and intact MII oocytes was performed by incubating them for 6 h (37°C, 5% CO2) in 10 mM SrCl2 in Ca2+-free CZB medium supplemented with 5 μg/mL CB (or 5 μM LatA in one of the experimental groups in the last set of experiments) to prevent second polar body extrusion. After activation, oocytes showing at least one pronucleus were washed extensively and cultured in KSOM with or without the epigenetic modifier.
Epigenetic modifier treatments
PsA (Santa Cruz Biotechnology, Heidelberg, Germany) was dissolved in dimethyl sulfoxide (DMSO) to prepare a 3 mM stock solution and stored frozen. VPA was dissolved in 3×distilled water to prepare a 0.2 M stock solution, filtered, and stored at 4°C for 3 weeks. The final concentrations of both epigenetic modifiers were prepared by dilution of the stock solutions in the culture or activation media, depending on the experimental procedure. Three different protocols were used for the treatment of reconstructed oocytes with the epigenetic modifiers: Embryos were exposed to PsA or VPA for 2–3 h after reconstruction and 6 h of activation (8–9 h treatment) or during 6 h of activation and 10 h or 18 h of the posterior culture (16 h and 24 h treatments, respectively). At the end of the treatment, embryos were extensively washed in drops of KSOM culture medium.
Blastocyst differential staining and cell counts
Cloned embryos that reached the blastocyst stage 96 h postactivation were used for the differential staining of trophectoderm (TE) and inner cell mass (ICM) cells. Following the protocol described by Thouas et al. (2001), blastocysts were first incubated for 12 sec in a permeabilization solution consisting of 1% (vol/vol) Triton X-100 and 100 μg/mL propidium iodide (PI) in HEPES-buffered KSOM. They were then quickly transferred to a fixative solution of 100% ethanol with 25 μg/mL Hoechst 33258 (Molecular Probes, Life Technologies, Alcobendas, Spain) and stored at 4°C overnight. Finally, embryos were washed in a drop of glycerol and mounted onto a glass microscope slide in a microdrop of glycerol. A little pressure was exerted on the mounted samples to flatten the blastocysts to obtain a two-dimensional projection of the blastocyst cells and facilitate cell counting without the use of a confocal microscope (Mallol et al., 2013). Samples were examined with an Olympus IX71 epifluorescence microscope fitted with a filter that allows the visualization of PI and Hoechst staining simultaneously (U-MNV, Olympus, Hospitalet del Llobregat, Spain) and an image capture system (Cell A 2.6, Olympus, Hospitalet del Llobregat, Spain). ImageJ software (Image J 1.45q, Wayne Rasband, National Institutes of Health, Bethesda, MD, USA) was used for cell counting.
Immunofluorescence staining and image analysis
Cloned and in vivo–fertilized embryos at different developmental stages were removed from KSOM culture medium, washed in phosphate-buffered saline (PBS), fixed in 4% (vol/vol) paraformaldehyde in PBS for 15 min, permeabilized with 0.1% (vol/vol) Triton X-100 in PBS for 30 min at room temperature (RT), and stored in 0.01% (vol/vol) Tween 20 and 0.2% (wt/vol) sodium azide in PBS until approximately 15–25 embryos per stage were accumulated for each marker to be evaluated. Then both control (fertilized) and cloned embryos were blocked overnight at 4°C in PBS containing 1% (wt/vol) bovine serum albumin (BSA) and 0.05% (vol/vol) Tween 20 and stained simultaneously to reduce experimental variability.
For the detection of the pluripotency markers NANOG and OCT4, morulae and blastocysts were first incubated with either a 1:200 dilution of rabbit polyclonal anti-NANOG antibody (Abcam, Cambridge, UK) for 1 h at RT or a 1:50 dilution of rabbit polyclonal anti-OCT4 antibody (Santa Cruz Biotechnology, Heidelberg, Germany) for 1.5 h at RT, and then with 6 μg/mL goat anti-rabbit Alexa Fluor 594 secondary antibody (Molecular Probes, Life Technologies, Alcobendas, Spain) for 2 h or 30 min, respectively, at RT. In the case of blastocysts, a double OCT4 and CDX2 detection was performed, in which OCT4 staining was preceded by an overnight incubation at 4°C with a 1:100 dilution of a mouse monoclonal anti-CDX2 primary antibody (BioGenex, Fremont, CA, USA), followed by a 2-h incubation at RT with 6 μg/mL chicken anti-mouse Alexa Fluor 488 secondary antibody (Molecular Probes, Life Technologies, Alcobendas, Spain). In all cases, DNA was stained with 10 μg/mL Hoechst 33258 (Molecular Probes, Life Technologies, Alcobendas, Spain) before mounting the samples onto a glass microscope slide in a microdrop of Vectashield (Vector Laboratories, Peterborough, UK).
Acetylation of H3K14 (H3K14ac) was analyzed in cloned embryos at 10 min and 1 h after nuclear transfer and at 6 and 16 h after parthenogenetic activation, and in cloned and control fertilized embryos at the two-cell and blastocyst stages, by incubating the embryos for 1 h at RT with a 1:500 dilution of rabbit polyclonal anti-H3K14ac antibody (Millipore, Madrid, Spain). DNA was stained with 10 μg/mL Hoechst 33258 before mounting the samples. Dimethylation of H3K9 (H3K9me2) was detected in cloned and control fertilized morulae and blastocysts by incubation with a 1:500 dilution of rabbit polyclonal anti-H3K9me2 antibody (Upstate, Millipore, Madrid, Spain), for 1 h at RT. In both cases, goat anti-rabbit Alexa Fluor 594 (Molecular Probes, Life Technologies, Alcobendas, Spain) was used as secondary antibody (6 μg/mL; 2 h at RT).
