
Abstract
Abstract
The present study was undertaken to examine the effect of Scriptaid treatment on histone acetylation, DNA methylation, expression of genes related to histone acetylation, and development of buffalo somatic cell nuclear transfer (SCNT) embryos. Treatment of buffalo SCNT embryos with 500 nM Scriptaid for 24 h resulted in a significant increase in the blastocyst formation rate (28.2% vs. 13.6%, p<0.05). Meanwhile, treatment of buffalo SCNT embryos with Scriptaid also resulted in higher acetylation levels of H3K18 and lower methylation levels of global DNA at the blastocyst stage, which was similar to fertilized counterparts. The expression levels of CBP, p300, HAT1, Dnmt1, and Dnmt3a in SCNT embryos treated with Scriptaid were significantly lower than the control group at the eight-cell stage (p<0.05), but the expression of HAT1 and Dnmt1a was higher than the control group at the blastocyst stage (p<0.05). When 96 blastocysts developed from Scriptaid-treated SCNT embryos were transferred into 48 recipients, 11 recipients (22.9%) became pregnant, whereas only one recipient (11.1%) became pregnant following transfer of 18 blastocysts developed from untreated SCNT embryos into nine recipients. These results indicate that treatment of buffalo SCNT embryos with Scriptaid can improve their developmental competence, and this action is mediated by resulting in a similar histone acetylation level and global DNA methylation level compared to in vitro–fertilized embryos through regulating the expression pattern of genes related to histone acetylation and DNA methylation.
Introduction
S
Many efforts have been made to improve the efficiency of SCNT, including ameliorating enucleation procedures (Kuhholzer et al., 2000), using different types of donor cells (Enright et al., 2003a), and prolonging intervals from fusion to activation (Wakayama et al., 2003; Wang et al., 2011). In the past decades, analysis of cloned embryos or offspring revealed that aberrant epigenetic modification, such as histone acetylation, DNA methylation, histone phosphorylation, adenosine diphosphate (ADP)-ribosylation, ubiquitination, and sumoylation, may be an important factor affecting the cloning efficiency (Xu et al., 2009). Thus, more studies were focused on histone acetylation modification.
It is thought that epigenetic modification accumulated in the donor nuclei following SCNT can be fully reprogrammed into the totipotent state of preimplantation embryos. Acetylation and deacetylation, catalyzed by specific enzymes, including histone acetyltransferases (HATs) and histone deacetyltransferases (HDACs), alter the higher-order chromatin structure to regulate transcriptional machinery (Lee et al., 1993). Considering that the reprogramming of nuclei following nuclear transfer only happens during a limited time, the relaxation of chromatin structure by histone acetylation, which maintains a transcriptionally permissive state, might improve the cloning effciency. Many drugs have been used to regulate histone acetylation modification, such as trichostatin A (TSA) (Hai et al., 2011; Iager et al., 2008; Kishigami et al., 2006), sodium butyrate (NaBu) (Das et al., 2010), valproic acid (Xu et al., 2012), and Scriptaid (Van Thuan et al., 2009; Zhao et al., 2009, 2010). It has been shown that TSA could significantly improve the developmental ability of bovine SCNT embryos by correcting the pattern of acetylation modification in H4K5 and decreasing the expression level of histone deacetylases (HDAC1 and HDAC2) (Lee et al., 2011). Some reports indicated that treatment of embryos derived from SCNT with TSA could raise the level of lysine acetylation to the level of normal embryos in mouse (Wang et al., 2007), pig (Li et al., 2008a), and rabbit (Shi et al., 2008).
In addition, treatment of SCNT embryos with TSA decreased the expression of genes related to DNA methylation and increased the expression of genes related to pluripotency in mouse blastocysts (Li et al., 2008b). However, some reports suggested that TSA treatment had detrimental effects on the development of SCNT embryos and resulted in low developmental competency and even teratogenic cloning offspring (Svensson et al., 1998). Wu et al. (2008) reported that treatment of donor cells with TSA resulted in significantly lower blastocyst development in bovine SCNT. The offspring developed from SCNT embryos treated with TSA died within a month, whereas four rabbit pups from untreated embryos grew into adulthood (Meng et al., 2009).
On the basis of the detrimental effects of TSA on SCNT embryos, we chose a relatively low-toxicity HDAC inhibitor, Scriptaid, which had been proved to enhance transcriptional activity and protein expression to treat buffalo SCNT embryos (Su et al., 2000). It has been reported that treatment of cloned embryos with Scriptaid not only increased the histone acetylation level on H4K8 (Zhao et al., 2010), but also improved the cloning efficiency in inbred miniature pig (Zhao et al., 2009) and inbred mice (Van Thuan et al., 2009), in which the overall cloning efficiency was increased from 0% to 3.7%. Meanwhile, Whitworth et al. (2011) reported that the expression levels of three transcripts (H3F3A, CAPG, and SEPT7) of embryos treated with Scriptaid returned to the similar level of porcine in vivo embryos, and the global DNA methylation level in blastocysts developed from SCNT embryos treated with Scriptaid also was reduced.
