
Abstract
Abstract
Since human embryonic stem cells (hESCs) were first isolated and successfully cultured in vitro, the pluripotent potential of hESCs has been underestimated. The pluripotency of mouse embryonic stem cells (mESCs) can be categorized as naïve and primed, depending on their corresponding in vivo developing phases. mESC morphology differs at distinct pluripotent states, which differ in signaling dependence, gene expression, epigenetic features, and developmental potential. hESCs resemble mouse stem cells at primed pluripotency, and consequently are believed to correspond to a later developmental stage in vivo than mESCs. Nevertheless, recent studies indicate that a naïve state of pluripotency may exist in hESCs, and the pluripotency of hESCs also can be enhanced by genetic modification or optimized culture systems. These findings provide novel insight into the properties and differentiation potential of hESCs. Here, we review the recent advances in characterization of ESC states and investigate the mechanisms regulating hESC pluripotency.
Introduction
T
Distinct Features of mESCs at Different Pluripotent States
Naïve mESCs are derived from preimplanted ICM (Brook and Gardner, 1997; Evans and Kaufman, 1981; Martin, 1981), whereas primed mEpiSCs were separated from the postimplanted epiblast of egg cylinders (Brons et al., 2007; Tesar et al., 2007), and their features respectively correspond to in vivo pre- and postimplanted embryos, respectively (Fig. 1) (Brons et al., 2007; Buecker and Geijsen, 2010; Han et al., 2010; Rossant, 2008; Tesar et al., 2007). Naïve mESCs typically form colonies that can be dissociated by trypsin and survive as single cells; however, flattened mEpiSCs do not tolerate trypsin treatment, and single-cell cloning efficiency is low (Brons et al., 2007; Tesar et al., 2007). While leukemia inhibitory factor (LIF) is required to maintain the undifferentiated growth of naïve mESCs, basic fibroblast growth factor (bFGF) is the classic