
Abstract
Abstract
Faulty epigenetic reprogramming of somatic nuclei is thought to be the main reason for low cloning efficiency by somatic cell nuclear transfer (SCNT). Histone deacetylase inhibitors (HDACi), such as Scriptaid, improve developmental competence of SCNT embryos in several species. Another HDACi, Oxamflatin, is about 100 times more potent than Scriptaid in the ability to inhibit nuclear-specific HDACs. The present study determined the effects of Oxamflatin treatment on embryo development, DNA methylation, and gene expression. Oxamflatin treatment enhanced blastocyst formation of SCNT embryos in vitro. Embryo transfer produced more pigs born and fewer mummies from the Oxamflatin-treated group compared to the Scriptaid-treated positive control. Oxamflatin also decreased DNA methylation of POU5F1 regulatory elements and centromeric repeat elements in day-7 blastocysts. When compared to in vitro–fertilized (IVF) embryos, the methylation status of POU5F1, NANOG, and centromeric repeat was similar in the cloned embryos, indicating these genes were successfully reprogrammed. However, compared to the lack of methylation of XIST in day-7 IVF embryos, a higher methylation level in day-7 cloned embryos was observed, implying that X chromosomes were activated in day-7 IVF blastocysts, but were not fully activated in cloned embryos, i.e., reprogramming of XIST was delayed. A time-course analysis of XIST DNA methylation on day-13, -15, -17, and -19 in vivo embryos revealed that XIST methylation initiated at about day 13 and was not completed by day 19. The methylation of the XIST gene in day-19 control cloned embryos was delayed again when compared to in vivo embryos. However, methylation of XIST in Oxamflatin-treated embryos was comparable with in vivo embryos, which further demonstrated that Oxamflatin could accelerate the delayed reprogramming of XIST gene and thus might improve cloning efficiency.
Introduction
A
Oxamflatin has a strong potency as a hydroxamate-based HDACi. According to the in vitro testing and calculation of the half-maximal inhibitory concentration (IC50) by Huber et al. (2011), Oxamflatin is 100 times more potent than Scriptaid in inhibiting the activity of nuclear specific class I HDACs 1–3 and 8. Oxamflatin treatment (1 μM Oxamflatin for 9 h) significantly improved development of SCNT bovine embryos constructed from in vitro–matured oocytes (Su et al., 2011) and full-term development in the mouse (Ono et al., 2010). A limited study in pig SCNT embryos with either 9- (Park et al., 2012b) or 15-h (Hou et al. 2014) treatment of 100 or 1000 nM Oxamflatin showed that the treatment could be beneficial. However, the 9-h treatment is logistically inconvenient for our routine SCNT and embryo transfer, and the high dose may also be toxic to embryo and fetal development.
Abnormal epigenetic modifications such as DNA methylation and histone modifications often occur in cloned embryos (Dean et al., 2001; Santos et al., 2003). Thus, aberrant epigenetic reprogramming of somatic nuclei is thought to be a major reason for low cloning efficiency (Whitworth et al., 2011; Zhao et al., 2010b). The importance of correct reprogramming during cloning is highlighted by the finding that trichostatin A (TSA), another HDACi, improved the developmental rate of SCNT in mice (Kishigami et al., 2006). DNA methylation is a key player in epigenetic gene regulation. Erasure of DNA methylation was suggested to be one of the critical steps limiting the efficiency of cloning (Ficz et al., 2009). During normal early embryo development in pigs, locus-specific patterns of methylation and demethylation are observed (Zhao et al., 2013). DNA methylation defects of specific genomic regions and repetitive elements, and abnormal inactivation of the X chromosome in females, likely underlie the premature death of cloned animals because the offspring of live cloned animals appear normal (Ficz et al., 2009). Genes associated with pluripotency, POU5F1 and NANOG, centromeric repeat element, and XIST genomic regions were chosen for study of their level of DNA methylation in SCNT embryos. Promoter DNA demethylation in the POU5F1 and NANOG genes is necessary for reprogramming somatic cells into induced pluripotent stem cells (Maherali et al., 2007). The POU family transcription factor Pou5f1 (also known as Oct3/4) is required in mice for inner cell mass formation and pluripotency (Nichols et al., 1998). NANOG is required for the ground state of pluripotency in mouse embryos (Silva et al., 2009). It is specifically localized in nascent epiblasts, thus demarcating the epiblast from the hypoblast.
The centromere is a unique region on the chromosome that is required for attachment to the mitotic spindle and chromosome segregation and is essential to prevent aneuploidy. However, 44.5% of day-7 SCNT pig embryos can be aneuploid (You et al., 2010). In bovine cloned embryos, an aberrant hypermethylation in the centromeric repeat elements has been reported (Kang et al., 2001a). In porcine embryos, the methylation level of centromeric repeat elements was studied in in vitro–fertilized (IVF) blastocysts (Zhao et al., 2013). However, data were lacking in cloned embryos.
Female mammalian cells develop a dose compensation mechanism to prevent a difference in the expression of X-encoded proteins between males and females (Payer and Lee, 2008). Xist RNA, a long noncoding RNA, transcribed from the inactivated X chromosome, plays an important role in triggering X chromosome inactivation (XCI) (Payer and Lee, 2008). The XIST gene is expressed exclusively in female cells. High expression of Xist in the cloned mouse (Inoue et al., 2010) and porcine (Park et al., 2012a) blastocysts has been reported. This high expression of Xist was thought to be the main cause of adverse effects on the gene expression of cloned embryos, because normalization of Xist expression by genetic knockout remarkably improved the expression patterns of not only X-linked genes but also a number of autosomal genes in the cloned embryos. As a result, many of the knockout- (Inoue et al., 2010) and knockdown- (Matoba et al., 2011) cloned embryos survived to term, resulting in a significant increase in overall cloning efficiency. Aberrant expression of XIST was suggested to contribute to prenatal death of pig cloned embryos as well (Jiang et al., 2008). These findings indicate that mis-reprogramming of XIST after SCNT would compromise the development of cloned embryos.
Here we determined an effective dose of Oxamflatin treatment on SCNT embryo development in vitro. The effects of Oxamflatin at 150 nM on embryo development in vitro and full-term development in vivo were compared with another HDACi, Scriptaid, which increases SCNT embryo development and cloning efficiency compared to no treatment. Oxamflatin's role in regulating POU5F1, NANOG, centromeric repeat elements, and XIST DNA methylation and gene expression was also determined. The degree of XIST DNA methylation during early pregnancy at days 13, 15, 17, and 19 was examined. Finally, XIST methylation at day 19 of pregnancy was compared between SCNT cloned embryos and in vivo embryos, and between Oxamflatin-treated and nontreated control embryos.
Materials and Methods
All animal procedures were approved by the University of Missouri Animal Care and Use Committee (ACUC) and met the National Institutes of Health (NIH) guidelines (2011) for the care and use of animals. Unless described elsewhere, all chemicals and reagents were purchased from Sigma (St. Louis, MO, USA). Oxamflatin was purchased from Enzo Life Sciences Inc. (Farmingdale, NY, USA).
Donor cells
One genetically modified female cell line was used for the study of embryo development in vitro, DNA methylation, and gene expression. The primary cell line was established as previously described (Lai and Prather, 2003). Primary cultures were frozen in fetal bovine serum (FBS) containing 10% (vol/vol) dimethylsulfoxide (DMSO). The day before SCNT, fibroblasts were thawed and cultured in 50% Tissue Culture Medium-199 (TCM-199) and 50% Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% FBS, penicillin-streptomycin, and 5 ng/mL basic fibroblast growth factor (bFGF) overnight. A suspension of single cells was prepared by trypsinization of the cultured cells, followed by resuspension in manipulation medium [25 mM HEPES-buffered TCM-199 with 3 mg/mL bovine serum albumin (BSA)]. Six genetically modified cell lines, equally balanced between two treatment groups, were used for the in vivo full-term development study in conjunction with the service requirements of National Swine Resource and Research Center (NSRRC) at University of Missouri.