Therefore, the present study was undertaken to investigate the effects of Scriptaid treatment on the in vitro and in vivo developmental abilities of buffalo SCNT embryos. Moreover, the levels of histone H3K18 acetylation and global DNA methylation as well as the expression profiles of genes related to histone acetylation and DNA methylation in buffalo SCNT embryos treated with and without Scriptaid were also detected.
Materials and Methods
Unless otherwise stated, all chemicals used in this study were purchased from Sigma Chemical Company (St. Louis, MO, USA). All of the culture medium was sterilized by passing through a 0.22-μm filter (Millipore).
In vitro maturation of oocytes
Buffalo ovaries were obtained from a local slaughterhouse and transported to the laboratory at 37°C in saline solution within 4 h. Cumulus–oocyte complexes (COCs) were collected by aspiration of ovarian antral follicles (2–7 mm diameter) with a 12-gauge needle fixed to a 10-mL syringe. The COCs with intact layers of cumulus cells were selected from cellular debris and washed three times in a washing medium consisting of Tissue Culture Medium-199 (TCM-199), 5.0 mM NaHCO3, 5 mM HEPES, and 2% fetal bovine serum (FBS). After washing, COCs were cultured in a 10-mm glass dish containing the maturation medium for 22–24 h under a humidified 5% CO2-in-air atmosphere at 38.5°C. The maturation medium consisted of TCM-199, 26.2 mM NaHCO3, 5 mM HEPES, 0.1 μg/mL follicle-stimulating hormone (FSH; Hyclone, Logan, UT, USA), 5% FBS (Hyclone), 60 mg/L penicillin G, and 100 mg/L streptomycin sulfate.
Donor cell preparation
Tissues from the skin of a buffalo female fetus (about 3 months) were manually cut into small pieces after sterilizing with 70% alcohol and digested enzymatically with 0.25% trypsin and 0.05% EDTA for 30 min, and then 0.2% collagenase for 45 min. The disaggregated cells were washed three times in Dulbecco's Modified Eagle Medium (DMEM, Gibco) supplemented with 10% FBS by centrifugation at 1500 rpm for 5 min and cultured in a 60-mm culture dish under a humidified 5% CO2-in-air atmosphere at 38.5°C. After 7–10 days of culture, confluent fibroblast monolayers were obtained and then routinely passaged with an enzymatic solution (0.25% trypsin and 0.05% EDTA) for 5 min. After four to seven passages, these cells were used as donor cells for nuclear transfer. Before nuclear transfer, cells were treated with an enzymatic solution for single-cell isolation.
Nuclear transfer, fusion, and activation
Nuclear transfer was performed according to the method reported by us previously (Shi et al., 2007) with some modifications. Briefly, trypsinized donor cells were transferred into the perivitelline space of enucleated oocytes with a 25-μm micropipette. The reconstructed complexes were transferred to a 25 μL droplet of fusion medium [0.28 M mannitol, 0.1 mM CaCl2, 0.1 mM MgSO4, 5 mM HEPES, and 0.1% bovine serum albumin (BSA)] covered with mineral oil and then placed on the micromanipulator with two platinum needle electrodes. Cell fusion was induced with an alternating current pulse of 2 V for 1 sec and then three direct current pulses of 1 kV/cm for 15 μsec by an ECM-2001 Electrocell Manipulator (BTX Inc., CA, USA). Couplets were then incubated in culture medium (CM) (TCM-199, 3% FBS, CM) for 30 min at 38.5°C. The fusion of couplets was checked under an inverted microscope. The activation of reconstructed embryos was induced by exposure to 5 μM ionomycin in CM for 5 min and immediately incubation in 2 mM 6-dimethylaminopurine in CM for 3 h.
In vitro fertilization
For in vitro fertilization (IVF), the frozen semen straw (0.25 mL per straw) was gently shaken at room temperature in air for 10 sec and then thawed for 10 sec in a 37°C water bath. The thawed semen was layered under 1.5 mL of modified Tyrode's medium in a conical tube for the swim-up procedure. After incubation for 30 min at 38.5°C, the top 0.8–1.0 mL of medium containing the more motile sperm was collected and centrifuged for 5 min at 1500 rpm to harvest the pelleted sperm. The spermatozoa pellet was resuspended in modified Tyrode's medium at a concentration of 5.0×106 sperm/mL for fertilization. After in vitro maturation of 24 h, oocytes were washed twice in modified Tyrode's medium and transported into a droplet of 30 μL modified Tyrode's medium under sterile mineral oil (20 oocytes per drop). Then, 4–5 μL of semen was added to the droplet containing oocytes to give a final sperm concentration at 1–1.5×106 sperm/mL. This was incubated for 24 h at 38.5°C under a humidified 5% CO2-in-air atmosphere.
In vitro culture of embryos
Embryos derived from IVF and SCNT were placed in co-culture with granulosa cell monolayers in a 30-μL droplet of CM overlaid with mineral oil under a humidified 5% CO2-in-air atmosphere at 38.5°C. The granulosa cell monolayers were established at 48–72 h before introduction of embryos. After introduction of embryos, half of the medium was replaced with fresh medium every 24 h. At 48 h of co-culture, the cleavage rate of embryos was checked, and the number of developed blastocysts was recorded within 8 days of co-culture.