supplemental factor for mEpiSC culture (Brons et al., 2007; Tesar et al., 2007). The fundamental distinction between naïve and primed mESCs is their developmental potential. Only mESCs can efficiently contribute to blastocyst chimeras, whereas mEpiSCs have the potential to differentiate into primordial germ cells in vitro (Guo et al., 2009; Hayashi and Surani, 2009; Nichols and Smith, 2009; Tesar et al., 2007).
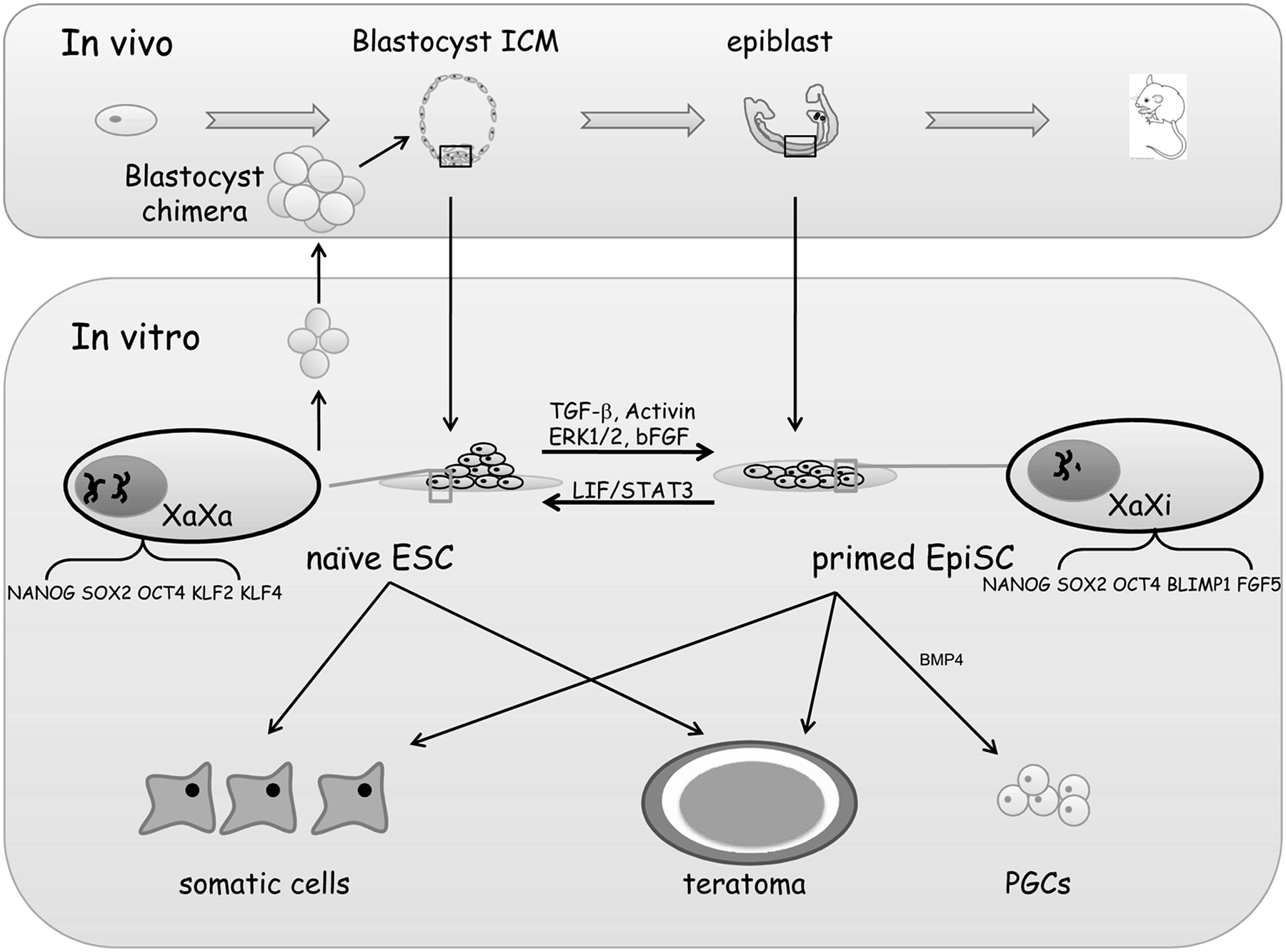
Comparision of properties of mESCs and mEpiSCs.
These differences can be attributed to different epigenetic modifications and alterations in gene expression. For example, FGF5, T (brachyury), and Lefty are expressed in mEpiSCs, whereas STELLA (Dppa3) and REX1 (Zfp42) are expressed only in naïve mESCs. Other genes, such as NANOG and KLF4, are expressed in both primed and naïve cells, however at significantly different levels (Bao et al., 2009; Guo et al., 2009). Interestingly, under specific conditions, both epigenetic and genetic features could be swapped, and consequently naïve cells can be converted into primed pluripotent cells and vice versa (Bao et al., 2009; Guo et al., 2009; Silva et al., 2009). Several specific in vitro reprogramming methods have been previously reported, potentially as a result of the range of different culturing systems applied in different laboratories (Bao et al., 2009; Guo et al., 2009).
Guo et al. have successfully converted mEpiSCs to the naïve pluripotent state by introducing a single reprogramming factor, Krüppel-like factor 4 (KLF4) to the 2i/LIF culturing conditions (culture in the presence of two inhibitors and LIF) (Guo et al., 2009). The original epigenetic features were erased and the converted cells were stably reprogrammed, as their properties were maintained after deleting the exogenous KLF4 gene. These converted cells regained the capacity to produce chimeras that yielded germ-line transmission, hence demonstrating that primed cells possess the potential for naïve pluripotency.
