
Abstract
Abstract
ID1 can mediate transforming growth factor-β (TGF-β)/activin receptor-like kinase-1 (ALK1)-induced (and Smad-dependent) migration in endothelial cells (ECs). However, the role that ID1 plays during differentiation of human embryonic stem cells (hESCs) into ECs induced by TGF-β1 remains unclear. In this study, a hESC differentiation model that recapitulates the developmental steps of vasculogenesis during the early stages of embryonic development was used to explore this question. We found that TGF-β1 increases endothelial cell differentiation and inhibits endothelial tube formation. Furthermore, at an early stage of differentiation, TGF-β1 may induce in vitro differentiation of hESCs into ECs by inhibiting expression of ID1, while at a later stage of differentiation, TGF-β1 may stimulate the proliferation and migration of ECs via the ALK1/Smad1/5/ID1 pathway. Downregulation of ID1 by gene silencing can lead to acceleration of TGF-β1–induced hESC differentiation into ECs and inhibition of proliferation and migration of ECs. This study may reveal some mechanisms of in vivo vasculogenesis in the early stages of embryonic development.
Introduction
B
Transforming growth factor-β1 (TGF-β1) is one of the most important cytokines that functions during vascularization. It has been shown to play a multifunctional role during nonsprouting angiogenesis (Goumans et al., 2002; Risau, 1997; Roberts et al., 1986). TGF-β1 transduces signals through its receptors, TGF-β type II receptor (TβR-II), TGF-β type I receptor (TβR-I), including activin receptor-like kinase-5 (ALK5), and activin receptor-like kinase-1 (ALK1), and their downstream effectors, such as the Smad family proteins, to regulate transcription of specific target genes controlling cell proliferation (Heldin et al., 1997). TGF-β1 can activate two distinct type I receptors (ALK5 and ALK1)/Smad signaling pathways with opposite effects (Levenberg et al., 2002; Kurisaki et al., 2003; Nonaka et al., 2008). The TGF-β/ALK5 pathway leads to inhibition of cell migration and proliferation, whereas the TGF-β/ALK1 pathway induces endothelial cell (EC) migration and proliferation. The ratio of TGF-β signal via ALK1 versus ALK5 determines whether TGF-β1 has a pro- or antiangiogenic effect (Goumans et al., 2002).
The inhibitor of differentiation/DNA binding-1 (ID1) protein can prevent basic helix–loop–helix (HLH) transcription factors from binding to DNA by its functional HLH structure (Norton, 2000). Research on the ID1 gene shows that it plays an essential role in the inhibition of the cell cycle and terminal cell differentiation in both normal and cancerous cells (Lin et al., 1999; Ryu et al., 2007; Schindl et al., 2003; Zheng et al., 2004). Inactivation of the ID1 protein by microinjection of anti-ID1 antibodies leads to the inhibition of G1–S progression and growth arrest in 3T3 cells (Peverali et al., 1994). Furthermore, primary embryo fibroblasts from ID1 knockout mice undergo premature senescence (Alani et al., 2001), whereas overexpression of ID1 results in the extension of life span in human keratinocytes (Alani et al., 1999; Nickoloff et al., 2000). ID1 overexpression has been observed in many types of human cancers, for example, prostate cancer (Ling et al., 2002). ID proteins have been reported to be required for the proliferative and invasive phenotype of ECs during angiogenesis (Lyden et al., 1999; Volpert et al., 2002), and ID1 can enhance proliferation, migration, and invasiveness of mouse mammary ECs (Lin et al., 2000; Sakurai et al., 2004). Recently, it was found that ID1 is a specific downstream target gene of ALK1 in ECs, and it can mediate the TGF-β/ALK1- induced (and Smad-dependent) migration (Goumans et al., 2002). However, it remains unclear what role ID1 plays during TGF-β1–induced differentiation of human embryonic stem cells (hESCs) into ECs.
hESCs, established from the inner cell mass of blastocysts (Thomson et al., 1998), are capable of differentiating into different cell types under various culture conditions in vitro, including hematopoietic precursors and ECs (Keller, 2005). They can provide a useful model system for studying the mechanisms involved in early differentiation and development of human embryonic blood vessels, because the progressive hematopoietic differentiation of hESCs through sequential hematoendothelial primitive and definitive stages resembles human yolk sac development (Gerecht-Nir et al., 2003; Lu et al., 2004; Zambidis et al., 2005).