Somatic cell nuclear transfer
For SCNT, pig oocytes were purchased from ART Inc. (Madison, WI, USA) and matured according to the supplier's instructions. After 40 h of in vitro maturation, the oocytes were stripped of cumulus cells by gentle vortexing in 0.5 mg/mL hyaluronidase. Oocytes with good morphology and a visible polar body [metaphase II (MII)] were used for SCNT. Using micromanipulators on an inverted microscope, oocytes were enucleated by aspirating the polar body, MII plate, and a small amount of surrounding cytoplasm using a beveled glass pipette with an inner diameter of 17–20 μm. The medium for micromanipulation consisted of HEPES-buffered TCM-199, 0.3% BSA, and 7.5 μg/mL cytochalasin B (CB) for enucleation and no CB for injection. After enucleation, a single donor cell was injected into the perivitelline space adjacent to the oocyte membrane (Lai and Prather, 2003). Oocyte–donor cell couplets were placed into Porcine Zygote Medium 3 (PZM3) (Yoshioka et al., 2002) until fusion and activation. The fusion of oocyte–donor cell couplets and cloned embryo activation were accomplished with two direct current pulses of 1.2 kV/cm for 30 μsec with a 1-sec interval provided by a BTX Electro Cell Manipulator 2001 in a fusion medium (0.3 M mannitol, 1.0 mM CaCl2, 0.1 mM MgCl2, and 0.5 mM HEPES, pH 7.3). Oocytes were then incubated for 30 min in PZM3 and evaluated for fusion under a stereomicroscope. Only the fused embryos were used for experiments.
Postactivation treatment and embryo culture
Stock solutions of Oxamflatin and Scriptaid were dissolved in DMSO at 1 mM and stored at −80°C. Immediately after electrical activation, SCNT embryos were treated with various concentrations of Oxamflatin or 500 nM Scriptaid in PZM3 for 16 h according to the experimental design. After treatment, embryos were washed three times before culture in 25-μL drops (10–12 embryos per drop) of PZM3 supplemented with 1.69 mM of arginine at 38.5°C in humidified atmosphere of 5% O2, 90% N2, and 5% CO2 for 7 days (Bauer et al., 2010) for in vitro studies while embryos used for transfer were cultured for 17–20 h until surgical embryo transfer. For in vitro experiments, the percentage of cleaved embryos at 28 h and blastocyst formation at day 7 were evaluated with the day of SCNT designated as day 0. Pools of 10–15 blastocysts at day 7 were snap-frozen in liquid nitrogen and stored in −80°C freezer until RNA/DNA isolation.
Embryo transfer
Surrogate pigs on the first or second day of standing estrus (designated as gestational day 0) were used. The surrogates were prepared aseptically, and a caudal ventral incision was made to expose the reproductive tract. Embryos were transferred into one oviduct through the ovarian fimbria. Pigs were checked for pregnancy by abdominal ultrasound examination around day 30 and then checked once a week through gestation until parturition. The cloned piglets were delivered by cesarean section on day 116 or 117 of gestation and were hand raised.
Determination of cell numbers
Nuclei of blastocysts on day 7 of culture were stained with 4 μg/mL Hoechst 33342 for 15 min, and blastocysts were mounted on a slide in phosphate-buffered saline (PBS). The total number of nuclei was counted using a fluorescence microscope.
IVF and embryo culture
For embryo production by IVF, ovaries were collected from prepubertal gilts in a local slaughterhouse. Cumulus–oocyte complexes were aspirated and selected on the basis of uniform cytoplasm and multiple layers of cumulus cells. Oocytes were cultured in in vitro maturation (IVM) medium covered with mineral oil for 40–44 h at 38.5°C in a humidified atmosphere of 5% CO2 in air. After maturation, cumulus cells were removed as described as above. For IVF, 30 mature oocytes were transferred to a 50-μL droplet of equilibrated modified Tris-buffered medium (mTBM) (Katayama et al., 2007) covered with mineral oil at 38.5°C in 5% CO2 in air. For each replication, a frozen semen pellet was thawed in 3 mL of 0.1% (wt/vol) BSA in PBS, placed on 3 mL of 45% Percoll solution, and centrifuged for 25 min at 1900×g. The sperm pellet was transferred to 5 mL of mTBM and centrifuged for 10 min. The supernatant was removed. Sperm was reconstituted to 0.5×106 cells/mL, and 50 μL of resuspended sperm was added to each droplet containing MII oocytes. The sperm–oocyte-containing droplets were subsequently incubated at 38.5°C in 5% CO2 in air for 4–6 h. Then the oocytes were washed three times and cultured in PZM3 with 3 mg/mL BSA and 1.69 mM arginine at 38.5°C in a humidified atmosphere of 5% O2, 90% N2, and 5% CO2. Blastocysts were collected at day 7. Zonae were removed by brief acidic (pH 1.7) saline solution treatment. Embryos were washed three times in saline and frozen and stored at −80°C until assayed.
In vivo day-13, -15, -17, and -19 conceptus tissue collection
Gilts were artificially inseminated on the first day of estrus detection (designated day 0). On days 13, 15, 17, and 19 postinsemination, the gilts were euthanized and reproductive tracts were removed. The excised uterine horns were trimmed free of the broad ligament and ovary. The uteri were cut open with scissors, laid on a tray, and covered with saline to allow the conceptus to float up. Each individual conceptus was collected into a petri dish. Day-13, -15, and -17 embryos were elongated, and trophectoderm tissues were collected using a stereomicroscope. Day-19 fetuses had developed extraembryonic membranes and fetuses; extraembryonic membranes were collected, washed in saline solution, and stored at −80°C until analysis.
Sex determination of conceptus by PCR
The sex of embryos collected at days 13, 15, 17, and 19 was determined by using a PCR-based assay described previously (Hao et al., 2006). Briefly, genomic DNA was extracted from trophectoderm and extraembryonic membrane by using a DNeasy Kit (Qiagen, Germantown, MD, USA) and following the manufacturer's instruction. PCR was performed by using GoTaq Green Master Mix (Promega, Madison, WI, USA) with primers specific for sex-determining region Y (SRY, GenBank acc. no. NM_214452, Y chromosome-specific) and nuclear receptor DAX1 (GenBank acc. no. AF035816, X-chromosome-specific) loci. Oligonucleotides were purchased from Integrated DNA Technologies (IDT, Coralville, IA, USA). The reaction conditions were 94°C for 5 min followed by 30 cycles of 94°C (30 sec), 58°C (30 sec), 72°C (30 sec) with a final elongation step at 72°C for 3 min. Female embryos had a single DAX1 band at 179 bp, whereas male embryos had both the SRY (131 bp) and DAX1 bands. Genomic DNA from a known male and female pig was used as control, and water was used as the nontemplate control.
Genomic DNA and RNA isolation and quantitative RT-PCR
Genomic DNA and total RNA were isolated from day-7 blastocysts by using the Dynabeads mRNA DIRECT Purification Kit (Life Technologies, Grand Island, NY, USA) by following the manufacturer's instructions. After RNA was bound to the beads, the supernatant containing DNA was transferred into a new tube, precipitated by 3 M sodium acetate (pH 5.5) and 100% ethanol, and washed by 70% ethanol. The DNA pellet was dried completely to remove any trace of ethanol and resuspended with H2O for bisulfite sequencing assay.