Furthermore, Bao et al. reported that both freshly isolated and cultured epiblast cells could overcome the epigenetic barrier, altering the transcriptome in response to LIF-STAT3 signaling without exogenous transcription factors, resulting in stable reprogramming to ES-cell-like cells (rESCs). Unlike mEpiSCs, DNA demethylation, X reactivation, and expression of E-cadherin occurred in rESCs. Moreover, rESCs were capable of contributing to somatic tissues and germ cells in chimeras, which is not possible with mEpiSCs (Bao et al., 2009).
In our previous work employing human amnion epithelial cells as the feeder layer, we have successfully converted one primed mESC line into naïve-like cells without genetic modification (Chen et al., 2013). During this process, genetic and epigenetic features were reprogrammed, and different culture supplements were required to maintain undifferentiated growth. Thus, we described a method by which primed mESCs can be converted into naïve pluripotent cells.
hESC Pluripotency
As with mESCs, hESCs are also derived from the preimplanted ICM (Thomson et al., 1998). However, canonic hESCs were believed to be at the primed pluripotent state, resembling primed mEpiSCs in morphology, developmental potency, gene expression, and epigenetic modification (Buecker and Geijsen, 2010; Hanna et al., 2010; Kerr and Cheng, 2010; Tesar et al., 2007; Thomson et al., 1998). Mouse and human induced pluripotent stem cells (iPSCs), obtained by introducing similar transcription factors into somatic cells also exhibit these characteristics (Takahashi and Yamanaka, 2006; Takahashi et al., 2007).
The undifferentiated growth of canonic hESCs and human iPSCs depends on bFGF and transforming growth factor-β (TGF-β) signaling. Flattened colonies of those cells are sensitive to trypsin treatment, and they grow more slowly than naïve mESCs (Buecker and Geijsen, 2010; Nichols and Smith, 2009). Thus hESCs are typically split by mechanical means or by collagenase (Yu and Thomson, 2008). Although the core pluripotent factors OCT4, SOX2, and NANOG are expressed in hESCs, mEpiSCs, and mESCs, gene expression profiling has indicated that hESCs exhibit an expression pattern more similar to primed mEpiSCs than naïve mESCs (Bao et al., 2009; Brons et al., 2007; Guo et al., 2009; Tesar et al., 2007).
OCT4 is a critical transcriptional factor for stemness, and a hallmark of certain kinds of stem cells (Schöler et al., 1990). Despite the uniform expression of OCT4 in mESCs and hESCs, in naïve cells the transcription of OCT4 is driven by the distal enhancer, whereas the proximal enhancer is used selectively in primed mEpiSCs and hESCs (Gafni et al., 2013; Chia et al., 2010; Hanna et al., 2010; Tesar et al., 2007).
X-chromosome inactivation (XCI), which occurs in one of the two X chromosomes in female cells, is an important mechanism of dosage compensation during embryonic development and is regarded as a feature of primed female ESCs. In most female hESCs, XCI exists in the XaXi state (one active X chromosome) (Silva et al., 2008). Lengner et al. derived and established XaXa (two active X chromosomes) hESCs under physiological oxygen concentrations (∼5%, pO2, 36 mmHg), and demonstrated that exposure to atmospheric oxygen (∼20%, pO2, 142 mmHg) alone is sufficient to initiate irreversible XCI (Lengner et al., 2010).
Other aspects of epigenetic modification have also been reported to differ between mESC and hESC. In comparison to mESCs, the DNA methylation of hESCs is globally enhanced, and the repressive H3K27me3 modification is detected in hESC naïve pluripotent genes (Hanna et al., 2009). Therefore, hESCs were presumed to adopt only primed pluripotent state; however, this conclusion has been recently challenged by studies revealing novel pluripotent states of hESCs (Gafni et al., 2013; Buecker et al., 2010; Hanna et al., 2010; Lengner et al., 2010; Xu et al., 2010).