In the current study, the in vitro model of hESC differentiation into ECs induced by TGF-β1 was established, and the expression and function of ID1 were analyzed during this process. We found that, at the early stages of differentiation, TGF-β1 may induce the in vitro differentiation of hESCs into ECs by inhibiting expression of ID1, whereas at later stages of differentiation TGF-β1 may stimulate the proliferation and migration of ECs via the ALK1/Smad1/5/ID1 pathway. However, TGF-β1 has a negative effect on angiogenic sprout formation during angiogenic stages. In addition, the in vitro differentiation of hESCs into ECs and TGF-β1–induced ID1 expression are not only time-dependent, but are also dependent on the concentration of TGF-β1. Downregulation of ID1 by gene silencing can lead to acceleration of TGF-β1–induced hESC differentiation into ECs and inhibition of proliferation and migration of ECs. Detection of the expression characteristics of signaling molecules of the TGF-β1 signaling pathway may help us to understand the mechanisms of how these signaling molecules function during the processes of differentiation and vasculogenesis.
Materials and Methods
hESCs culture and differentiation into the endothelial lineage
Undifferentiated hESCs (mSDU-hES 7 cell line, established by our lab; Huan et al., 2010) were grown on mitotically inactivated mouse embryonic fibroblasts (MEFs), cultured from fetuses of day-13.5 postcoital BALB/c mice by our lab) in knockout Dulbecco's Modified Eagle Medium (KO-DMEM) supplemented with 20% Knockout Serum Replacement (all from Invitrogen, Carlsbad, CA, USA), 0.1 mM β-mercaptoethanol (Roche, Nutley, NJ, USA), 0.1 mM nonessential amino acids (Invitrogen, Carlsbad, CA, USA), 1 mM
Small interfering RNAs preparation and cell transfection
Small interfering RNAs (siRNA, Invitrogen, Carlsbad, CA, USA) corresponding to ID1 mRNA were designed by Invitrogen. The pair of gene-specific sequences was 5′-UUUACCUGCUGCUCGUCCAGCAGGG-3′ and 5′-CCCUGCUGGACGAGCAGCAGGUAAA-3′. Transient transfection of siRNA into EBs or differentiated cells was performed with Lipofectamine™ 2000 reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's protocol. Transient transfections of only Lipofectamine™ 2000 without siRNA into EBs or differentiated cells were performed as negative controls.
Immunostaining of EBs and differentiated cells
EBs cultured in suspension were fixed overnight at 4°C in methanol-dimethyl sulfoxide (4:1), rehydrated in PBS for 30 min, and preincubated in PBS containing 2% bovine serum albumin (BSA) and 0.1% Tween 20 (PBT) for another 30 min. The attached EBs, and cells differentiated from the EBs, were fixed in 4% paraformaldehyde for 10 min, and then permeabilized with 0.1% Triton X-100 in PBS for 5 min. After treatment with 1% normal goat serum for 30 min at room temperature, the EBs and cells were incubated with rabbit anti-human platelet/endothelial cell adhesion molecule (PECAM1)/CD31 antibody (1:500; Boster, Wuhan, China) at 4°C overnight. EBs and cells were washed with PBT or PBS and then incubated with a fluorescent isothiocyanate (FITC)-conjugated goat anti-rabbit immunoglobulin G (IgG) secondary antibody (0.1 mg/mL; Sino-American Biotechnology, Luoyang, China) for 1 h. After washing with PBT or PBS, the stained slides were mounted with 0.5 μg/mL 4′,6-diamidino-2-phenylindole (DAPI; Beyotime, Haimen, China). The results were observed using a fluorescence microscope (Olympus, New York).
ECs enrichment with anti-PECAM1 antibodies
Because PECAM1 is abundantly expressed in ECs (Li et al., 2005; Schimmenti et al., 1992), it might be an ideal antigen candidate to detect ECs from the cell suspensions obtaining pluripotent, mesodermal, vascular-committed precursors, endothelial progenitors, and committed ECs using immunomagnetic beads coated with anti-PECAM1 antibodies using the manufacturer's protocol (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany). First, the cell suspension was diluted with buffer [0.5% fetal calf serum, 2 mM ethylenediamine tetraacetic acid (EDTA) in PBS, pH 7.2] and incubated with the anti-PECAM1 antibody-loaded microbeads for 30 min at 2–8°C, then washed with buffer solution. The cell suspension was transferred to a column placed in the magnetic field of a magnetic-activated cell-sorting (MACS) Separator (Miltenyi Biotec, Bergisch Gladbach, Germany). This allowed separation of the magnetically labeled PECAM1+ cells that were retained on the column while the unlabeled (PECAM1−) cells ran through the column. The column was then removed from the magnetic field and the PECAM1+ cells were eluted and collected. Then total RNA from the selected PECAM1+ cells was isolated using TRIzol reagent according to the manufacturer's protocol (TaKaRa, Dalian, China).