Finally, for the detection of 5-methylcytosine (5meC), cloned and control fertilized morulae and blastocysts were treated with 4 M HCl at RT for 15 min or 1 h, respectively, and subsequently neutralized for 20 min with 100 mM Tris/HCl buffer (pH 8.5) before blocking. Incubation with a 1:100 dilution of mouse monoclonal anti-5meC antibody (Calbiochem, Millipore, Madrid, Spain) was performed at RT and then embryos were incubated with 6 μg/mL chicken anti-mouse Alexa Fluor 488 secondary antibody for 2 h at RT.
In morulae, a double detection of H3K9me2 and 5meC was performed, and DNA was stained with 2 μM POPO-1 iodide (Molecular Probes, Life Technologies, Alcobendas, Spain) before mounting the samples. In blastocysts, H3K9me2 and 5meC were detected separately, and whereas 10 μg/mL Hoechst 33258 was used as DNA stain for H3K9me2, when detecting 5meC DNA was stained with 500 nM PI.
All samples were examined with an Olympus Bx41 (Olympus, Hospitalet del Llobregat, Spain) epifluorescence microscope fitted with specific filters and an image-capture and analyzing system (Isis software version 5.4.5, Metasystems, Boston, MA, USA). For each antibody, images were acquired using the same exposure times and settings for all embryos and analyzed with ImageJ software for cell counting and fluorescence quantification. All individual nuclei of embryos at one-cell, two-cell, and morula stages and 30 nuclei of random regions per blastocyst were outlined for mean fluorescence intensity calculation.
Embryo transfer
CD-1 female mice were mated with vasectomized males of the same strain, and those with a vaginal plug were used as recipients at 0.5 day postcoitum (dpc). Between nine and ten two-cell cloned embryos were co-transferred with one parthenogenetic embryo into each oviduct of each female (Meng et al., 2008). As control for the embryo transfer procedure, six in vivo–fertilized embryos per oviduct were also transferred to other pseudopregnant recipients. The pups and their corresponding placentas were delivered by cesarian section at 19.5 dpc and weighed on an analytical scale. Finally, the cloned pups were fostered to CD-1 mothers that had given birth the day before.
Statistical analysis
Each experiment was repeated at least three times, and the results obtained were pooled. Data on in vitro and in vivo embryonic development were analyzed by chi-squared test or Fisher's exact test. Comparisons of blastocyst cell numbers, fluorescence intensity, and body and placenta weights were performed by Kruskall–Wallis analysis. In all cases, the GraphPad InStatTM program (v. 3.05 for Windows 95, GraphPad Software, La Jolla, CA, USA) was used. A probability value of p<0.05 was considered statistically significant.
Results
Effect of different concentrations of PsA and VPA on the in vitro development and quality of cloned embryos
In a first set of experiments, cloned embryos were exposed to different concentrations of PsA (5, 10, and 20 μM) or VPA (2 and 4 mM) for 2–3 h after reconstruction and 6 h of activation (total treatment duration 8–9 h) to determine the best concentration of each epigenetic modifier in terms of embryo development in vitro and blastocyst quality.
In the case of PsA treatments, the highest concentration (20 μM) resulted in the lowest rates of embryonic development among the three groups, although morula and blastocyst rates were only significantly lower than those of embryos treated with the intermediate concentration (10 μM). In fact, embryos treated with 10 μM PsA had also significantly higher rates of morula and blastocyst formation than nontreated embryos and embryos treated with 5 μM PsA. Blastocysts of all three PsA-treated groups showed significantly higher ratios of ICM/total cells than those of the nontreated group (Table 1).
Values with different superscripts differ significantly within the same column (p<0.05; chi-squared test for embryonic development and Kruskall–Wallis test for blastocysts cell numbers).
NT, nontreated cloned embryos; PsA, psammaplin A; VPA, valproic acid; SEM, standard error of the mean; ICM, inner cell mass; TE, trophectoderm.
Cloned embryos treated with VPA showed similar rates of activation and development and similar blastocyst cell numbers irrespective of the concentration used (Table 1). When compared with nontreated embryos, both treated groups showed higher rates of blastocyst formation, although this was only significant in the case of the 2 mM treatment. As in the case of PsA, both VPA treatments resulted in significantly higher ratios of ICM/total cells when compared with nontreated embryos.
On the basis of these results, 10 μM PsA and 2 mM VPA were considered the most beneficial concentrations in terms of embryo development and were selected for subsequent experiments. Both treatments resulted in similar rates of blastocyst formation and similar ICM/total cells ratios (Table 1).
Effect of different durations of PsA and VPA treatments on the in vitro development and quality of cloned embryos
In a second set of experiments aimed at determining the best treatment duration for each epigenetic modifier, cloned embryos were exposed to either 10 μM PsA or 2 mM VPA during 8–9 h, 16 h, or 24 h, and their in vitro development and blastocyst quality were assessed. All treatments with PsA resulted in equivalent rates of embryo development at all stages and significantly improved morula and blastocyst rates compared with the nontreated group (Table 2). But only blastocysts from the 16-h and 24-h treatments had significantly higher numbers of ICM cells and ratios of ICM/total cells than the nontreated group (Table 2). Between these two treatments, we decided to select the shorter one (16 h) for the following experiments aimed at studying epigenetic reprogramming and full-term development.
Values with different superscripts differ significantly within the same column (p<0.05; chi-squared test for embryonic development and Kruskall–Wallis test for blastocysts cell numbers).