First-strand complementary DNA (cDNA) was synthesized by using a SuperScript VILO cDNA Synthesis Kit (Life Technologies). Real-time quantitative PCR (qRT-PCR) was performed by using iQ SYBR Green Supermix in an iCycler iQ Single-Color Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). Melting curves were generated following RT-PCR to assess the specificity of amplicons. Each sample had only one peak. Thus, no samples were excluded for final analysis. Primer sequences for four genes were: POU5F1, forward (5′-TTT GGG AAG GTG TTC AGC CAA ACG-3′) and reverse (5′-TCG GTT CTC GAT ACT TGT CCG CTT-3′); NANOG, forward (5′-AGG ACA GCC CTG ATT CTT CCA CAA-3′) and reverse (5′-AAA GTT CTT GCA TCT GCT GGA GGC-3′) (Zhao et al., 2009); XIST, forward (5′-TCT TTG AAG GTC CTC TTC TGT CCA-3′), and reverse (5′-GAG TAG AAG AAG TCC ACA GCA ATT C-3′); YWHAG, forward (5′-TCC ATC ACT GAG GAA AAC TGC TAA-3′), and reverse (5′-TTT TTC CAA CTC CGT GTT TCT CTA-3′) (Whitworth et al., 2005).
Gene expression data determined by qRT-PCR were obtained from four independent biological and two technical replicates and were calculated relative to the housekeeping gene, tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, gamma polypeptide (YWHAG) (Whitworth et al., 2005). Expression levels were determined using the comparative threshold cycle (CT) method for each gene. The CT for the target gene was subtracted from the CT for YWHAG to obtain the change (Δ) in CT (ΔCT). The relative amount of each mRNA was calculated by assuming an amplification efficiency of two and using the equation 2−ΔCT. The 2−ΔCT values were first analyzed for normality and skewness before being log transformed. The log-transformed values were then analyzed using the general linear model (PROC GLM) in the Statistical Analysis System (SAS v. 9.3; SAS Institute, Cary, NC, USA).
Bisulfite sequencing analysis
Within each replicate, 10–15 blastocysts from the same treatment group were pooled to generate enough DNA for bisulfite conversion and subsequent analysis. Genomic DNA was treated with sodium bisulfite and immediately purified by using an EpiTect Fast Bisulfite Conversion Kit (Qiagen). The bisulfite primers were designed by using online MethPrimer software (www.urogene.org/methprimer/index1.html). The primer sets for POU5F1, NANOG, and the centromeric repeat element had been tested, validated, and published previously by Zhao et al. (2013). XIST primers were designed on the basis of the complete XIST sequence (GenBank acc. no. KC753464.1) (Hwang et al., 2013). The XIST PCR amplicon was located in exon 1 and within the CpG island that could be defined by the MethPrimer software. The forward primer sequence for XIST bisulfite PCR was 5′-ATG TGG ATA GTA TGG TAG TTT GTT A-3′, and reverse primer sequence as 5′-TCC ACC ACC CTT TCT AAT TAA ATA TAT-3′. PCR was performed by using a GoTaq Green Master Mix (Promega) with bisulfite-converted genomic DNA as the template. The PCR program for all genomic regions studied consisted of an initial denaturation step at 95°C for 4 min, followed by 40 cycles of denaturation at 95°C for 45 sec, annealing at 56°C for 1 min, and extension at 72°C for 45 sec. A final extension of 72°C for 15 min was also included.
The PCR product was loaded and resolved in a 1.5% agarose gel, and then extracted and purified by using a Wizard SV Gel and PCR Clean-Up System (Promega). Purified PCR fragments were then cloned into a pCR4-TOPO vector, which was included in a TOPO TA Cloning Kit for sequencing (Invitrogen, Grand Island, NY, USA). The TOPO cloning reaction was subsequently transformed into One Shot TOP10 chemically competent cells (Invitrogen) and grown on Luria–Bertani–kanamycin (50 mg/mL) agar plates overnight. For each transformation, 10–15 clones were randomly selected, and plasmid DNA was isolated by using a PureLink Quick Plasmid Miniprep Kit (Invitrogen). Plasmids were further screened by PCRs, and only positive clones were submitted to the DNA core facility at University of Missouri for sequencing.
Data interpretation
Sequencing data were aligned to the reference sequences by using MacVector (v. 11.6, MacVector Inc., Cary, NC, USA). Reference sequences were created by replacing a C with a T in non-CpG sites but leaving the C in CpG sites unchanged. The bisulfite treatment converts an unmethylated C into a U, which will be amplified as a T after PCR but will not change the methylated C. Therefore, for each CpG site, if it is a TG, instead of CG, this means that this CpG is unmethylated and is represented as an open circle in the diagrams. On the other hand, if it is a same CG, it means that this CpG is methylated and prevented from bisulfite conversion, and this methylated C is represented with a filled circle. The representative clones were carefully selected so that each one was at least one nucleotide (either in CpG or non-CpG site) different from another within the amplicon.
Combined bisulfite restriction analysis
The semi-quantitative combined bisulfite restriction analysis (COBRA) of the XIST methylation level within the PCR amplicon (191 bp) was performed as described by Xiong and Laird (1997). In brief, the bisulfite PCR was carried out as described above by using the same reaction conditions. PCR products were purified by using a QIAquick PCR Purification Kit (Qiagen). Purified DNA was digested with BstUI (Thermo Scientific, Waltham, MA, USA) restriction enzyme with a recognition sequence of CGCG and cut between the two CGs at 121 of the 191-bp XIST PCR amplicon at 37°C for 2 h. Polyacrylamide gel electrophoresis (PAGE) was performed to resolve the restricted products. Gel images were taken with FOTO/Analyst Investigator/FX Systems (Fotodyne, Hartland, WI, USA). The band intensity was determined by NIS-Elements software (v. 4, Nikon Inc. Melville, NY) to determine if both alleles were being amplified at equal rates during bisulfite PCR.
Statistical analysis
Data for the dependent variables, percent cleavage at 28 h of culture, percent blastocyst formation at day 7, number of nuclei within the blastocyst-stage embryos, total embryos for each transfer, cloning efficiency, and litter size were analyzed using the PROC GLM procedure of SAS software (v. 9.3) (Statistical Analysis Systems, 2010), with treatment as the main effects. The number of nuclei in blastocyst-stage embryos was square root transformed to approach a normal distribution. The liveborn and number of mummies between Oxamflatin and Scriptaid groups were compared by using Pearson's chi-squared test. DNA methylation data generated from bisulfite sequencing for POU5F1, NANOG, centromeric repeat elements, and XIST gene were calculated as methylated CpGs/total CpGs*100 for each allele. Subsampling nested analysis of variance (ANOVA) (allele nested within culture well) was performed by using PROC GLIMMIX of SAS. The experimental unit was the culture well in which embryos were treated and cultured. In the results, the least-square means and the standard errors of means are presented. A value of p<0.05 was considered to be statistically significant.
Results
Determination of optimal Oxamflatin concentration in vitro
The objective of this experiment was to determine a low, nontoxic, but effective, dose of Oxamflatin to improve SCNT embryo development in vitro. Therefore, on the basis of its IC50 of 15 nM (Huber et al., 2011), the effects of Oxamflatin at 15, 30, and 60 nM treatment for 16 h were first determined on the percentage of cleaved embryos at 28 h after activation, blastocyst formation, and total cell number at day 7. None of these doses had any effect on embryo development (such as percent blastocyst formation at day 7: 32.3±3.3, 23.0±2.2, 28.6±2.9, 28.2±4.5% for the zero control, 15, 30, and 60 nM groups, respectively), indicating that low concentrations of Oxamflatin did not improve embryo development.
The calculation of IC50 by Huber et al. (2011) indicated that a 200 nM dose of Oxamflatin would inhibit over 95% of the deacetylase activity of nuclear-specific class I HDACs. A new set of Oxamflatin doses at 0, 150, 300, and 500 nM were tested on embryo development in vitro. There were no effects on the percentage of cleaved embryos by Oxamflatin treatment (Fig. 1A). However, Oxamflatin treatment at 150 and 300 nM enhanced blastocyst formation (Fig. 1B), whereas 500 nM was not different from the control, suggesting that 500 nM may start to have negative effects on SCNT embryos. There was no difference in blastocyst nuclei numbers among the dose groups, and they averaged 34.6±2.4.