Western blot analysis
Cells were harvested and lysed with RIPA buffer [PBS, 1% Nonidet P-40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate (SDS), 10 μg/mL phenylmethylsulfonyl fluoride (PMSF), 20 μg/mL aprotinin, 10 μg/mL leupeptin, and 100 mM sodium orthovanadate] for protein extraction. The concentration of each protein extracted was measured using the Protein Quantitative Assay Kit (Shenergy Biocolor, Shanghai, China). Fifty micrograms of each cell lysate was heated (5 min, 95°C) and loaded on 5–10% Tris-HCl Criterion gels (Bio-Rad, Hercules, CA, USA), separated by SDS-polyacrylamide gel electrophoresis (PAGE) and then transferred to polyvinylidene fluoride (PVDF) membranes. The membranes were blocked with 10% nonfat milk, washed with TBST (20 mM Tris–HCl, 137 mM NaCl, 0.1% Tween 20), and then incubated with a monoclonal rabbit anti-human PECAM1 antibody (1:400; Boster, Wuhan, China), monoclonal rabbit anti-human phosphorylated Smad1/5 antibody (1:1000; Cell Signaling, Danvers, MA, USA), and monoclonal rabbit anti-human β-actin antibody (1: 5000; Jingmei Genevale Technology, Beijing, China). The membranes were then washed with TBST and reacted with horseradish peroxidase–conjugated secondary antibodies (1:10,000; Jingmei Genevale Technology, Beijing, China), and visualized by enhanced chemiluminescence (ECL) according to the manufacturer's recommendations (Millipore, Billerica, MA, USA).
qRT-PCR analysis of hEB cells, differentiated cells, and selected endothelial cells
Total RNA from EBs or from differentiated cells of purified ECs was isolated using TRIzol reagent according to the manufacturer's protocol (TaKaRa, Dalian, China). First-strand cDNA was generated using 0.5 μg of RNA, Superscript Reverse Transcriptase (Promega, Madison, WI, USA), random hexamer primers (Promega, Madison, WI), and oligo(dT) (Promega, Madison, WI). Gene-specific primers were designed with Primer3 software (Whitehead Institute/MIT Center for Genome Research), as shown in Table 1. PCR reactions were carried out by a 7500 Sequence Detection System (Applied Biosystems, Foster, CA) using a SYBR Green qPCR Kit (Applied Biosystems, Foster, CA, USA) according to the manufacturer's instructions.
The content of selected genes was normalized to the content of GAPDH, and a nontemplate control was performed for every gene as the blank control. Standard curves were generated using 10 ng of cDNA per 20-μL reaction volume. Samples were heated for 2 min at 50°C and 10 min at 95°C, and then subjected to 41 cycles of denaturation at 95°C for 15 sec and annealing and elongation at 60°C for 1 min. Finally, at the dissociation stage, samples were denatured at 95°C for 15 sec and annealed at 60°C for 15 sec. All PCR products were checked by melting curve analysis to exclude the possibility of multiple products or products of incorrect size. PCR analyses were conducted in triplicate for each sample. The level of target gene expression was determined by the comparative cycle threshold (Ct) method, whereby the target is normalized to the endogenous reference gene GAPDH. The Ct value is the cycle number at which the fluorescence level reaches the threshold. The ΔCt is determined by subtracting the Ct of the GAPDH control from Ct of target gene ΔCt=Ct (target)−Ct (GAPDH)]. This relative value of target to endogenous reference is described as the fold of GAPDH=2−ΔCt.
Statistical analysis
An unpaired Student's t-test and chi-squared analysis were performed for statistical analysis by using the statistical software SPSS13. Differences were considered significant when p<0.05.
Results
TGF-β1 enhances hESC differentiation into ECs and promotes proliferation of differentiated ECs from hESCs, but inhibits endothelial tube formation
To study the effects of TGF-β1 during differentiation of hESCs into ECs, and during the proliferation and migration of the differentiated ECs, different concentrations of TGF-β1 (0, 2, and 10 ng/mL) were added into the differentiation medium every other day from day 3 to day 14 of hESC differentiation. Platelet/EC adhesion molecule (PECAM1, CD31 molecule) was used as a marker of ECs (Schimmenti et al., 1992; Li et al., 2005).
In the presence of 0, 2, and 10 ng/mL TGF-β1, the percentage of 7-day-old EBs containing PECAM1-positive cells, as detected by immunostaining, was 70.50%, 81.60%, and 88.10%, respectively, in one representative experiment. The percentage of labeling in the 10 ng/mL TGF-β1 group was significantly higher than in the non-TGF-β1 group. This result demonstrated that TGF-β1 treatment increased the number of vascular EBs (p<0.05) (Fig. 1I).