Reprogramming of Primed hESCs
Novel stem cells, termed hLR5 iPSCs, were obtained by ectopically expressing five reprogramming factors (OCT4, SOX2, KLF4, c-MYC, and NANOG) in human iPSCs in the presence of LIF. hLR5 iPSCs adopted similar morphological, molecular, and functional properties to naïve mESCs (Buecker et al., 2010; Kerr and Cheng, 2010); however, it should be noted that maintenance of hLR5 iPSCs required continuous expression of those five reprogramming factors.
Hanna et al. produced naïve-like hESCs from human iPSCs by ectopically expressing OCT4, KLF4, and KLF2 in the presence of small molecule inhibitors against glycogen synthase kinase 3β (GSK3β) and stimulation of the mitogen-activated protein kinase pathway (ERK1/2) (Hanna et al., 2010). These converted cells resembled naïve mESCs in terms of growth, gene expression profile, signaling dependence, and X-chromosome activation state (XaXa). Furthermore, in the presence of an agonist of the protein kinase A pathway, forskolin, reprogramming occurred in the absence of the previously described ectopic factors. However forskolin-dependent naïve hESCs could not be propagated for more than 15–20 passages, whereas the ectopically modified naïve hESCs could be maintained for more than 50 passages. The limited capacity for propagation may due to the toxicity of forskolin, or its inability to fully substitute for ectopic transgene expression (Hanna et al., 2010).
As discussed above, with a combination of genetic modifications and molecular supplementation, hESCs could be reprogrammed into a naïve pluripotent state. Furthermore, recent studies have demonstrated that by altering culture conditions. hESCs could be interconverted between the two pluripotent states of (Gafni et al., 2013; Xu et al., 2010).
By transferring hESCs to mESC culture conditions, including inhibitors of mitogen-activated protein kinase/extracellular signal-regulated kinase (MEK) and p38, Xu et al. generated mESC-like hESCs (Xu et al., 2010). The undifferentiated growth of mESC-like hESCs was maintained with LIF. They formed small domed colonies, growing faster than the original cells, and were less vulnerable to trypsin dissociation. The converted hESCs also maintained pluripotent features—they expressed marker genes and developed into all three germ layers in vitro (Xu et al., 2010).
The 2i/LIF culturing medium, which consists of LIF and small molecule inhibitors of ERK1/ERK2 and GSK3β signaling, can efficiently maintain mESCs, but not primed mEpiSCs and hESCs, in vitro (Hanna et al., 2010). Although primed hESCs rapidly differentiated in 2i/LIF, naïve pluripotent hESCs when supplemented with additional factors could be derived directly from blastocysts. Under these conditions, previously established primed hESCs or iPSCs also successfully adopted naïve pluripotency (Gafni et al., 2013). 2i/LIF, p38i, JNKi, FGF2, and TGF-β1 have been identified as essential supplementations for the conversion and maintenance of naïve pluripotent hESCs. Rho-associated coiled-coil kinases and protein kinase C inhibitors could further promote the viability and growth of naïve cells. Finally, Gafni et al. defined optimized conditions for naïve hESCs, termed naïve human stem cell medium (NHSM).
Naïve hESCs formed domed colonies, and their doubling time, single-cell cloning efficiency, and gene expression were distinct from the original hESCs, but similar to naïve mESCs. Naïve pluripotency-associated genes, such as NANOG and DUSP, were upregulated, whereas the expression of lineage-commitment genes, such as ZIC1, SOX6, and SOX11, was suppressed. The distal enhancer of OCT4 was activated, and a pre-X inactivation state was maintained in naïve hESCs. This finding demonstrated that an erasing reaction on epigenetic modification occurred during the conversion to pluripotency. The enriched nuclear transcription factor TFE3 is a novel marker of naïve mESCs, and enrichment of TFE3 in the nucleus was also detected in naïve hESCs in this study (Betschinger et al., 2013).