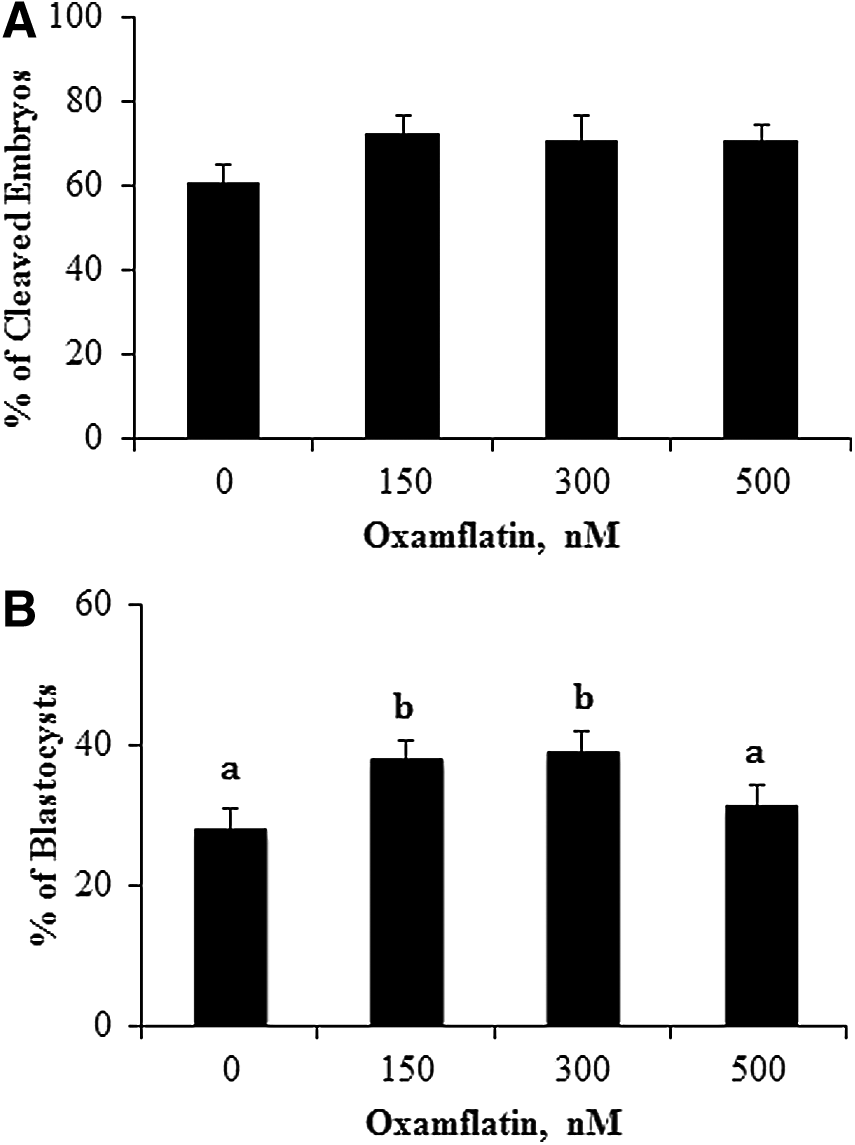
Percentage of embryos cleaved at 28 h (
Scriptaid treatment (500 nM) has been shown to improve in vitro and in vivo developmental competence of cloned embryos (Zhao et al., 2009; Zhao et al., 2010a). The treatment effects on embryo development were therefore compared between Oxamflatin at 150 nM and Scriptaid at 500 nM. There was no difference in percentage of cleaved embryos at 28 h postactivation, blastocyst formation, and total cell number at day 7 between the two groups (Fig. 2).

Percentage of cleavage at 28 h postactivation, blastocysts formed (%) at day 7, and total cell number of blastocysts in the Oxamflatin and Scriptaid groups. Embryos were treated with 150 nM Oxamflatin or 500 nM Scriptaid for 16 h, then were washed and cultured in PZM3 until day 7.
Comparison of Oxamflatin with Scriptaid for embryo full-term development
As previously demonstrated, Scriptaid treatment of SCNT embryos enhanced in vivo full-term development, compared to the nontreated control (Zhao et al., 2009; Zhao et al., 2010a). As described above, there was no difference in cleavage, blastocyst formation, and total cell number between the 150 nM Oxamflatin and 500 nM Scriptaid groups. The objective of this experiment was to compare Oxamflatin to Scriptaid (a positive control group) on the effects on full-term development. For simple comparison, the nontreated negative control was not included. Ten out of 14 surrogates (71%) were pregnant at day 114 in the Oxamflatin-treated group and 12 out of 14 (85%) were pregnant in the Scriptaid group (Table 1). Twenty-two live piglets and one mummy were born from the 10 pregnant surrogates in the Oxamflatin group, and 15 live pigs and six mummies were born from the 12 pregnant surrogates in the Scriptaid group (chi-squared test, p<0.03). All of the live cloned piglets looked normal. If based on only pregnant surrogates, the resulting cloning efficiency of 0.95±0.31% in the Oxamflatin-treated group was numerically higher than that of 0.59±0.29% in the Scriptaid group, but it was not statistically different. If all surrogates were included, the cloning efficiency was 0.68±0.25% and 0.51±0.25% for the Oxamflatin and Scriptaid groups, respectively, with no significant difference between them.
Cloning efficiency 1 was the mean that was calculated based on only pregnant recipients.
Cloning efficiency 2 was the mean that calculated based on all recipients, with nonpregnant and return recipients as zero.
Numbers differ with different superscripts by chi-squared test (p<0.03).
Effects of Oxamflatin treatment on the level of methylation in NANOG, POU5F1, XIST, and centromeric repeat element in day-7 SCNT blastocysts and comparison to IVF counterparts
Reprogramming by SCNT involves extensive DNA demethylation (Jaenisch et al., 2004). The DNA methylation status of POU5F1, NANOG, XIST and centromeric repeat elements was examined in donor cells before SCNT, as a reference point, and in day-7 blastocysts. The methylation level of POU5F1 was 37.4±2.7% in the donor cells and 6.7±1.6% in the day-7 SCNT control embryos. Oxamflatin treatment decreased POU5F1 methylation to 3.5±1.8% in SCNT blastocysts (p<0.05) (Fig. 3, Table 2). The methylation level in the NANOG promoter area was 24.2±1.8% in the donor cells before nuclear transfer. After SCNT and culture to day 7, it dropped to 2.1±1.0% and 0.6±0.9% for the SCNT control and Oxamflatin-treated embryos, respectively, with no difference between them (Fig. 4, Table 2). In donor cells, 45.3±5.8% of CpGs in centromeric repeat elements were methylated. In the SCNT embryos, Oxamflatin reduced methylation from 27.3±3.1% measured in the control embryos to 18.2±3.2% (p<0.05) (Fig. 5, Table 2). The average methylation of XIST in donor cells was 42.4±6.4%. This CpG island had two sites that were not methylated in any of the samples. However, the remaining nine CpGs were methylated in eight of 15 clones, e.g., suggesting a parental imprint of about 50%. This provides indirect evidence that the CpG islands studied represented the real-time status of the XIST locus in the cell and provided a good marker for reprogramming studies. The XIST methylation level in day-7 blastocysts was lower than the donor cells, but not different between Oxamflatin (11.8±3.2%) and control SCNT (11.7±3.2%) groups (Fig. 6A, B, Table 2).