In the model system of hESCs differentiating into ECs, TGF-β1 can enhance differentiation and promote vasculogenesis, while also inhibiting angiogenic sprout formation. (
Representative images show the expression of PECAM1 in control (CTL) and TGF-β1–treated (2and 10 ng/mL) EBs, and on differentiated cells, on days 7, 10 and 14 (Fig. 1II). During the process of hESCs differentiating into ECs (day 7) and during the early stages of adherent culture (day 10), TGF-β1 enhances the production of PECAM1-positive cells. In the presence of a high concentration of TGF-β1 (10 ng/mL), the enhancement is more prominent (Fig. 1II, A1, B1, C1, A2, B2, C2). On the contrary, results show that during the later stages of adherent culture (day 14), TGF-β1 inhibits the formation of a vascular-like structure from differentiated ECs, which is also concentration dependent, with high concentrations of TGF-β1 exerting a more inhibitory role (Fig. 1II, A3, B3, C3). The effect of TGF-β1 on angiogenic sprouting by measuring the length of PECAM1-positive sprouts was also studied (Fig. 1III). Quantitative analysis revealed that the addition of TGF-β1 reduced, in a dose-dependent manner, the length of angiogenic sprouts induced by angiogenic growth factors (p<0.05). Addition of TGF-β1 (10 ng/mL) had a dramatic negative effect on endothelial sprouting. This result demonstrates that, in the model of the hESCs differentiating into ECs, TGF-β1 can enhance differentiation and promote vasculogenesis, while also inhibiting angiogenesis.
TGF-β1 inhibits the expression of the ID1 gene during hESCs differentiation into ECs while enhancing the expression of the ID1 gene during the phase of vasculogenesis
To understand the response of the downstream gene ID1 to TGF-β1 during stages of hESC differentiation into ECs, and during proliferation of the differentiated ECs, 0 and 10 ng/mL TGF-β1 was added into the differentiation medium every other day from days 3 to 12 of EB formation.
The expression of the ID1 gene in the TGF-β1 (10 ng/mL)-treated group was compared to the control group on days 1, 4, 7, and 12 of differentiation, respectively. The expression of the ID1 gene is upregulated during all of the differentiation stages, in contrast to the initial undifferentiated state, whether induced by TGF-β1 or not (Fig. 2A). At an early differentiation stage on day 4, expression of ID1 was slightly lower in TGF-β1–treated group. As differentiation proceeded on day 7, TGF-β1 exerted a more inhibitory effect on the expression of ID1, and the disparity of expression between the two groups was much greater. From day 8, cells grew out from EBs, were adherent to the dishes, and formed a single-layer structure before entering the proliferation stage. However, during the vasculogenesis phase on day 12, expression of ID1 in the TGF-β1–treated group was greater than in the control group.

TGF-β1 induces hESCs to differentiate toward the endothelial lineage by downregulating expression of ID1, but promotes proliferation of differentiated ECs by upregulation of ID1 expression. Quantitative real-time PCR analysis of mRNA expression of ID1 (
Expression of PECAM1 and KDR (kinase insert domain receptor, the type II VEGF receptor) mRNA was detected simultaneously as an indicator of the substrate change of the ECs (Fig. 2B, C). To confirm that the PECAM1-positive cells being studied corresponded to ECs, the expression of KDR mRNA, one of the earliest markers of developing ECs, was also analyzed (Hirashima et al., 1999). Expression of PECAM1 and KDR increased continuously along with differentiation and proliferation. Furthermore, expression of these two marker genes in the TGF-β1–treated group was almost higher than in the control group, which confirmed that TGF-β1 could enhance the production of PECAM1-positive cells during differentiation and vasculogenesis.
Kinetics of ID1 expression during differentiation and vasculogenesis induced by TGF-β1
The expression tendency of the ID1 gene during differentiation and vasculogenesis was studied next. TGF-β1 (10 ng/mL) was added every other day to the differentiation medium from day 3 to day 12. Expression of the ID1 gene in multiple intermediates containing pluripotent, mesodermal, vascular-committed precursors, endothelial progenitors, and committed ECs at the population level was detected on days 1, 4, 5, 6, 8, 10, and 12 (Fig. 3A). At the differentiation stages (days 4, 5, 6), a lower level of ID1 was expressed. When it came to the vasculogenesis stages (days 8, 10, 12), expression of ID1 increased dramatically and maintained a higher level. Expression of PECAM1 and KDR mRNA was also detected simultaneously as the indication of the change in EC substrate (Fig. 3C, D), both of which increased continuously along with TGF-β1 induced differentiation and vasculogenesis. Expression of the ID1 gene in purified ECs was also detected on days 1, 4, 5, 6, 8, 10, and 12 (Fig. 3B). The result shows that the expression tendency of the ID1 gene in purified ECs was slightly similar to that in multiple intermediates, although some differences exist. At the differentiation stages (days 4, 5, 6), a lower level of ID1 was expressed. When it came to the vasculogenesis stages (days 8, 10, 12), expression of ID1 increased more dramatically.