Recently, Theunissen et al. (2014) established a reporter system specific for naïve human pluripotency using transcription activator-like effector nuclease (TALEN)-based genome editing and performed an iterative chemical screen to identify a combination of five kinase inhibitors to define the culture requirements of naïve human pluripotent cells. These optimized conditions enabled both the interconversion between conventional and naïve human ESCs in the absence of reprogramming factors and the direct isolation of naïve ESCs from human blastocysts. To assess naïve human pluripotency, they also generated interspecies chimeras by injection of human ESCs into mouse morulae; however, it seems that this assay proved too inefficient to become employed routinely (Gafni et al., 2013).
Potential Mechanisms Regulating hESC Pluripotency
Although the mechanisms regulating naïve hESC pluripotency remain elusive, the establishment of naïve or naïve-like hESCs has provided the opportunity to examine the pathways regulating pluripotency in hESCs. OCT4, SOX2, and NANOG are three core hallmarks of ESC pluripotency, but expression is dependent on LIF or bFGF in naïve or primed pluripotent states, respectively. Among those transcription factors, only NANOG is a downstream target of both the LIF and bFGF pathways (Xu et al., 2008); consequently, NANOG is believed to be an important regulator of pluripotency (Buecker and Geijsen, 2010). NANOG is a homeodomain protein expressed in mammalian pluripotent cells and developing germ cells (Chambers et al., 2003; Mitsui et al., 2003). In mouse cells, low NANOG expression leads to a more primed pluripotent state (Buecker and Geijsen, 2010; Chambers et al., 2007), whereas its overexpression facilitates the reprogramming of primed mEpiSCs to naïve mESCs (Silva et al., 2009). Hanna et al. reported that in hESCs the upstream regulator KLF4 is required for reprogramming primed pluripotency to the naïve state (Hanna et al., 2010).
The FGFR and TGFR pathway play an important role during hESCs reprogramming from primed to naïve pluripotency (Gafni et al., 2013). Gafni et al. found that exogenous bFGF and TGF-β1 enhanced the conversion efficiency without exogenous genetic modification. It is interesting to note that bFGF and TGF-β have evolutionary divergent functions. In hESCs, bFGF and TGF-β induce expression of KLF4 and NANOG, which are naïve pluripotent transcription factors, but in murine EpiSCs they fail to actively promote self-renewal through NANOG (Greber et al., 2010). These signaling pathways are distinctly regulated during the early embryonic development of humans and mice (Hassani et al., 2014), likely contributing to the differences observed between naïve mESCs and primed hESCs.
In the mouse, LIF-stimulated STAT3 expression has been reported to be required for maintenance of naïve mESC pluripotency and viability (Nichols and Smith, 2009), and the LIF-STAT3 pathway represents a crucial mechanism by which primed mEpiSCs are reprogrammed to naïve mESCs (Bao et al., 2009). STAT3, a transcription factor activated by LIF, is weakly expressed in primed mEpiSCs (Nichols and Smith, 2009; Yang et al., 2010). Withdrawing LIF led to impaired conversion of mEpiSCs to rESCs; and, in contrast, constitutive expression of STAT3 restored the conversion of mEpiSCs to rESCs (Yang et al., 2010). Consistent with naïve mESCs, naïve hESC pluripotency also depends on the LIF-STAT3 pathway. Blocking the STAT3 pathway led to rapid differentiation of hESCs; furthermore, constitutively active STAT3 could maintain naïve hESCs without addition of LIF (Gafni et al., 2013), demonstrating that regulation of the LIF-STAT3 pathway in hESCs is similar to that in naïve mESCs (Sridharan et al., 2009; Guo et al., 2009; Silva et al., 2008).