NT, nontreated cloned embryos; PsA, psammaplin A; VPA, valproic acid; SEM, standard error of the mean; ICM, inner cell mass; TE, trophectoderm.
In the VPA group, the longest treatment (24 h) resulted in a significant decrease in morula and blastocyst rates compared with the other two treatments, and development to the blastocyst stage was equivalent to that of the nontreated group (Table 2). In contrast, VPA treatments for 8–9 h and 16 h showed significantly higher rates of blastocyst formation than the nontreated group. Whereas ICM/total cell ratios were equivalent between VPA-treated and nontreated embryos, ICM cell numbers were significantly higher in the group treated with VPA for 16 h (Table 2). On the basis of these results, the optimal duration of the VPA 2 mM treatment was established at 16 h.
It is worth noting that between the two selected treatments (2 mM VPA for 16 h and 10 μM PsA for 16 h), the one with PsA resulted in significantly higher rates of blastocyst formation than the one with VPA.
Effect of PsA and VPA treatments on pluripotency and trophectoderm markers
In the third set of experiments, fertilized, nontreated cloned embryos and cloned embryos treated for 16 h with either 10 μM PsA or 2 mM VPA were fixed at the morula and blastocyst stages and immunostained for OCT4 and NANOG pluripotency markers and the trophectoderm marker CDX2. All groups of morulae showed equivalent signal intensities for NANOG, except for those derived from VPA-treated cloned embryos, which were less intensively stained than the fertilized morulae (Fig. 1). At