Embryo transfer
Day 6 blastocysts developed from SCNT embryos treated with or without Scriptaid were transferred nonsurgically into the uterine horn ipsilaterally to the ovary containing a palpable corpus luteum of recipient Chinese swamp buffalos at day 6 of the native estrous cycle. Each recipient received two embryos. The pregnancy status was determined on 60 days after embryo transfer by rectal palpation.
Immunohistochemistry
Embryos at different developmental stages (two-cells, eight-cells, morula, and blastocyst) were fixed in 4% paraformaldehyde for at least 30 min at room temperature. Subsequently, embryos were washed three times in phosphate-buffered solution (PBS) supplemented with 0.01% Triton X-100 and 0.3% BSA (TBP) and then permeabilized with 1% Triton X-100 in PBS for 10 min at room temperature. Then, the embryos were incubated with 1% BSA in PBS for 1 h. After washing three times with PBS, samples were incubated with primary antibody against histone H3 acetylated lysine 18 (1:100, rabbit, H3K18; Upstate Biotechnology), 5-methylcytidine (5-mC; mouse, 1:300; Abcam, Cambridge, UK) overnight at 4°C. Thereafter, embryos were washed three times in TBP and incubated with fluorescein isothiocyanate (FITC)-conjugated second antibody [goat anti-rabbit immunoglobulin G (IgG), 1:200, Millipore; goat anti-mouse IgG, 1:1500, CST] for 2 h at room temperature in the dark. Meanwhile, nuclei of embryos were counterstained with 10 μg/mL propidium iodide (PI) for 5 min. Samples were mounted on slides with a drop of anti-fade solution (Fluoromount-GTM, SouthernBiotech, Birmingham, AL, USA) and analyzed under a LSM 510 META Laser Scanning Microscope (Zeiss, Germany) with Zeiss image software. Measurements were performed in three replicates for embryos at each developmental stage with at least 10 embryos. IVF embryos at the two-cell stage were set as reference.
Embryo collection and reverse transcription
Five excellent embryos at the eight-cell or blastocyst stage were collected and treated using a Cells-to-cDNA™ II Kit (Ambion Co., Australia) according to the manufacturer's instructions with some modifications. Embryos were washed three times with PBS and then incubated with 8 μL of cell lysis II for 15 min at 75°C. The genomic DNA was digested in 1 μL of DNase I (Fermentas) at 37°C for 30 min, and the DNase was inactivated with 1 μL of EDTA at 65°C for 15 min. The reverse transcription was performed according to the SuperScript™ II Reverse Transcriptase (Invitrogen) instructions. The reaction system consisted of 4 μM random primer (TaKaRa), 10 mM mixture deoxynucleotides (dNTPs; TaKaRa), 10 IU of RNase inhibitor (TaKaRa), 4 μL of 5× First-Strand Buffer, 2 μL of dithiothreitol (DTT), and 0.25 μL of Fast Reverse Transcriptase. The reaction mixture was incubated at 25°C for 10 min, 42°C for 90 min, and 95°C for 10 min to inactivate the reverse transcriptase (RT) enzyme. Finally, sterile H2O was added to adjust the final volume of cDNA to 0.2 μL per embryo. The cDNA was stored at −80°C until use.
Quantitative real-time polymerase chain reaction
The quantification of all gene transcripts was carried out in an ABI 7500 Fast Real-Time System (Applied Biosystems), and primers were designed by the Oligo 6.0 software (Table 1). The β-actin of housekeeping gene was used as the internal control, and reaction mixture in each well included 10 μL of SYBR Premix Ex Taq™ (TaKaRa), 0.3 μL of each primer (10 nM), 0.4 μL of ROX Reference Dye II (50×), 1 μL of cDNA, and 8.3 μL of H2O (total volume of 20 μL). The thermal cycle was 95°C for 3 min (denaturation) followed by 40 cycles of 95°C for 30 sec, 55°C for 30 sec, and 72°C for 30 sec. Each detection was repeated three times using different samples for each gene. The expression level of target genes was determined using the formula 2−ΔΔCT.
Experimental design
Experiment 1
This experiment was designed to examine the effect of treatment of buffalo SCNT embryos with different concentrations of Scriptaid on their in vitro development. After activation, reconstructed embryos were treated with 250, 500, or 750 nM of Scriptaid for 12 h, and then placed into CM for in vitro culture. Scriptaid was dissolved in dimethyl sulfoxide (DMSO) at 5 mM and then added to the medium at working concentrations in which the concentration of DMSO was less than 0.02%. The cleavage rate of embryos was checked at 48 h of culture, and the number of blastocysts developed was recorded within 8 days of culture.
Experiment 2
Effects of Scriptaid treatment time on the in vitro development of buffalo SCNT embryos were examined in this experiment. After activation, reconstructed embryos were treated with 500 nM Scriptaid for 3 h, and then placed into CM containing 500 nM Scriptaid for cultures of 6, 12, 18, 24, 30, or 36 h. After Scriptaid treatment, the reconstructed embryos were placed into CM for in vitro culture.