The balance of E-cadherin and N-cadherin is also an important controller of pluripotency of stem cells (Chou et al., 2008). In mouse ESCs, E-cadherin is under the positive regulation of LIF-STAT3 signaling and is therefore expressed in naïve pluripotent cells, whereas N-cadherin is detected in primed pluripotent cells (Bao et al., 2009). In hESCs, E-cadherin signaling is essential for cell survival. After treatment with trypsin, most E-cadherin is degraded in both hESCs and mESCs. However, in mouse ESCs, newly synthesized E-cadherin is much more stable than in hESCs, in which the irreparable disruption of E-cadherin signaling causes cell death in response to trypsin exposure (Xu et al., 2010). In mESC-like hESCs, E-cadherin is more abundant and more stable than in original hESCs. Blocking E-cadherin significantly inhibited the attachment and proliferation of the mESC-like hESCs (Xu et al., 2010), suggesting that the E-cadherin/N-cadherin pathway may also regulate the different pluripotent states of hESCs.
KLF family proteins are evolutionarily conserved zinc finger transcription factors involved in many biological processes (McConnell et al., 2007). KLF circuitry has been reported to regulate key pluripotency genes and integrate into the NANOG transcriptional network, controlling ESC gene expression (Hanna et al., 2010; Guo et al., 2009; Hall et al., 2009; Hanna et al., 2009; Jiang et al., 2008). KLF4, one of the four transcription factors required for iPSC establishment, promotes self-renewal of mESCs (Li et al., 2005). KLF4 is normally downregulated during the differentiation from mESCs to mEpiSCs, and its re-expression could induce mEpiSCs to the naïve pluripotent state under appropriate conditions (Guo et al., 2009). In naïve mESCs, but not mEpiSCs, KLF4 could be induced by LIF-STAT3 signaling (Yang et al., 2010), and stimulating expression of KLF4 and KLF2 in hESCs by forskolin could replace ectopic transgene expression, thus reprogramming primed hESCs to a naïve state (Hanna et al., 2010).
In addition, LKB1 (serine/threonine kinase 11, STK11), an evolutionarily conserved kinase with multiple tumor suppressor functions, is essential for embryonic development in mice. The absence of LKB1-mediated signaling has been reported to be an important determinant of feeder cell-mediated support of hESC renewal and to contribute to the pluripotent state of hESCs in multiple culture systems (Lai et al., 2012).
As discussed above, several signaling pathways have been implicated in the maintenance and reprogramming of hESC pluripotency, but the overall picture is not well understood. Perhaps network crosstalk, rather than a single, clearly defined pathway, regulates and stabilizes the different pluripotent states in ESCs. Uncovering the details of these regulatory networks may illuminate the mechanisms regulating pluripotency in hESCs and facilitate definition of optimized culture conditions for naïve hESCs.
Perspective
Four main types of pluripotent cells have been obtained and established in vitro: Embryonal carcinoma cells (ECs) originating from teratocarcinomas (Hogan, 1976); ESCs derived from the ICM of developing blastocysts (Evans and Kaufman, 1981); embryonic germ cells (EGs) derived from primordial germ cells (Labosky et al., 1994); and EpiSCs derived from the postimplantation epiblast (Brons et al., 2007; Tesar et al., 2007). Among those cell types, hESCs represent a powerful tool for research on embryonic development, cell replacement therapy, and tissue engineering. However, the lack of hESC naïve pluripotency limits their application.
Immortalized naïve and primed ESCs are believed to correspond to distinct in vivo developmental stages and present distinct features in vitro. Studies about mESCs have proved that the naïve and primed pluripotent states could be interconverted in vitro (Bao et al., 2009; Guo et al., 2009; Silva et al., 2009). Recently, studies have also revealed that hESCs also could be maintained at the naïve pluripotency, which is meaningful for uncovering the human embryonic development mechanism and for understanding the characteristics of hESCs. Consequently, defining an ideal in vitro system to obtain naïve hESCs is critical for their clinical therapeutic application in cell transplantation and regenerative medicine and for constructing disease models in vitro.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (81070533 and 81370678), Shanghai Municipal Council for Science and Technology (12431902201 and 14YF1408200), Shanghai Municipal Health & Family Planning Commission (XBR2011069 and 20134Y128), and Shanghai Jiaotong University School of Medicine (13XJ10067).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.