the blastocyst stage, the percentages of NANOG-positive cells and NANOG signal intensities were significantly lower in the three groups of cloned embryos when compared with control fertilized ones. Significantly lower levels of OCT4 were also detected in all groups of cloned embryos when compared with fertilized ones, both at morula and blastocyst stages (Fig. 2). However, treated blastocysts showed a higher percentage of OCT4-positive cells than nontreated ones, although results were only significant for the PsA-treated group. Finally, with regard to CDX2, all groups of cloned embryos showed a higher percentage of CDX2-positive cells when compared with fertilized blastocysts, but differences in signal intensities were only detected between nontreated and VPA-treated cloned blastocysts (Fig. 3).

NANOG analysis in fertilized embryos and cloned embryos nontreated (NT) and treated with 10 μM PsA or with 2 mM VPA for 16 h. (
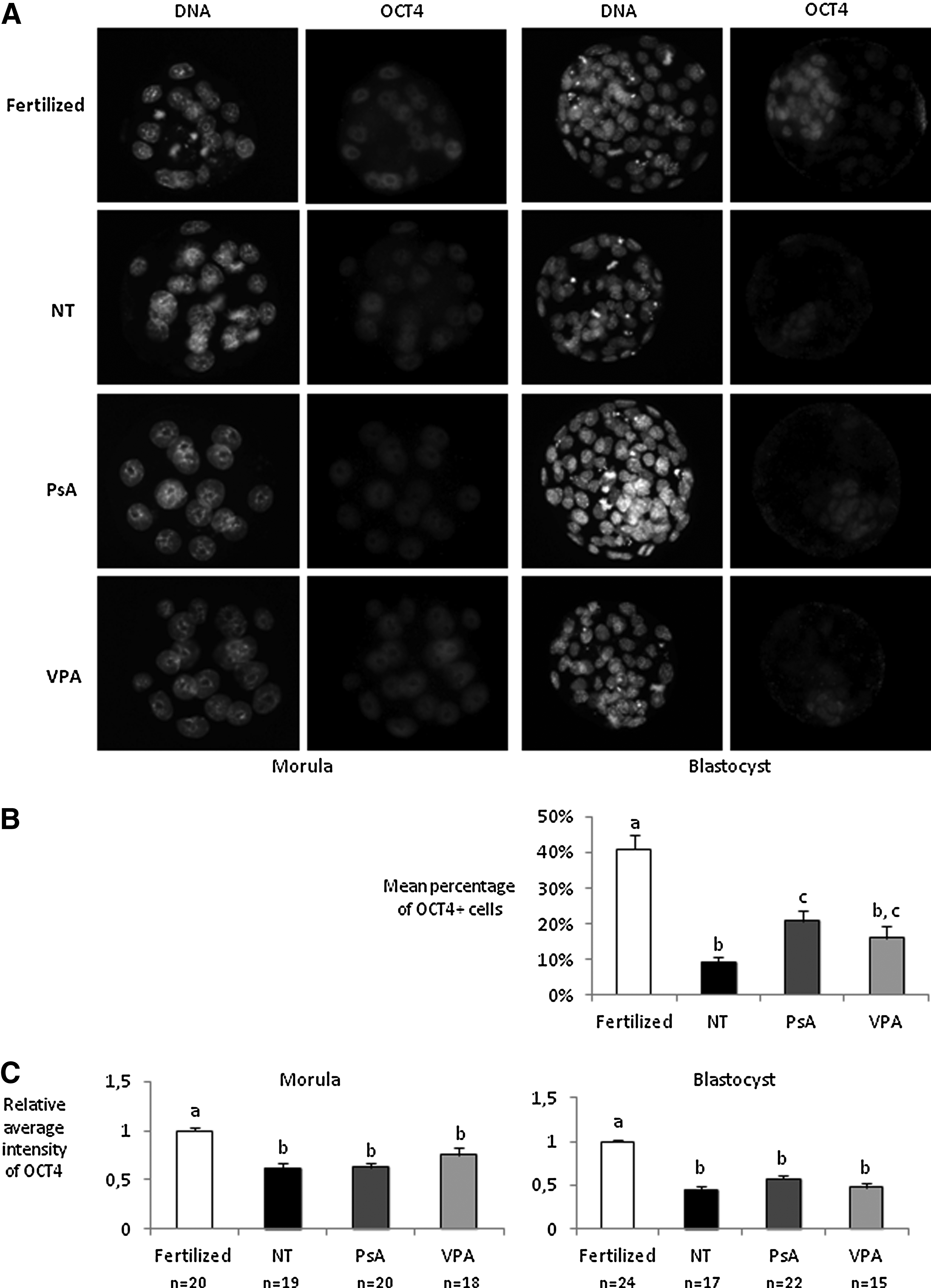
OCT4 analysis in fertilized embryos and cloned embryos nontreated (NT) and treated with 10 μM PsA) or with 2 mM VPA for 16 h. (
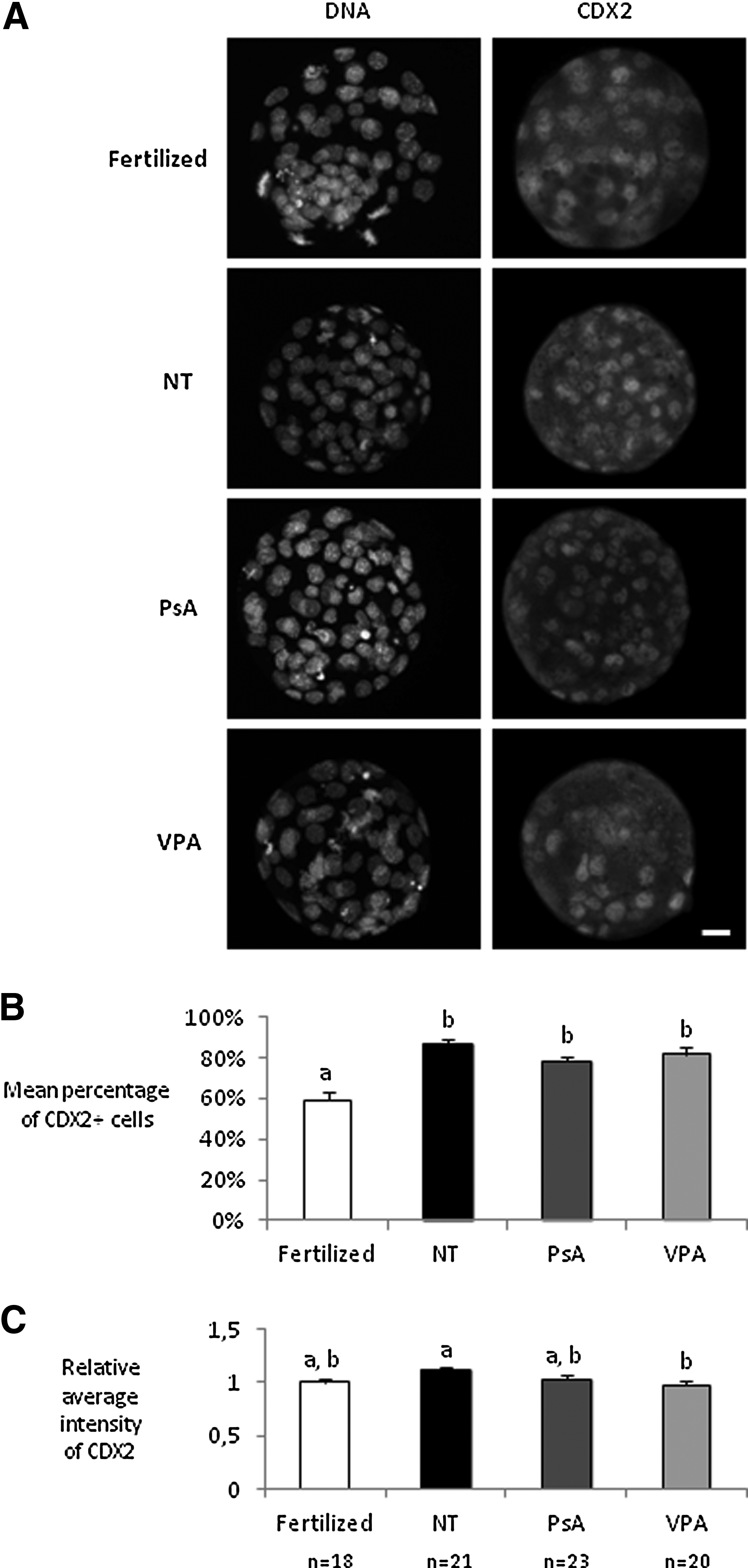
CDX2 analysis in fertilized blastocysts and cloned blastocysts nontreated (NT) and treated with 10 μM PsA or with 2 mM VPA for 16 h. (
Effect of PsA and VPA treatments on epigenetic marks
In the fourth set of experiments, fertilized, nontreated cloned embryos and cloned embryos treated for 16 h with either 10 μM PsA or 2 mM VPA were fixed at different developmental stages and immunostained for H3K14ac, H3K9me2, and 5MeC marks.