Experiment 3
Effects of Scriptaid treatment on the histone acetylation and DNA methylation of SCNT embryos were investigated in this experiment. Buffalo embryos that had been treated with 500 nM Scriptaid for 24 h were collected for qRT-PCR and immunohistochemistry analysis at the two-cell, eight-cell, morula, and blastocyst stages. The H3K18 acetylation and global DNA methylation was determined by immunohistochemical analysis, and the expression of genes related to histone acetylation (CBP, p300, and HAT1) and DNA methylation (Dnmt1 and Dnmt3a) was determined by qRT-PCR. The embryos developed from the nontreated SCNT and IVF embryos were set as the control.
Experiment 4
This experiment was designed to examine the effect of Scriptaid treatment on the in vivo development of SCNT embryos. Blastocysts developed from buffalo SCNT embryos treated with or without 500 nM Scriptaid for 24 h were transferred into recipient buffalos, and the pregnancy status was determined on 60 days after embryo transfer by rectal palpation.
Statistical analysis
Experiments were repeated at least three times. The difference between treatment in the embryonic development, H3K18 acetylation and global DNA methylation, and expression profiles of the target genes were analyzed using one-way repeated-measures analysis of variance (ANOVA) and SPSS 17.0 software. The pregnancy rate of recipients was also analyzed using independent samples t-test. Probability values less than 0.05 were considered statistically significant.
Results
Effects of treating buffalo SCNT embryos with different concentrations of Scriptaid on their subsequent development in vitro