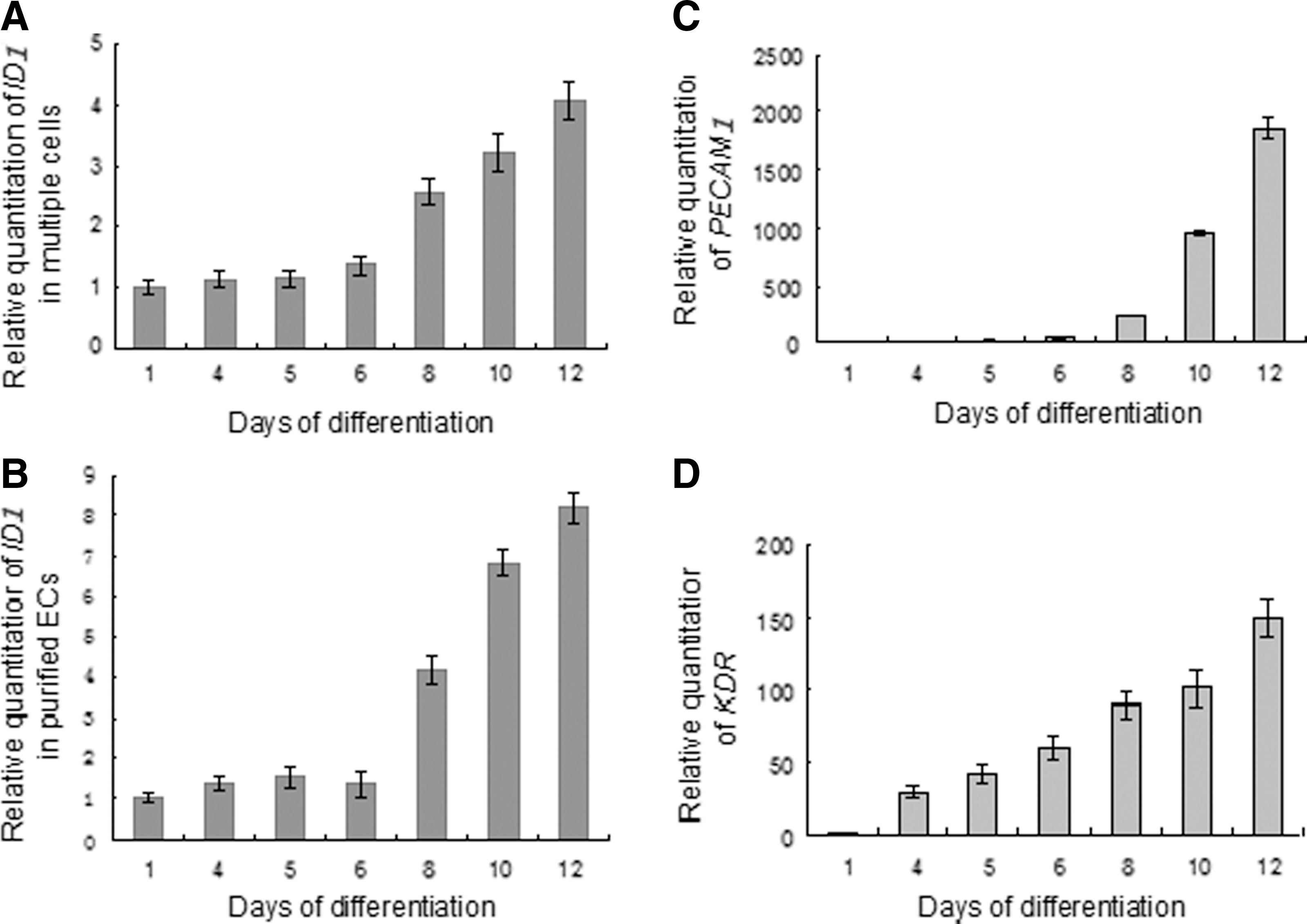
Quantitative real-time PCR analysis of ID1 expression in hESC-derived EBs, multiple cells, and purified ECs. RNA was extracted from differentiated hESCs (from days 1 to 12), and analyzed for the expression of ID1 (
Interfering with ID1 expression leads to promotion of hESC differentiation into ECs, but leads to inhibition of vasculogenesis
The function of the ID1 gene during differentiation and vasculogenesis was further studied by knocking down its expression. Small interfering (si) RNA was transfected into EBs and differentiated cells using Lipofectamine™ 2000 every other day to interfere with gene expression of ID1. The interfering effects were confirmed not only in multiple intermediates (Fig. 4 A1, B1, C1) but also in purified PECAM1+ ECs (Fig. 4 A2, B2, C2). On day 4, ID1 was successfully destroyed in the test group but not in the two control groups, whereas expression of PECAM1 mRNA and PECAM1 protein increased in the test group (Fig. 4A). On day 7 of differentiation, this phenomenon was more prominent (Fig. 4B). When it came to the vasculogenesis stage, on day 12, interfering with expression of ID1 led to downregulation of PECAM1 mRNA and PECAM1 protein (Fig. 4C). These data revealed that downregulation of ID1 expression could lead to promotion of hESCs differentiation into ECs, but also inhibition of the proliferation of ECs that had differentiated from hESCs.