We observed that H3K14 was highly acetylated in the cumulus cell nucleus 10 min after injection, but its acetylation markedly decreased 1 h after nuclear transfer into the enucleated oocyte (Fig. 4A). By the end of the parthenogenetic activation (6 h postactivation), treated and nontreated embryos displayed similar levels of H3K14ac. At the end of the treatment (16 h postactivation), both groups of treated embryos had significantly higher levels of H3K14ac than nontreated embryos, and PsA-treated embryos were significantly more acetylated than VPA-treated ones (Fig. 4). At the two-cell stage, when mouse embryonic genome activation takes place, nontreated cloned embryos were hypoacetylated when compared with fertilized controls. PsA-treated embryos still maintained significantly higher levels of H3K14ac than nontreated ones, and were more similar to control fertilized embryos, whereas H3K14ac in VPA-treated embryos had decreased to levels similar to those found in nontreated embryos. Finally, at the blastocyst stage, nontreated cloned embryos showed similar levels of H3K14ac as fertilized control embryos. PsA-treated embryos were still significantly more acetylated than VPA-treated ones and similar to fertilized embryos, although no significant differences were found at this stage between any of the treated groups and the nontreated embryos (Fig. 4).
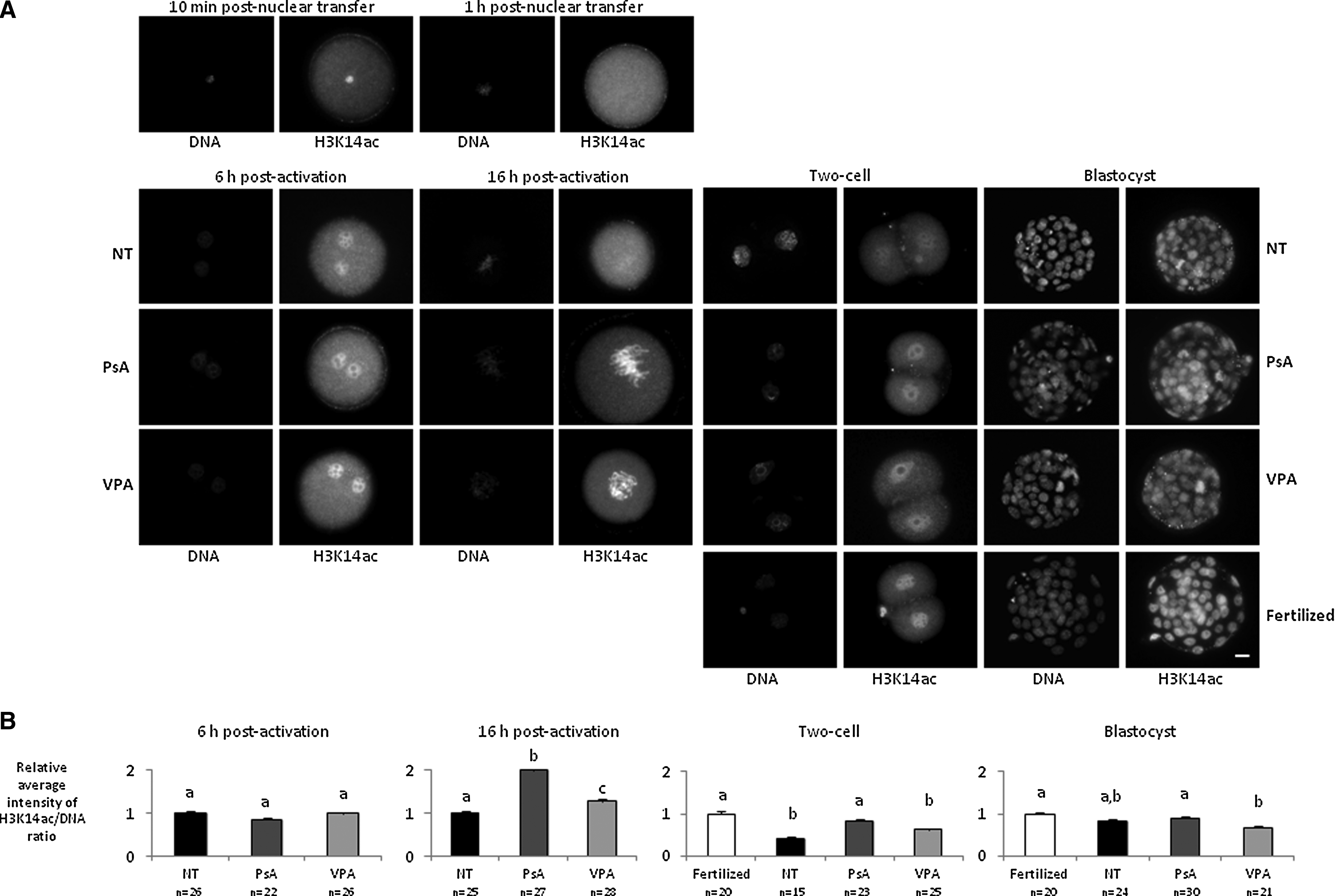
H3K14 acethylation analysis in fertilized embryos and cloned embryos nontreated (NT) and treated with 10 μM PsA or with 2 mM VPA for 16 h. (
With regard to H3K9 dimethylation, no significant differences were observed between nontreated cloned embryos and fertilized controls at any of the two developmental stages analyzed (Fig. 5). Both PsA- and VPA-treated morulae showed reduced H3K9me2 levels when compared with nontreated or fertilized ones. But at the blastocyst stage, H3K9me2 levels in the two groups of treated embryos had increased to similar levels as in the nontreated ones, and PsA-treated embryos showed even higher levels of H3K9me2 than fertilized blastocysts.