As shown in Table 2, treatment of SCNT embryos with 500 nM Scriptaid for 12 h resulted in a significantly increase (p<0.05) in the blastocyst development (19.2%) in comparison with SCNT embryos treated with 0 (10.3%), 250 nM (12.8%), and 750 nM (12.6%) Scriptaid for 12 h, although there was no significant difference in the cleavage rates among the groups.
Values with different superscripts in the same column are significantly different (p<0.05).
SCNT, somatic cell nuclear transfer; SEM, standard error of the mean.
In vitro development of buffalo SCNT embryos treated with Scriptaid for different times
As shown in the Table 3, the cleavage rate of SCNT embryos treated with 500 nM Scriptaid for 18 h (74.5%) was significantly higher than that of embryos treated for 30 h (56.6%, p<0.05). More SCNT embryos treated with Scriptaid for 24 h developed to blastocysts (28.2%) in comparison with SCNT embryos treated 500 nM Scriptaid for 0, 6, and 12 h (13.6%, 15.5%, 17.4%, p<0.05).
Values with different superscripts in the same column are significantly different (p<0.05).
SCNT, somatic cell nuclear transfer; SEM, standard error of the mean.
Effects of treating SCNT embryos with Scriptaid on their histone acetylation and DNA methylation
To examine the effects of Scriptaid treatment on histone acetylation and DNA methylation of SCNT embryos, H3K18 acetylation and global DNA methylation in SCNT and IVF embryos at different stages were measured by immunostaining. As shown in Figure 1 and Figure 3A(see below), the intensity of histone H3K18 signals in the IVF embryos decreased from the two-cell stage to the morula stage and increased to a peak at the blastocyst stage. In contrast, the relative levels of global DNA methylation increased from the two-cell to blastocyst stage (Fig. 2 and Fig. 3B, p<0.05). In the SCNT embryos, the intensity of H3K18 acetylation signals at the different