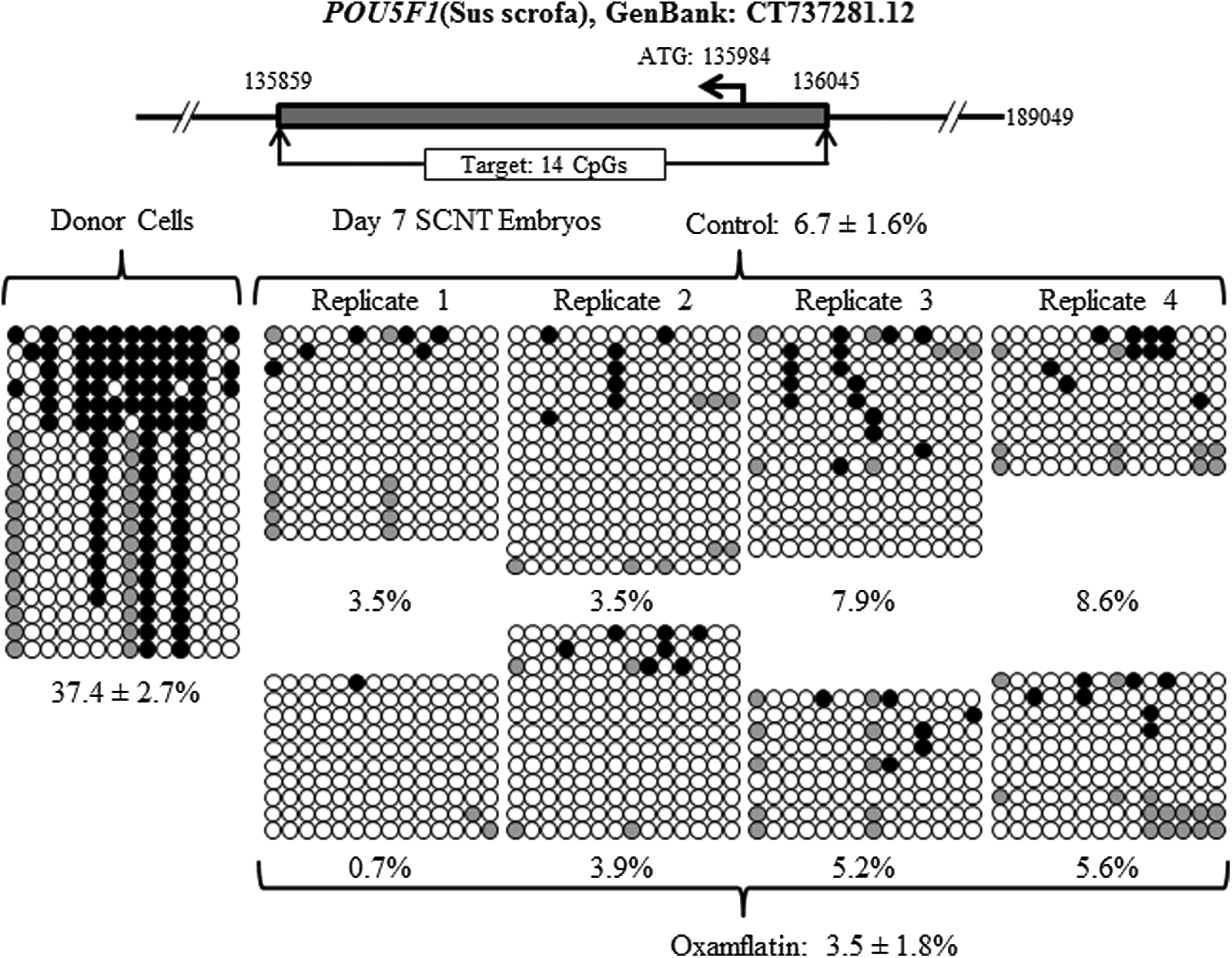
Methylation status of POU5F1 locus in the donor cells and day-7 SCNT embryos. A dramatic decrease in methylation from donor cells to day-7 embryos; Oxamflatin treatment further decreased its methylation (p<0.05). (Solid-filled circles) Methylated cytosine; (open circles) unmethylated cytosine in each CpG site; (gray circles) mutated and/or single nucleotide polymorphism (SNP) variation at certain CpG sites. Each row of circles represents an individual clone that contains the inserted amplicon. The top diagram in each figure following schematically denotes the genomic location of the target DNA methylation region. ATG, start codon. Note, the same legend applies to Figures 4–7. The top diagrams that indicate the location of interest genes studied were the same promoter region of POU5F1 and NANOG and centromeric repeat elements gene as reported by Zhao et al. (2013).
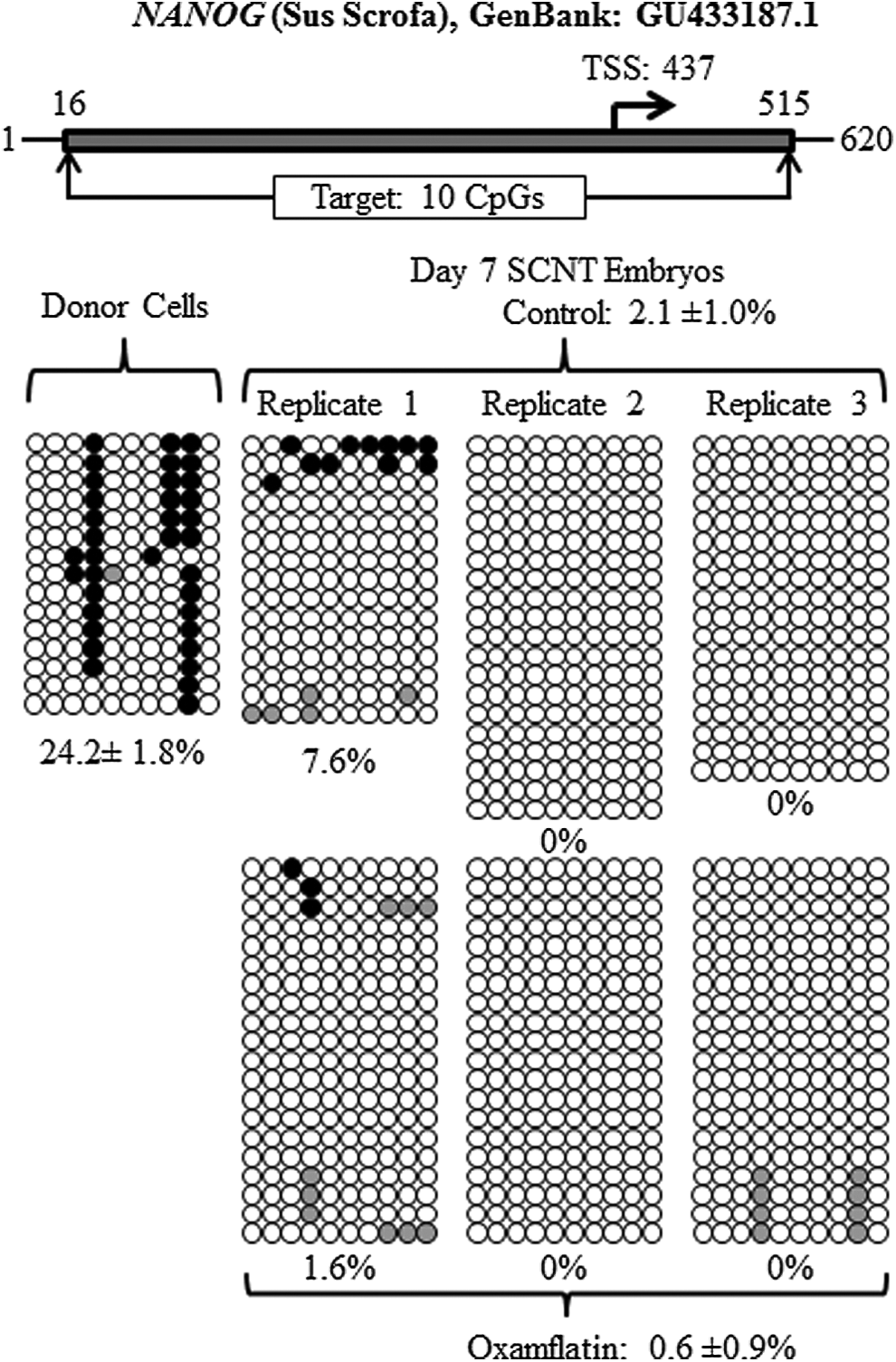
Methylation status of NANOG locus in the donor cells and day-7 SCNT embryos. A dramatic decrease in methylation from donor cells to day-7 embryos was observed. Oxamflatin treatment did not have any effect on NANOG methylation.