Downregulation of ID1 expression could lead to promotion of hESCs differentiation into ECs, but inhibition of proliferation of the differentiated ECs. siRNA was transfected into EBs and differentiated cells by Lipofectamine™ 2000 every other day to interfere with expression of ID1 (SRI). Transient transfection of only Lipofectamine™ 2000 without siRNA was performed as a negative control (NTC) and neither siRNA nor lipofectamine™ 2000 transfected group as a blank control (BLC). On days 4 (
Kinetics of TGF-β1 receptors and some signal proteins expression during differentiation and vasculogenesis stages
Finally, the expression of TGF-β1 receptors and some signaling proteins in the TGF-β/ALK signal transduction pathway were studied during stages of differentiation and vasculogenesis from day 0 to day 12 (Fig. 5). TGF-β1 regulates cellular processes by first binding to a type II serine/threonine kinase receptor, TβR-II (Massague, 1998). TβR-II was not expressed in undifferentiated hESCs and appeared after 2 days of differentiation. It increased to a higher level on day 6 and was maintained to day 12 (Fig. 5A). TβR-I ALK5 was present both in undifferentiated hESCs and throughout the differentiation and proliferation processes (Fig. 5B). In contrast, the other TβR-I, ALK1, was not expressed in undifferentiated hESCs. Its expression began on day 6 of differentiation and rapidly increased to a high lever on day 10 (Fig. 5C). TGF-β1 induces phosphorylation of Smad1/5 and Smad2 in ECs, and these effects can be blocked upon selective inhibition of ALK1 or ALK5 expression, respectively (Goumans et al., 2002). We detected protein expression of the downstream signaling molecules of ALK1 or ALK5, respectively, phosphorylated Smad1/5 and Smad2. The presence of phosphorylated Smad2 declined (Fig. 5D), whereas phosphorylated Smad1/5 increased (Fig. 5E) along with the process of differentiation and vasculogenesis.
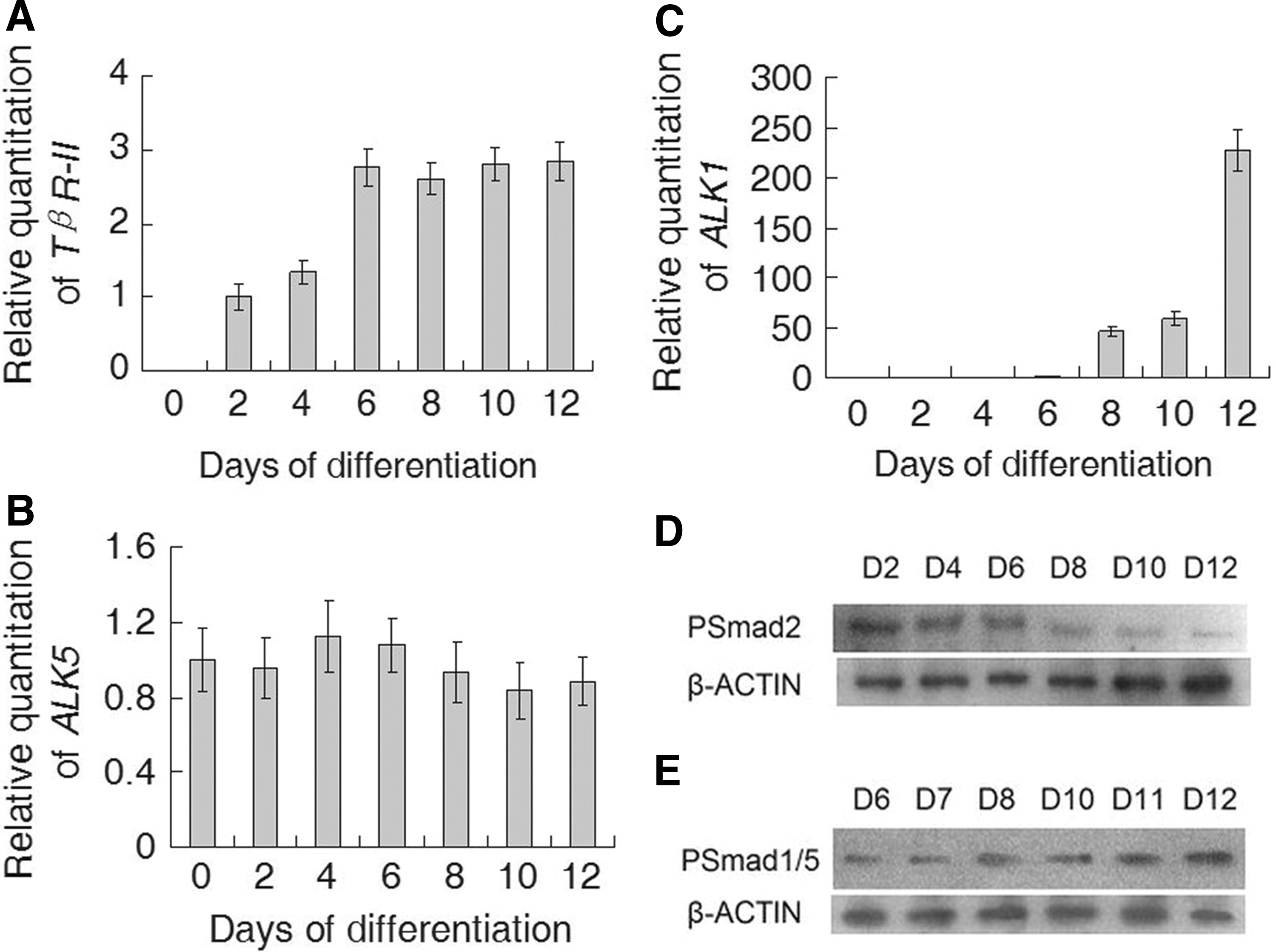
Kinetics of the expression of TGF-β1 receptor mRNA and specific signaling protein mRNA during stages of differentiation and vasculogenesis. RNA was extracted from undifferentiated hESCs (day 0) and differentiated EBs and ECs (from days 2 to 12) and analyzed for the expression of TβR-II (
Discussion
The main aim of this study was to identify the role that ID1 plays during the in vitro differentiation of hESCs into ECs induced by TGF-β1. We found that in this model at the early stage of differentiation TGF-β1 is an inducer of EC differentiation and may induce the in vitro differentiation of hESCs into ECs by inhibiting expression of ID1. At the late stage of differentiation, TGF-β1 may promote the proliferation and migration of the differentiated ECs via the ALK1/Smad1/5/ID1 pathway and the ID1 gene was upregulated and acted a positive role. We also demonstrated that TGF-β1 has a negative effect on the angiogenic sprout formation during angiogenesis stage. During in vitro differentiation of hESCs into ECs induced by TGF-β1, regulation of ID1 are both time and concentration dependent on TGF-β1. Downregulation of ID1 by siRNA interference of this gene could accelerate hESCs differentiation into ECs induced by TGF-β1 but weakens the proliferation and migration of ECs.