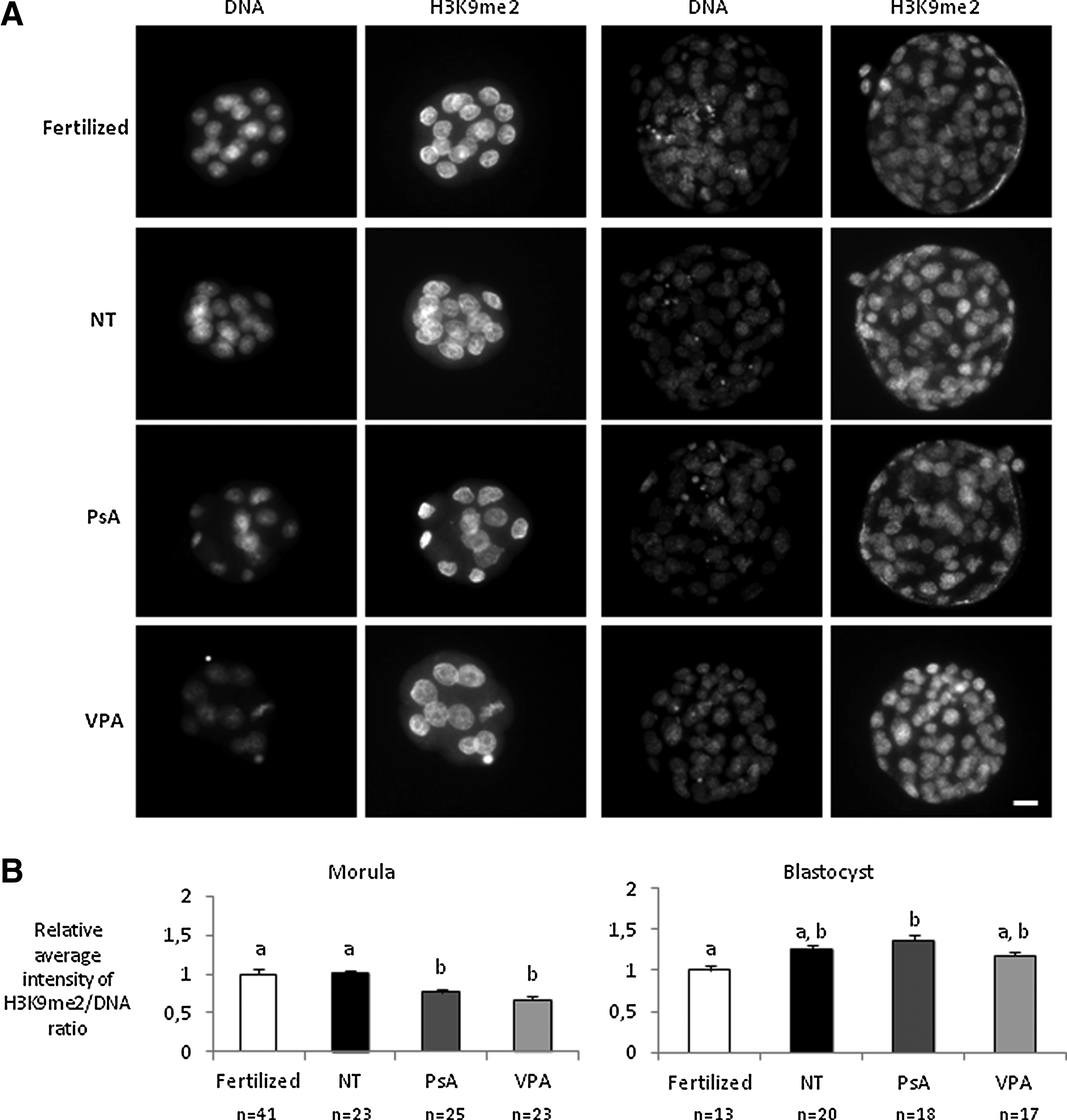
H3K9 dimethylation analysis in fertilized embryos and cloned embryos nontreated (NT) and treated with 10 μM PsA or with 2 mM VPA for 16 h. (
Finally, we observed that nontreated clones and fertilized embryos displayed similar levels of DNA methylation at the morula stage, but that the DNA of nontreated cloned embryos was significantly hypermethylated in blastocysts (Fig. 6). Both epigenetic modifiers reduced DNA methylation in the cloned morulae when compared with the untreated group, but at the blastocyst stage all three groups of cloned embryos showed equivalent levels of DNA methylation. Among the cloned embryos, only PsA-treated embryos had similar levels of DNA methylation as fertilized embryos at both morula and blastocyst stages.

DNA methylation analysis in fertilized embryos and cloned embryos nontreated (NT) and treated with 10 μM PsA or with 2 mM VPA for 16 h. (
Effect of PsA and VPA treatments on full-term development of cloned embryos
In the last set of experiments, nontreated cloned embryos and cloned embryos treated with either 10 μM PsA or 2 mM VPA for 16 h were transferred at the two-cell stage to recipient females to assess their full-term development. We found that both treatments, but especially the PsA one, resulted in higher birth rates than those obtained in the untreated group, although the differences were not statistically significant in any case (Table 3). Residual implantation sites were detected in all groups of recipient females receiving cloned embryos, but at significantly higher rates in the PsA-treated group compared with the untreated group. One placenta without a fetus was detected in the VPA-treated group. Although both pups and placentas were heavier in the cloned groups when compared to the fertilized one, differences were only significant for the PsA-treated group.
Values with different superscripts within the same column differ significantly (Fisher exact test for full-term development and Kruskall–Wallis test with Dunn posttest for body and placenta weights, p<0.05).
NT, nontreated cloned embryos; PsA, psammaplin A; VPA, valproic acid; SEM, standard error of the mean; LatA, latrunculin A.
When the actin polymerization inhibitor LatA was used instead of CB in the SCNT protocol during oocyte micromanipulation and activation, we obtained a three-fold increase in the birth rate of embryos treated with PsA (Table 3). This modification of the protocol also resulted in a significant higher number of residual implantation sites. A slight decrease in mean body and placenta weights was observed when compared to the use of CB, but the difference was not significant, and the resulting 11 pups in this treatment and their corresponding placentas were still the heaviest of all groups.