Methylation status of centromeric repeat elements in the donor cells and day-7 SCNT embryos. There was a decrease in methylation from donor cells to day-7 embryos. Oxamflatin treatment further decreased methylation from 27.3±3.1% of controls to 18.2±3.2% of the Oxamflatin-treated group (p<0.05).
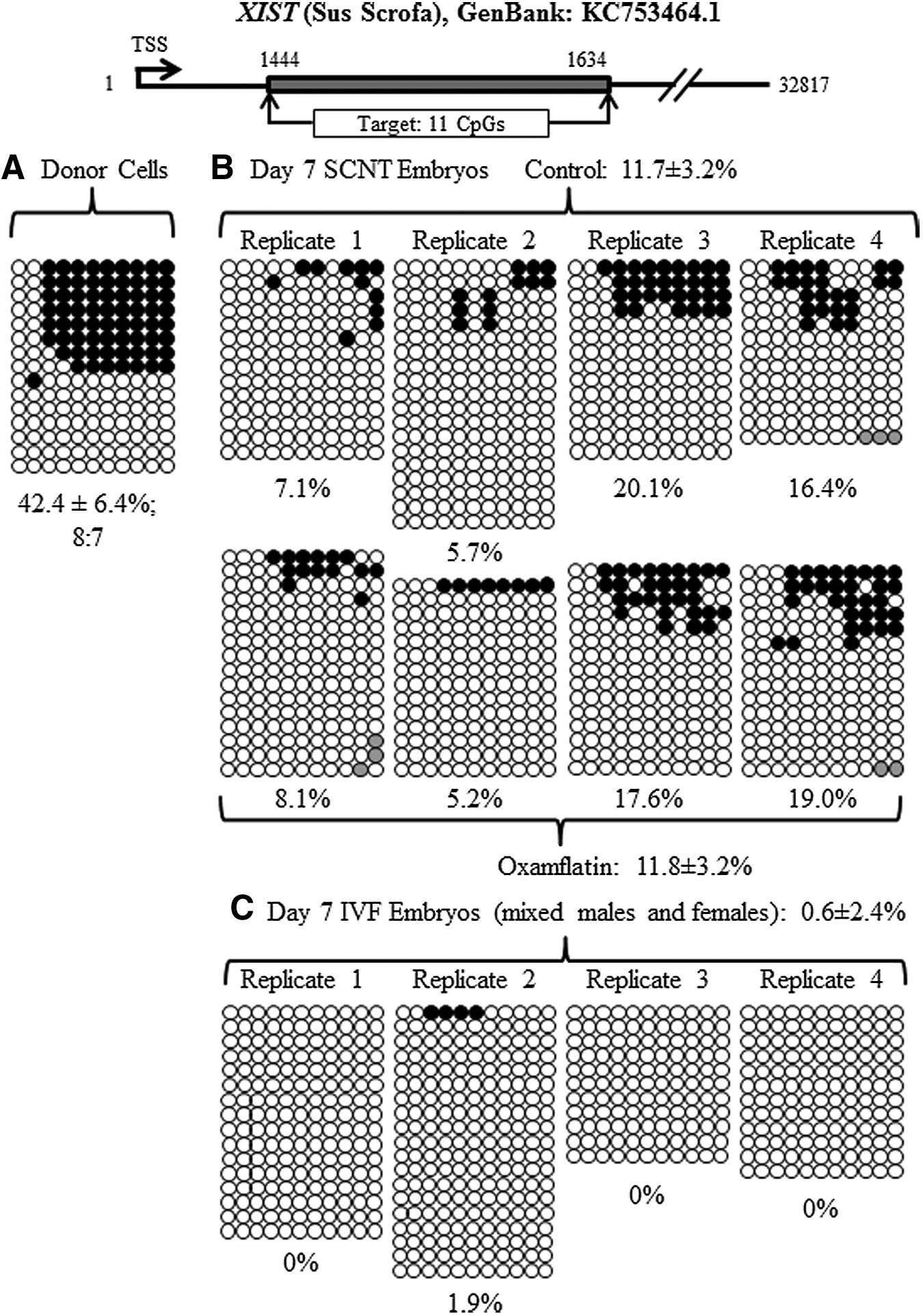
Methylation status of the XIST locus in the donor cells and day-7 SCNT and IVF embryos. There was a decrease in the XIST DNA methylation level from donor cells to day-7 embryos. Oxamflatin treatment did not have an effect on XIST methylation. A high level of methylation in the XIST gene indicated that reprogramming of XIST was not completed in SCNT embryos, as compared to no methylation of IVF embryos.
Methylation status in the day-7 in vitro–fertilized (IVF) embryos for POU5F1, NANOG, and centromeric repeat elements was taken from the report by Zhao et al. (2013) of our laboratory using the same bisulfite PCR primers to study the same gene region, thus direct comparison with cloned embryos could not be done.
Means with different superscripts within each column differ (p<0.05).
Using the same bisulfite sequencing approach, Zhao et al. (2013) reported methylation levels of 7.1% in POU5F1, 2.5% in NANOG, and 29.1% in centromeric repeat elements in day-7 pig IVF blastocysts. The methylation levels in these genomic regions of SCNT embryos were similar to IVF embryos, indicating that reprogramming of these loci in SCNT embryos was successful. The XIST gene methylation was not studied by Zhao et al. (2013). Therefore, we collected day-7 IVF blastocysts from four biological replicates and found the average methylation level of XIST was 0.6±2.4% (Fig. 6C); almost all alleles were unmethylated, indicating that the XIST gene or/and both X chromosomes were activated in day-7 IVF embryos. However, compared to IVF embryos, the overall XIST methylation in SCNT embryos from both control and Oxamflatin groups was higher (averaged at 11.7±1.7%, p<0.01), indicating incomplete reprogramming of XIST.
Expression of POU5F1, NANOG, and XIST and correlation with methylation
There was no difference in POU5F1 (14.1±0.8 vs. 16.3±0.8 relative arbitrary units) and NANOG (0.26±0.04 vs. 0.25±0.04) gene expression relative to a housekeeping gene (YWAHG) between the Oxamflatin and control groups. However, Oxamflatin treatment tended to decrease XIST expression in day-7 blastocysts compared to SCNT controls (18.8±0.8 vs. 21.7±0.8, p<0.1). Interestingly, as shown in Figure 7, XIST gene expression was positively correlated with its DNA methylation level (r=0.72, p<0.05).

Association between XIST gene expression and its methylation level in day-7 SCNT embryos. Each dot represents one pooled blastocyst sample. (Triangle) Oxamflatin-treated group; (diamond) control group.
XIST DNA methylation in day-13, -15, -17, and -19 in vivo embryos
Day-13, -15, -17, and -19 embryos/conceptuses were collected and sex genotyped. Only female samples were analyzed for XIST methylation. The percentage of methylated CpGs out of total sites analyzed was 24.0±7.5, 31.8±6.8, 23.4±6.7, and 31.3±5.4% for day-13,-15, -17, and -19 embryos, respectively, with no difference among them. In the somatic female tissue, a 50% methylated and 50% unmethylated allele ratio was expected; however, about half of the assayed tissue samples showed less methylated than unmethylated alleles (Fig. 8A). To confirm if this is caused by biological variation and not by a PCR artifact, the bisulfite-mutagenized DNA of one day-13 and three day-19 embryos were subjected to COBRA. Donor cells (i.e., assay control) showed 50% methylated and 50% unmethylated alleles (Fig. 9), which supports the hypothesis that one X chromosome is inactivated in somatic female cells. DNA methylation levels of XIST in selected samples obtained by COBRA also agreed with bisulfite assay. The level of methylation at the first exon of XIST was higher in day-13 female embryos when compared to day-7 IVF embryos (24±7.5% vs. 0.6±2.4% methylated CpGs), indicating that XCI was underway. XCI in the pig is not yet completed at day-19 gestation, as evidenced by the lower than expected (i.e., 50% somatic cells; Fig. 9) level of methylation of this gene (31±5.4%; Fig. 7A).