In our present work, we analyzed qualitatively the effect of TGF-β1 with different concentrations on differentiation of hESCs into ECs and the proliferation of differentiated ECs. PECAM1 was used as the marker of differentiated ECs. Although its expression can be observed on some non-ECs at this differentiating stage, for example, hematopoietic precursors, PECAM1 has been proven to be a valuable tool for identifying ECs at this stage (Cho et al., 2007; Ferreira et al., 2007). The results showed that TGF-β1 could enhance the production of PECAM1-positive cells, and this effect was positively related to its concentration. This coincides with some in vitro and in vivo studies reported previously. It has been demonstrated that latent TGFβ-binding protein-1 promotes in vitro differentiation of mouse ESCs toward the endothelial lineage (Gualandris et al., 2000). Accordingly, TGF-β1–deficient mice show defective vasculogenesis due to a defect in endothelial differentiation (Dickson et al., 1995). However, during the angiogenic sprouting stage of hESC-differentiated ECs, the effect of TGF-β1 was completely reversed in a concentration-dependent manner. Therefore, it is difficult to draw conclusions about the complete effect of TGF-β1 on the differentiation of hESCs toward the endothelial lineage because it will have different functions on vessel formation at different stages of the angiogenic process.
Further work will be required to identify how TGF-β1 regulates one of its downstream genes, ID1, during the process of hESCs differentiation toward the endothelial lineage and also during the proliferation stage of the differentiated ECs. We discovered that expression of the ID1 gene was downregulated during the differentiation stages but upregulated during the stages of proliferation. Research shows that the ID1 gene is required for inhibition of the cell cycle and terminal cell differentiation (Alani et al., 2001; Nickoloff et al., 2000; Peverali et al., 1994). Downregulation of the Id protein is necessary for terminal differentiation in many developmental processes, including angiogenesis and tumor vascularization (Norton, 2000). We have shown that TGF-β1 could promote the production of PECAM1-positive cells during differentiation; thus, it may be concluded that during the TGF-β1–induced differentiation of hESCs toward the endothelial lineage, TGF-β1 may exert a positive effect on endothelial-oriented differentiation by inhibiting expression of the ID1 gene.