developmental stages (two-cell, eight-cell, morula, and blastocyst) was lower than in the IVF embryos, and the relative levels of global DNA methylation were higher than in the IVF embryos (p<0.05). However, treatment of SCNT embryos with 500 nM Scriptaid for 24 h resulted in an increase in the intensity of H3K18 signals and a decrease in the relative levels of global DNA methylation in comparison with the untreated group and similar to fertilized counterparts.
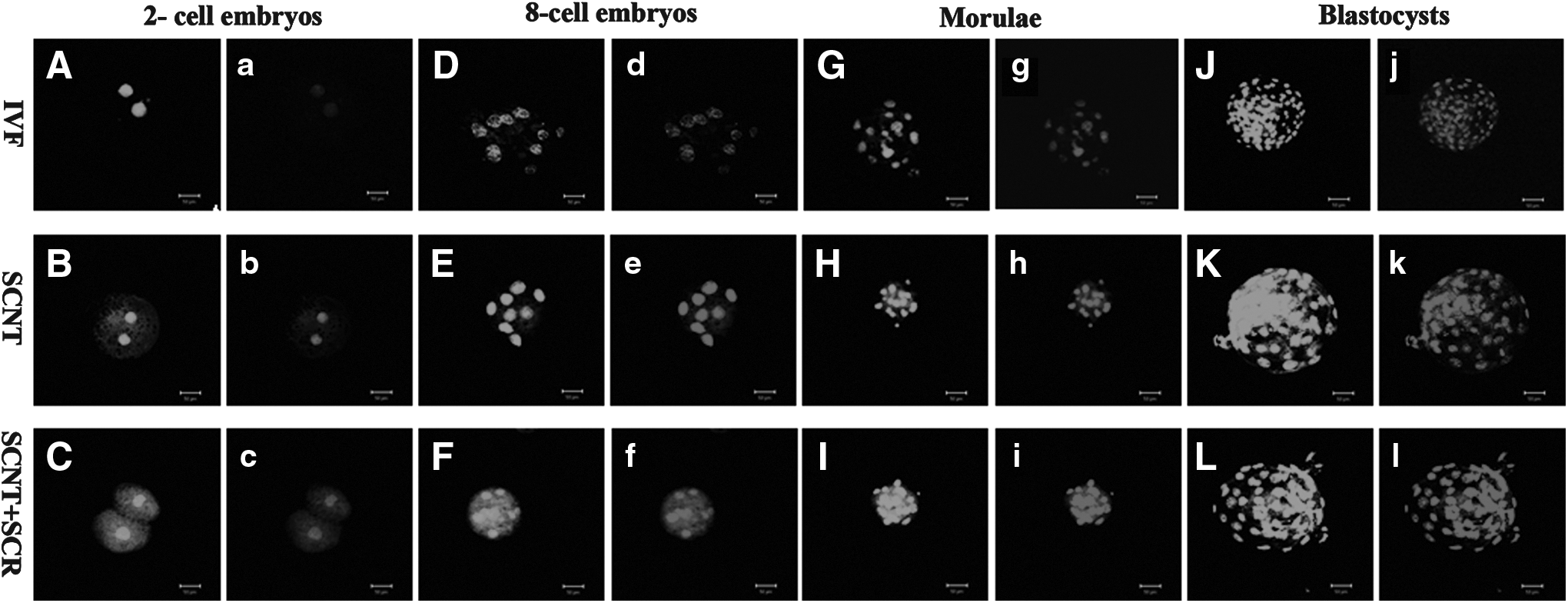
Acetylation pattern of histone H3 on lysine 18 (H3K18) in SCNT, Scriptaid-treated SCNT (SCNT+SCR), and IVF embryos at the two-cell (
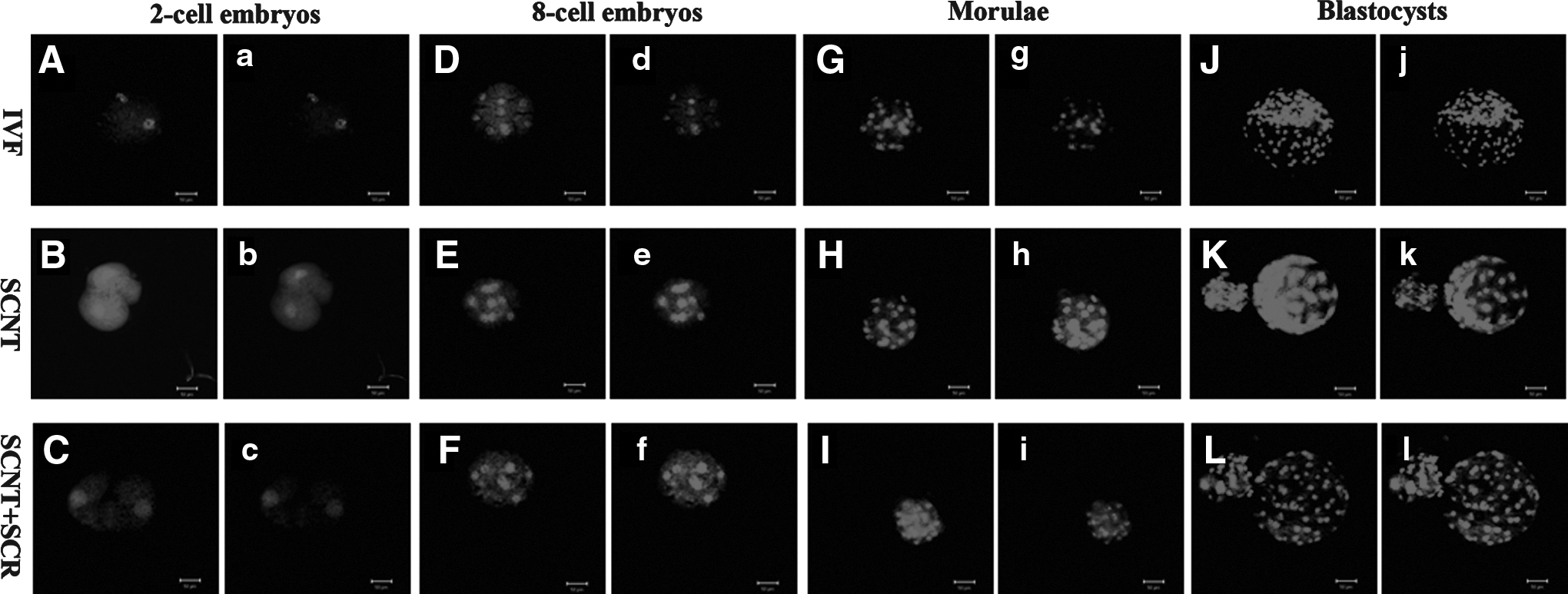
Level of global DNA methylation in SCNT, Scriptaid-treated SCNT (SCNT+SCR), and IVF embryos at the two-cell (
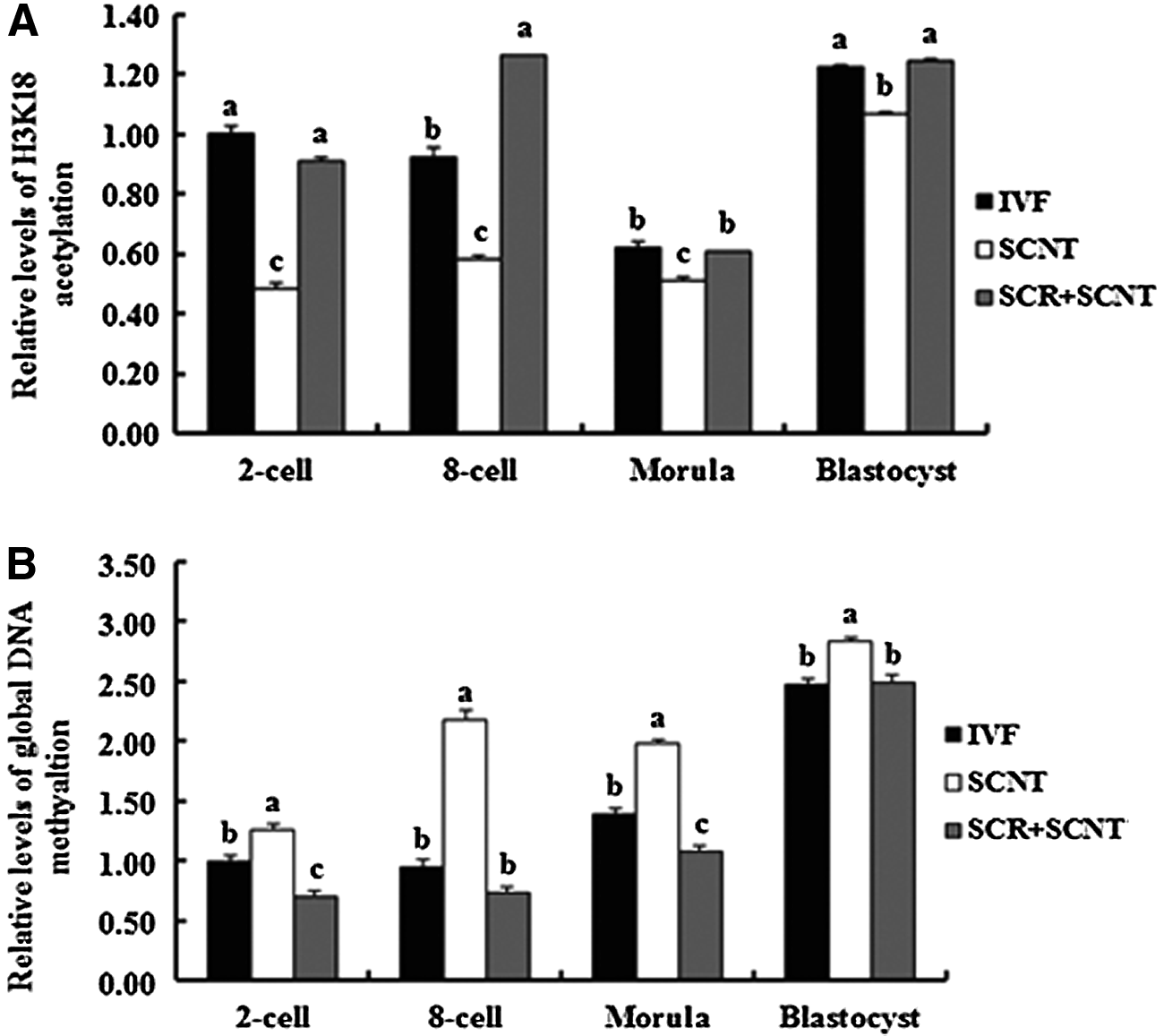
Relative levels of H3K18 acetylation (
Meanwhile, the expression of genes related to histone acetylation (CBP, p300, and HAT1) and DNA methylation (Dnmt1 and Dnmt3a) in the embryos at the eight-cell and blastocyst stages was also analyzed by real-time PCR. As shown in Figures 4 and 5, treatment of SCNT embryos with Scriptaid resulted in a decrease in the expression levels of genes related to histone acetylation (CBP, p300, and HAT1) and DNA methylation (Dnmt1 and Dnmt3a) at the eight-cell stage (Fig. 4A–C, Fig. 5A; p<0.05), which was similar to the IVF embryos. However, the expression profile of HAT1 and Dnmt1 increased remarkably at the blastocyst stage (Fig. 4F, Fig. 5C; p<0.05), and the increase of Dnmt3a expression was prohibited by Scriptaid treatment (Fig. 5D, p<0.05). A similar expression profile of HAT1, Dnmt1, and Dnmt3a was found in blastocysts developed from Scriptaid-treated SCNT embryos and IVF embryos (Fig. 4D, E; p>0.05).