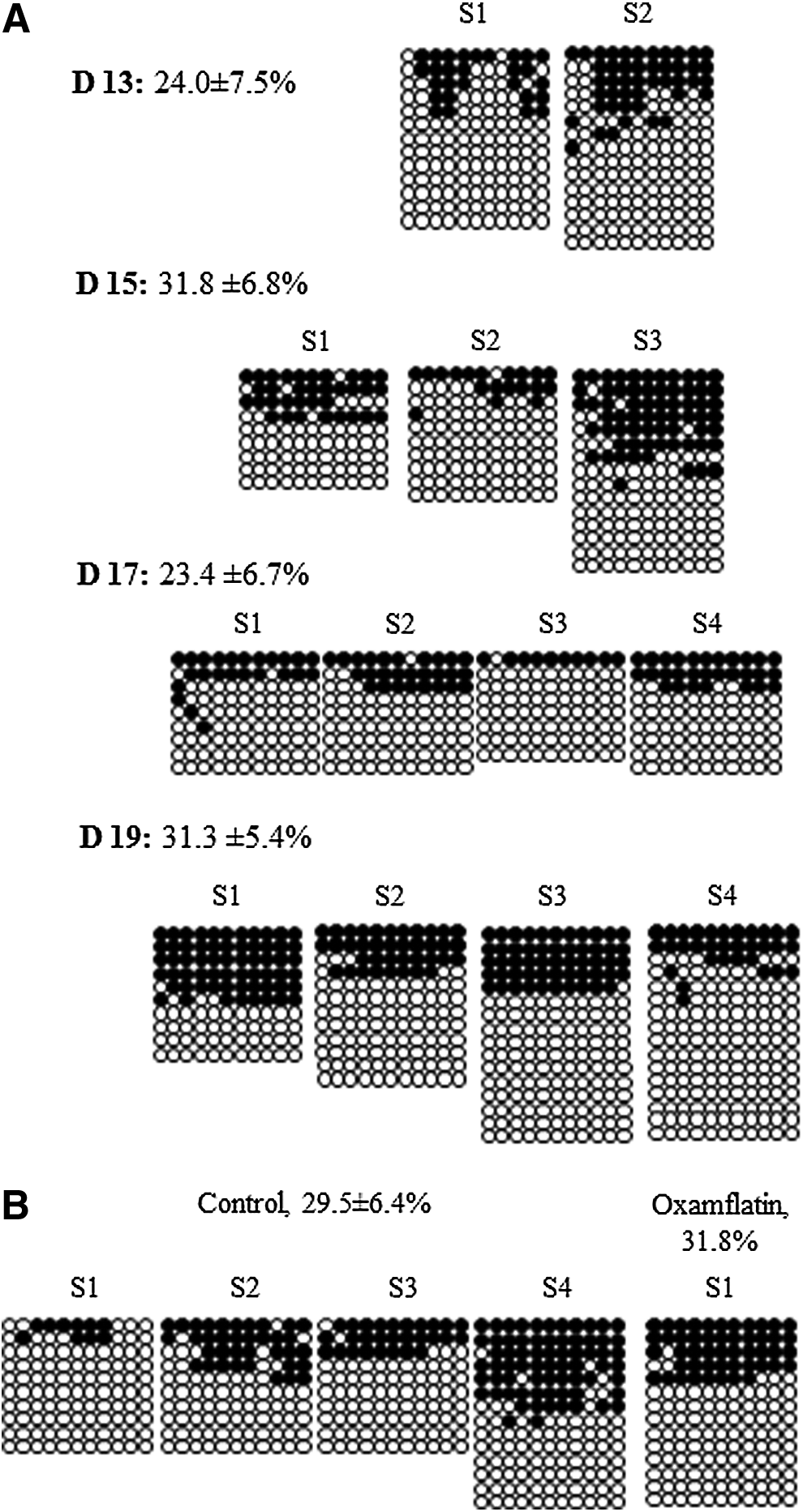
XIST DNA methylation in day-13, -15, -17, and -19 in vivo (
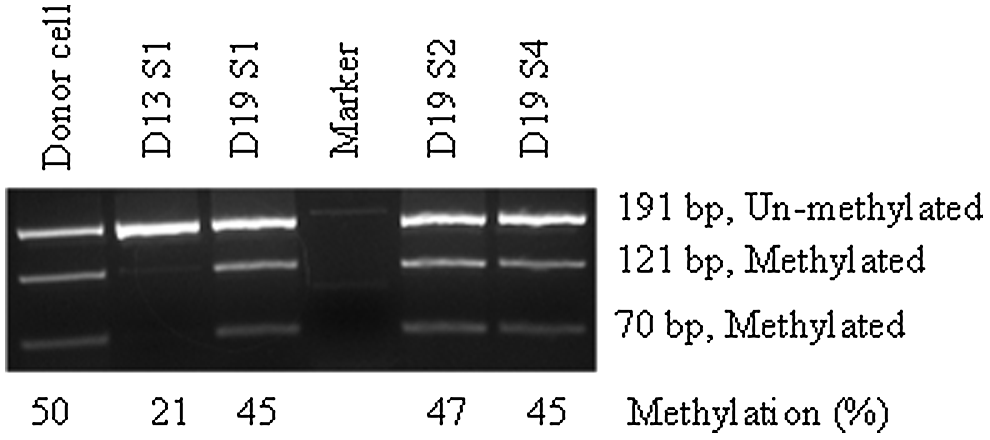
Validation of XIST methylation by COBRA. A group of selected samples were used to confirm bisulfite sequencing data. Even band intensity (50/50) for both the cut methylated allele and the uncut unmethylated allele in the donor cells and day-19 embryonic membrane was measured, whereas day 13 had almost no cut band. COBRA was performed using the restriction enzyme BstUI, which cuts the methylated allele at 121 bp between CpG 7 and 8 (amplicon size 191 bp).
Comparison of XIST methylation between day-19 SCNT extraembryonic membranes and in vivo counterparts
We then wanted to determine the methylation level of XIST in SCNT-cloned day-19 embryos and if Oxamflatin treatment had any effects on the XCI. To this end, SCNT embryos treated with DMSO as vehicle control or Oxamflatin were transferred into surrogates and embryos were collected at day 19. Eleven elongated embryos were recovered from the control group. They were all shaped as filaments, ranging from 10 to 20 mm in length; thus, they were developmentally behind compared to the well-developed in vivo embryos that were fully differentiated fetuses and had clear extraembryonic membranes. One such well-developed fetus was recovered from the Oxamflatin group, which looked similar to the day-19 in vivo counterparts. Four out of 11 embryos from the control and one from Oxamflatin group were assayed for XIST DNA methylation. The overall percentage of methylated CpGs was not different between Oxamflatin-treated embryo and nontreated controls (31.8 vs. 29.5%; Fig. 7B).
Discussion
Few studies have investigated the effects of Oxamflatin on SCNT embryo development, or on gene methylation and expression. No data are available about X chromosome activation and inactivation during early development either in normal embryos or in cloned pig embryos. Our results showed that Oxamflatin treatment (150 and 300 nM) for 16 h increased SCNT embryo blastocyst formation in vitro. Because Scriptaid treatment improves embryo development in vitro and full-term development in vivo, a comparison was warranted. Oxamflatin (150 nM) was as effective as Scriptaid (500 nM) in improving embryo development in vitro and better full-term development in vivo, as indicated by more live cloned pigs born (22 vs. 15). To address the role of Oxamflatin in regulating methylation and gene expression, POU5F1, NANOG, centromeric repeat elements, and XIST were studied. Oxamflatin decreased DNA methylation of POU5F1 and centromeric repeat elements. It did not affect DNA methylation of XIST, although Oxamflatin tended to decrease XIST gene expression in day-7 blastocysts. The methylation level of POU5F1, NANOG, and the centromeric repeat was not different between cloned and IVF embryos, but XIST methylation was significantly higher in cloned embryos than IVF embryos. The lack of DNA methylation of XIST in day-7 IVF embryos suggests that both X chromosomes were activated, but it was not known when the XCI occurred during pig embryogenesis. Further study detected increased DNA methylation of XIST between day 7 and days 13–19, suggesting that XCI was underway.