Unfortunately, the only cloned pup obtained from the untreated group was accidentally injured during the cesarean section and died soon after, and two of the cloned pups in the PsA-LatA treatment group were cannibalized by the foster mothers. Other two mice in the PsA-LatA group died prematurely due to unknown reasons. The two and four cloned pups from VPA and PsA treatments, respectively, and the remaining seven cloned pups from PsA-LatA treatment developed normally to adulthood and were able to reproduce when mated to normal B6CBAF1 males, giving rise to normal litters (6–11 pups).
Discussion
Here, we demonstrate for the first time that the epigenetic modifier PsA can improve the development and quality of B6CBAF1 cloned mouse embryos and that PsA-treated embryos can give rise to increased numbers of healthy fertile cloned mice. Furthermore, we show that PsA improves SCNT efficiency to a higher extent than the previously used HDACi VPA.
When treating cloned embryos with epigenetic modifiers, it is important to optimize the concentration and exposure time to obtain the maximum beneficial effects without embryotoxicity or other side effects. We chose a starting concentration of 5 μM PsA, because García et al. (2011) had observed an increase in histone acetylation and a decrease in HDAC1 activity, among others, in human acute myeloid leukemia cells using this concentration. Although cloned mouse embryos treated with 5 μM PsA showed an increased ICM/total cell ratio at the blastocyst stage, no improvement in in vitro developmental rates was observed. PsA concentration was then increased twofold (10 μM) and fourfold (20 μM), and whereas 10 μM significantly increased in vitro development of the cloned embryos, 20 μM did not. With regard to VPA, we previously reported a beneficial effect on both in vitro and in vivo development of cloned mouse embryos when treated at a concentration of 2 mM (Costa-Borges et al., 2010). However, the successful results obtained in recent studies with higher concentrations of VPA in porcine and bovine cloned embryos (Miyoshi et al. 2010; Kim et al. 2011; Xu et al. 2012) prompted us to investigate the effects of a higher concentration. We found that only 2 mM VPA, but not 4 mM VPA, improved embryo development. Therefore, 10 μM PsA and 2 mM VPA seem to be the optimal concentrations of these two HDACis for mouse cloning procedures.
To optimize treatment duration, we started with an 8–9 hr treatment, as used in our previous work with VPA (Costa-Borges et al., 2010). But due to the reported potential DNMTi activity of PsA (Piña et al. 2003), we decided to increase the duration of the treatment to 16 and 24 h so that the epigenetic modifiers were present during the first S phase. Whereas the oocyte form of the maintenance methyltransferase Dnmt1 is normally excluded from the nucleus at these early preimplantation stages, cloned embryos aberrantly express the somatic form of Dnmt1, and its presence in the nucleus may maintain DNA methylation levels during DNA replication and prevent passive demethylation (Chung et al., 2003; Dean et al., 2001). In addition, these longer treatments have shown beneficial effects in VPA-treated porcine and bovine cloned embryos (Miyoshi et al., 2010; Xu et al., 2012). Indeed, the 16 h treatment resulted in significantly increased blastocyst rate and quality both for PsA and for VPA and was selected as the optimal treatment duration. Interestingly, only the number of ICM cells or the ICM/total cells ratio was significantly increased as a result of both PsA and VPA treatments, whereas total cell numbers were only marginally increased, indicating that rather than affecting cell proliferation, these treatments may be increasing blastocyst quality by favoring cell allocation toward the ICM lineage.
To determine whether the increased blastocyst rate of PsA- and VPA-treated cloned embryos was correlated with an upregulation of pluripotency-related genes, the levels of OCT4 and NANOG were analyzed. In agreement with previous studies on Oct4 and Nanog expression and protein levels (Boiani et al., 2002, 2005; Cavaleri et al., 2008), we observed reduced intensities of OCT4 and NANOG signals in cloned embryos. Even though several studies have shown that treatment of cloned embryos with HDACis increases Oct4 mRNA (Jafari et al., 2011; Su et al., 2011) and protein (Isaji et al., 2013) and VPA has been shown to increase Oct4 and Nanog expression in fertilized mouse morulae (VerMilyea et al., 2009), we did not observe any significant effect of either PsA or VPA on OCT4 and NANOG protein levels. A similar result was obtained by Huang et al. (2011) for Oct4 mRNA abundance in VPA-treated cloned porcine embryos. In spite of this, both HDACis used in the present study, but especially PsA, increased the percentage of OCT4-positive cells in cloned blastocysts. Unlike with pluripotency-related markers, normal levels of mRNA and protein of the trophectoderm marker Cdx2 are usually found in cloned embryos (Kishigami et al., 2006b; Balbach et al., 2010; Cavaleri et al., 2008; Su et al., 2011), as observed in our study for CDX2 signal intensities. Taken together, our results seem to indicate that some of the genes essential for embryonic development, such as Cdx2, are more efficiently reprogrammed than others, such as Oct4 and Nanog, resulting in a failure to correctly establish and/or maintain the ICM lineage in most cloned embryos, as previously reported (Kishigami et al., 2006b). Neither VPA nor PsA treatments help to correct this defect. Even though these treatments increase the number of blastocyst cells allocated to the ICM lineage, they cannot rescue the normal levels of OCT4 and NANOG proteins present in fertilized embryos.