On the contrary, expression of the ID1 gene was upregulated during the stages of proliferation. Id proteins have been reported to play an essential role in the proliferative and invasive phenotype of ECs during angiogenesis (Lyden et al., 1999), and Id1 can enhance proliferation, migration, and invasiveness of mouse mammary ECs (Lin et al., 2000). Id1 has proven to serve as an effector for the TGF-β/ALK1 pathway in mediating a stimulatory effect on proliferation during mouse ESC differentiation (Goumans et al., 2002). In our study, we discovered that TGF-β1 may exert its stimulatory effect via upregulation of the ID1 gene expression during the TGF-β1–induced proliferation stage of EC differentiation.
To understand better the function of the ID1 gene on vasculogenesis and differentiation, this gene was knocked down by siRNA interference technology. These experiments demonstrated that downregulation of ID1 expression may promote hESCs differentiation into ECs while also inhibiting proliferation of the ECs that had differentiated from hESCs. This confirmed the conclusion that ID1 serves as an effector for the TGF-β1 pathway in mediating an inhibitory effect on differentiation but a stimulatory effect on proliferation of ECs.
To understand better the detailed mechanism of the pathway mediating the TGF-β1–induced endothelial lineage differentiation from hESCs and its proliferation, possible pathway members participating in these processes were detected. Because TβR-II appeared after 2 days of differentiation and remained at a high level from days 6 to 12, it may play roles during the entire time of differentiation and proliferation. Expression of one TβR-I receptor, ALK1, began during day 6 of differentiation and rapidly increased to higher levels on day 10, while the other TβR-I, ALK5, was expressed in both undifferentiated hESCs and throughout the differentiation and proliferation processes. Meanwhile, ALK1 expression followed PECAM1 and KDR expression. These results suggested that the provasculogenic effect of TGF-β1 on undifferentiated hESCs can probably be attributed to ALK5, whereas ALK1 is specifically expressed in differentiated ECs.
Analysis of ALK1- and ALK5-regulated genes suggested that ALK5 might be more important than ALK1 in the modulation of extracellular matrix and differentiation of periendothelial cells, whereas ALK1 may be more involved in vascular maturation through activation of intracellular regulators (Mallet et al., 2006). Both ALK1 and ALK5 are functional TβR-I in endothelial cells. The activation state of the endothelium may be dependent on the balance of ALK1 versus ALK5 activation induced by TGF-β1 (Goumans et al., 2002).
Our results also support the hypothesis that members of the TGF-β/ALK1 pathway play a pivotal role in mediating proliferation of differentiated ECs. Smads are key components in these signal transduction pathways. Smad1, Smad2, and possibly Smad5 interact with and become phosphorylated by specific type I serine/threonine kinase receptors and thereby act in a pathway-restricted fashion (Heldin et al., 1997). Examination of the relative levels of phosphorylated Smad1/5 and Smad2 demonstrated that phosphorylated Smad2 could function primarily during the differentiation stage, while phosphorylated Smad1/5 may act mainly during proliferation. It is established that TGF-β1/ALK1 activates the ID1 transcriptional reporter, which is a Smad-dependent response (Korchynskyi and ten Dijke, 2002). ALK1 signals through Smad1/5, whereas ALK5 signals through Smad2/3; each receptor induces a different set of genes with some overlap (Mallet et al., 2006). Combined with the expression kinetics of the ID1 gene, we demonstrated that TGF-β1 may stimulate differentiation toward the endothelial lineage by an activated ALK5 pathway, whereas during the proliferation process TGF-β1 may enhance the proliferation of differentiated ECs through the activated TGF-β1/ALK1/ID1, Smad1/5-dependent pathway.
Thus far, the participation of the ID1 gene in the sprouting process of angiogenesis has not been reported. Instead, it has been demonstrated that ALK5 signal transduction contributes to basement membrane formation in ECs, supporting a role for ALK5/Smad signaling in the maturation of blood vessels during the resolution phase (Goumans et al., 2002). Therefore, further studies of this laboratory will focus on identifying whether the ID1 gene is involved in the process of endothelial tube formation. This may help us to understand the complete function of the ID1 gene during the entire process of vascular formation.
Footnotes
Acknowledgments
We thank all of the staff in the central laboratory of Shandong Provincial Hospital for providing equipment and technical support for this study.
This work was supported by the National Natural Science Foundation of China (81200427), National Key Technology Research and Development Program of China (2011BAI17B00), Science Research Foundation Item of No-Earnings Health Vocation of China (201002013).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.