Compared to controls, Oxamflatin treatment (150 and 300 nM) of cloned embryos for 16 h improved the blastocyst formation in vitro and was not different from Scriptaid (150 nM Oxamflatin and 500 nM Scriptaid). We conclude that low concentrations of Oxamflatin are as effective as a high dose of Scriptaid. There were more (p<0.03) live cloned pigs (22 vs. 15) and fewer mummies (one vs. six) from the Oxamflatin-treated group compared to Scriptaid. Thus, Oxamflatin may be more potent and a less toxic HDACi than Scriptaid, which was supported by the lower IC50 of Oxamflatin than Scriptaid in the ability of inhibiting HDAC activity in vitro (Huber et al., 2011).
DNA methylation, a key epigenetic factor, modifies and regulates the chromatin structure and also plays a crucial role in SCNT reprogramming. To understand the possible underlying mechanism of how Oxamflatin, or HDACi, in general, improves cloning efficiency, DNA methylation of POU5F1, NANOG, centromeric repeat elements, and XIST and gene expression of POU5F1, NANOG, and XIST were studied in the Oxamflatin-treated and nontreated blastocysts. Oxamflatin decreased DNA methylation of POU5F1 and centromeric repeat elements. This observation supports the theory that HDACi can induce hyperacetylation of the core histones, resulting in structural changes in chromatin that permit transcription, as well as enhanced DNA demethylation of the somatic cell genome after SCNT (Kishigami et al., 2006), which is a necessary part of epigenetic reprogramming (Armstrong et al., 2006; Simonsson and Gurdon, 2004). Oxamflatin decreased DNA methylation and thus may activate genes that are key for development (Cervoni and Szyf, 2001).
In contrast, Oxamflatin had no effect on XIST DNA methylation. Both the control and Oxamflatin groups had 11.7% methylated CpGs. XIST may be a difficult gene to reprogram because Do et al. (2008, 2009) fused somatic cells with embryonic stem cells in mice and reported that pluripotency genes could be reprogrammed to a pluripotent state within 2 days, but it would take at least 9 days to reprogram/demethylate Xist. In the current study, Oxamflatin had no effect on XIST methylation but it tended to decrease Xist gene expression, which may be due to activation of Dnmt3a and Tsix, two negative regulators of Xist as demonstrated by Do et al. (2008) in the mouse. The inhibitory effect on XIST expression had also been reported in human embryo stem cells when they were treated with sodium butyrate, another HDACi (Ware et al., 2009). This inhibitory effect on XIST gene expression may also be a beneficial factor to enhance porcine cloned embryo development, but to less degree, as compared to the mouse knockdown and knockout of Xist, which resulted in dramatic improvements in cloning efficiency (Inoue et al., 2010; Matoba et al., 2011).
When compared with the IVF blastocyst-stage embryos as reported by Zhao et al. (2013), we found that the methylation level of POU5F1, NANOG, and centromeric repeat elements in day-7 cloned embryos were very similar, indicating that reprogramming of these genes were successful and completed at day 7. The similarity of DNA methylation in centromeric repeat elements between cloned pig embryos and IVF embryos agreed with a report by Kang et al. (2001b). An HDACi of the same category, TSA, not only modifies histone acetylation but also potentially induces DNA demethylation in human endometrial cells (Xiong et al., 2005). In bovine SCNT embryos, there is aberrant hypermethylation in the satellite I region (Kang et al., 2001a), and HDACi decreased DNA demethylation in the satellite I region (Wang et al., 2011; Wee et al., 2007). We found that Oxamflatin-treated SCNT embryos had significantly lower DNA methylation in the satellite I region than nontreated SCNT embryos. Like TSA, Oxamflatin treatment seems to have a ‘correcting’ effect on the DNA methylation status of the satellite I region.
In the pig, XIST gene methylation data were not available. Surprisingly, the first exon of the XIST gene was almost completely unmethylated at day 7 of development (Fig. 6C), suggesting that both X chromosomes were reactivated at this time. These observations were supported by the theory that two active X chromosomes are present in pluripotent stem cells and their embryonic ancestor cells such as epiblast, which all express characteristic pluripotency markers like Pou5f1 and Nanog (Payer and Lee, 2008). The Pou5f1 protein was expressed by all porcine embryo cells at day 7 (Kirchhof et al., 2000), thus supporting our current observation of unmethylated status in the XIST gene. Compared to IVF embryos, the higher DNA methylation in the XIST gene of cloned embryos indicated that reprogramming of the XIST gene was not completed at day 7.
For X inactivation maintenance, DNA methylation is a key stabilizing factor, as deletion of the maintenance DNA methyltransferase gene Dnmt1 results in X reactivation in the embryo proper (Sado et al., 2000). Thus, the degree of DNA methylation in the XIST gene may represent a powerful tool to study XCI. DNA methylation has long been correlated with imprinted XIST expression and thus involved in regulating XCI (Monk, 1995; Norris et al., 1994). Our observation on XIST methylation suggests that XCI in pigs is initiated by implantation and day 13 and not yet completed by day 19. As demonstrated by Spencer et al. (2006), POU5F1 gene expression is highest at days 10 and 12, and decreases significantly at days 15 and 17. Again the tight association between XCI and loss of pluripotency agrees with the theory that XCI is correlated with the differentiated cell state (Payer and Lee, 2008). Thus, the POU5F1 gene expression pattern as reported by Spencer et al. (2006) further supports our observation on the timing of XCI. In the mouse, preimplantation embryos undergo imprinted X-inactivation, and this inactive state is erased in the embryo proper before random X inactivation occurs. X reactivation occurs in the inner cell mass of blastocysts between E3.5 and E4.5 around the time point of implantation into the uterus (Mak et al., 2004; Okamoto et al., 2004). This reprogramming event is restricted to epiblast cells in the mouse. Our results show that similarities exist between the mouse and pig in the timing of X chromosome reactivation at the blastocyst stage and the initiation of inactivation during peri-implantation stages.
In summary, we optimized the Oxamflatin concentrations and demonstrated that Oxamflatin was a potent HDACi for improving cloned embryo development in vitro and in vivo. More live pigs were born to the Oxamflatin group than the Scriptaid group, which suggested to us that Oxamflatin may also be a more effective inhibitor. Oxamflatin treatment enhanced nuclear reprogramming by reducing DNA methylation, thus decreasing XIST gene expression. Also we demonstrated in the pigs that XCI is initiated around day 13 and is not yet finished by day 19 of gestation. However, more experiments are needed to compare with other HDACi, such as class I and IIa/b inhibitor, suberoylanilide hydroxamic acid (SAHA), and its hydrophobic derivative, 4-iodo-SAHA (I-SAHA), directly on DNA methylation and XIST reprogramming, so that the best inhibitor to enhance pig cloning efficiency can be selected. Besides, their effects on development may also be cell line specific, which requires further studies as well.
Footnotes
Acknowledgments
The authors would like to thank all the members in both Dr. Prather's laboratory and the National Swine Resource and Research Center. We thank Lonnie Dowell, Jason Dowell, Stephanie Murphy, and Josh Benne for managing surrogate gilts and surgical assistance. The authors would like to thank Dr. Mark Ellersieck for helping with the statistical analysis of the data. We appreciate funding from the National Institutes of Health (RR018877, U42 OD011140), and Food for the 21st Century Program.
Author Disclosure Statement
The authors declare that they have no conflicting financial interests.