The extent of nuclear reprogramming occurring in the cloned embryos was also analyzed through the study of histone acetylation and histone and DNA methylation at several time points and developmental stages. The levels of H3K14ac, correlated with transcriptional activation, were lower in nontreated cloned embryos than in fertilized embryos at the two-cell stage, as expected (Wang et al., 2007), but equivalent at the blastocyst stage. Treatment with either PsA or VPA increased H3K14ac levels at the one-cell and two-cell stages, especially in the PsA-treated group, which may correlate with the potent activity of this HDACi (Baud et al., 2012; Kim et al., 2007; Piña et al., 2003). However, the levels of H3K14ac became equivalent to those of nontreated cloned embryos at the blastocyst stage. These results agree with those of Huang et al. (2011), reporting that VPA increases acetylation of H3K14 and H4K5 in cloned porcine embryos at pronuclear and two-cell stages, but not at the blastocyst stage. PsA and VPA treatments also transiently reduced the levels of two transcriptionally repressive chromatin marks, DNA methylation and H3K9 dimethylation, at the morula stage. Rather than a direct effect of these epigenetic modifiers on DNA and histone methylation levels, this is probably a result of histone hyperacetylation during the early stages of development, because a negative correlation between these types of epigenetic modifications has been reported (Dobosy and Selker, 2001). The lack of significant differences in DNA methylation levels between PsA- and VPA-treated cloned embryos at both morula and blastocyst stages argues against a potential effect of PsA as a DNMTi (Piña et al., 2003), in agreement with recent studies (Baud et al., 2012; García et al., 2011).
Histone hyperacetylation during the early stages of preimplantation development may promote a more open chromatin configuration, facilitating the access of reprogramming and transcription factors to the genome, and may therefore be responsible for the higher developmental potential of PsA- and VPA-treated cloned embryos. The more potent HDACi activity of PsA with regard to VPA may explain the slightly better results obtained with this inhibitor in terms of in vitro and in vivo development. In fact, even though full-term development results were not significantly different between the two treatments or between these and the nontreated group (a very high number of embryos should be transferred to obtain a statistically significant difference with the present offspring rates), it is worth noting that VPA increased SCNT efficiency by almost two-fold, whereas the increase with PsA was almost four-fold. Alternatively, the set of genes whose expression is affected as a direct or indirect result of the treatment with either PsA or VPA may be different and this could influence the developmental potential of the treated embryos. Indeed, working with a human promyelocytic leukemia cell line, Halsall et al. (2012) have demonstrated that different HDACis alter the expression of different subsets of genes. It would be interesting to investigate whether the same applies for cloned embryos treated with different HDACis.
Although the low efficiency of SCNT is mostly attributed to faulty nuclear reprogramming, improvement of technical aspects of the SCNT procedure can also result in significant increases in the success rate, as recently proved by Terashita et al. (2012, 2013) with the use of LatA instead of CB for actin polymerization inhibition.
In our last set of experiments, we decided to incorporate this modification to the SCNT protocol and test it in a group of PsA-treated cloned embryos. We obtained an almost three-fold increase in the offspring rate compared with the PsA-treated group in which CB was used. Even though the cause of this improved development was not investigated in the present study, because it falls beyond its aims, Terashita et al. (2012, 2013) reported that abnormal F-actin localization in cloned embryos can be corrected by LatA treatment, and that this results in a decreased incidence of abnormal chromosome segregation and increased full-term development. On the other hand, given that actin is also present in the nucleus (Baarlink and Grosse, 2014) and that nuclear actin polymerization seems to be fundamental in the transcriptional reactivation of Oct4 after transfer of mouse somatic nuclei into Xenopus oocytes (Miyamoto et al., 2011), the effects of LatA on nuclear actin polymerization in addition to cytoplasmic actin polymerization cannot be ruled out. Further studies should be performed to elucidate the exact mechanism by which LatA improves cloning efficiency.
In general, cloning efficiencies obtained in the present work were lower than those reported in other mouse cloning studies, but it must be pointed out that the mouse strain used in our study is the hybrid B6CBAF1, whereas the majority of previous studies have been performed with the hybrid B6D2F1 strain, and the effects of the mouse strain on SCNT efficiencies are well-known (Ogura et al., 2013).
In conclusion, we have corroborated our previous results on the positive effects of VPA on nuclear reprogramming, embryonic development, and quality of mouse cloned embryos (Costa-Borges et al., 2010) and demonstrated that 2 mM is the optimal concentration of this HDACi, even though the treatment duration can be extended from 8–9 h to 16 h. More importantly, we have successfully applied PsA, for the first time, in mouse SCNT procedures and we have shown that PsA improves mouse SCNT efficiency, even to a higher extent than VPA. Specifically, PsA increases embryo developmental rates, blastocyst quality (number of ICM cells), and nuclear reprogramming (histone acetylation) of the cloned embryos. Furthermore, when PsA is combined with the use of LatA instead of CB, it results in an 11.5-fold increase in full-term development. Further studies should be performed to analyze whether PsA has also beneficial effects in the generation of ESCs from the cloned embryos, in the cloning efficiency of other mouse strains and mammalian species, and in the nuclear reprogramming of somatic cells into induced pluripotent stem cells (iPSCs), as other epigenetic modifiers do.
Footnotes
Acknowledgments
We thank Jonatan Lucas and Ma Jesús Álvarez for their technical assistance and the staff at the Servei d'Estabulari of the Universitat Autònoma de Barcelona for taking care of the mice. This work was supported by the Spanish Ministerio de Ciencia e Innovación (AGL 2011-23784) and the Generalitat de Catalunya (2009 SGR 282). A.M. is a predoctoral fellow of the Universitat Autònoma de Barcelona.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.