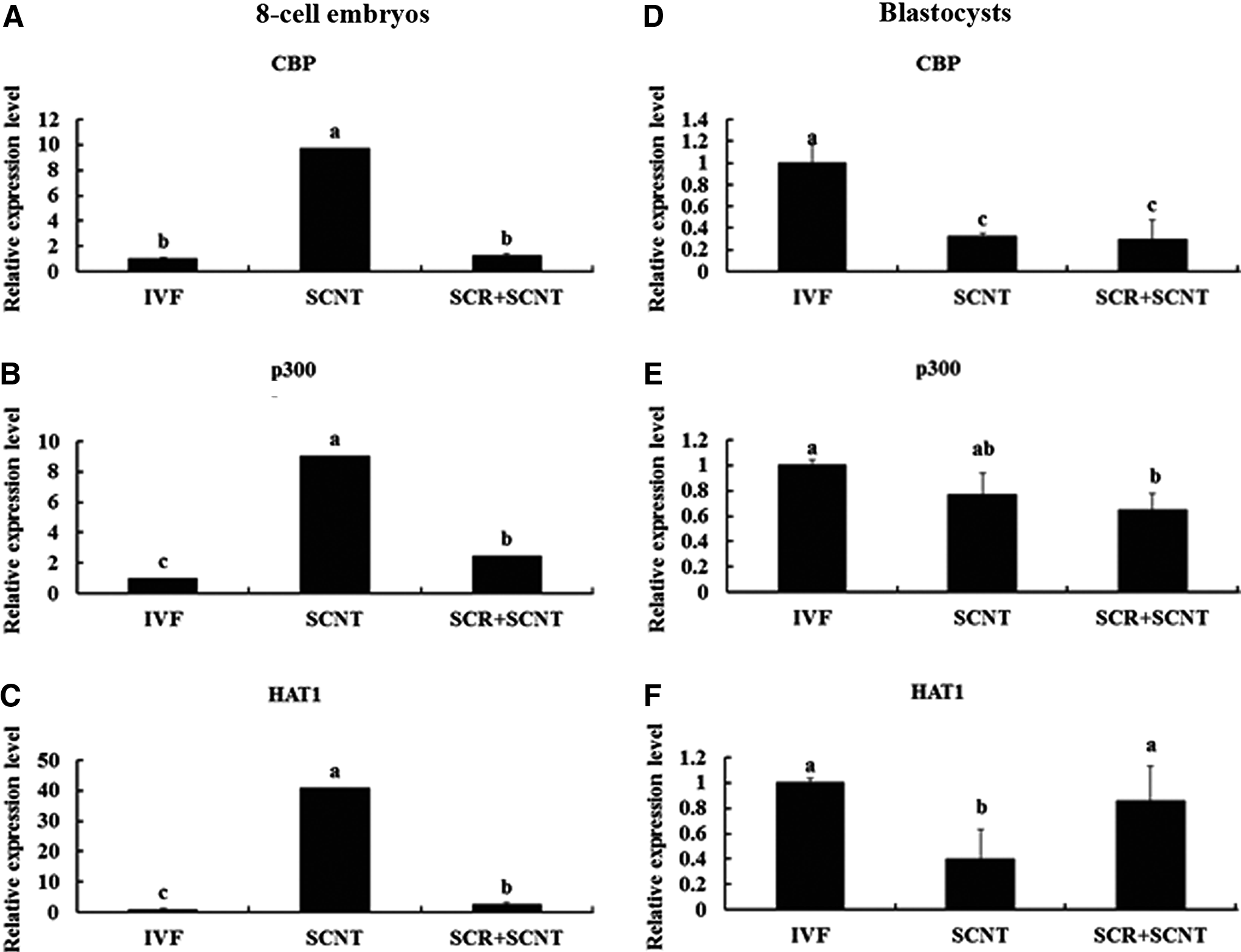
Relative expression levels of genes related to histone acetylation (CBP, p300, and HAT1) in SCNT, Scriptaid-treated SCNT (SCR+SCNT), and IVF embryos at the eight-cell (
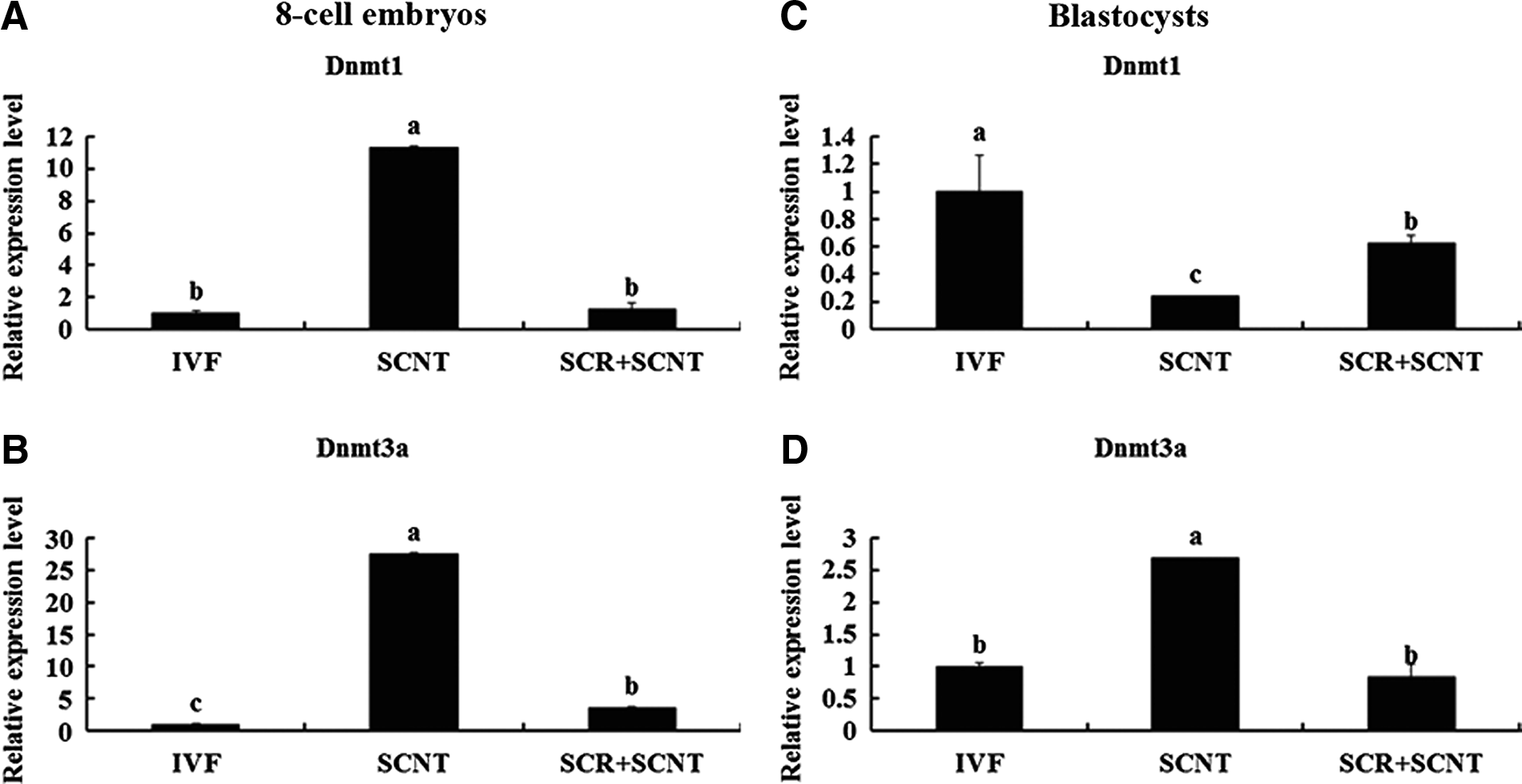
Relative expression profiles of genes related to DNA methylation (Dnmt1 and Dnmt3a) in SCNT, Scriptaid-treated SCNT (SCR+SCNT), and IVF embryos at the eight-cell (
Effects of Scriptaid treatment on the in vivo development of SCNT embryos
Blastocysts developed from 500 nM Scriptaid-treated and nontreated SCNT embryos were transferred nonsurgically into recipients. The pregnancy status of recipients was examined by rectal palpation at 60 days after embryo transfer (Table 4). Only one of nine recipients was pregnant in the nontreated SCNT group, and 11 of 48 recipients were confirmed to be pregnant following transfer of blastocysts derived from the SCNT embryos treated with 500 nM Scriptaid.
SCNT, somatic cell nuclear transfer.
Discussion
Although many species have been cloned through the SCNT technique, the overall efficiency is quite low (Rideout et al., 2001). Many studies have showed that the developmental defects of cloned embryos fail to eliminate epigenetic-specific markers completely and establish the related gamete markers themselves (Kishigami et al., 2006; Latham, 2005; Rideout et al., 2001; Shi et al., 2003). Moreover, many studies also found that an abnormal epigenetic modification might result in the developmental failure of SCNT embryos, such as abnormal histone acetylation and DNA methylation in reprogramming (Bohrer et al., 2014; Enright et al., 2005; Jafarpour et al., 2011; Sangalli et al., 2014). Histone acetylation is a part of epigenetic modification (Grunstein, 1997; Turner, 2002) and is related to transcriptional activation by regulating the chromosome structure. A high histone acetylation level leads to the relaxation of chromatin structure and formation of a transcriptionally permissive state and then regulates the transcription of genes related to embryonic development (Berger, 2002; Lee et al., 1993; Zlatanova et al., 2000). As such, DNA methylation is also a key epigenetic reprogramming factor that modifies and regulates chromatin structure, which may be critical for sustaining genomic stability, regulating genomic imprinting, and activating or suppressing gene expression (Lan et al., 2010). Thus, histone acetylation and DNA methylation play important roles in epigenetic modification and display remarkable variability during embryonic development. Dynamic interactions between histone acetylation and DNA methylation are thought to regulate gene expression. Thus, an effort was made to improve the development of buffalo SCNT embryos by treatment with Scriptaid, a histone deacetylase inhibitor.
It has been reported that the level of histone acetylation in cloned embryos is lower than in fertilized embryos (Iager et al., 2008; Zhao et al., 2010), which may be related to the low efficiency of SCNT (Enright et al., 2003b). Thus, most histone deacetylase inhibitors had been used to improve SCNT efficiency by increasing histone acetylation levels. Scriptaid, a deacetylase inhibitor with lower toxicity (Van Thuan et al., 2009), can greatly increase the level of histone acetylation and gene transcription (Su et al., 2000) and had been proved to improve the development of SCNT embryos in miniature pigs (Zhao et al., 2009) and cattle (Akagi et al., 2011). In the present study, treatment of buffalo SCNT embryos with 500 nM Scriptaid for 24 h resulted in an increase in blastocyst development and pregnancy rate following embryo transfer. These results were similar to the Zhao et al. (2010) report in pigs and Panda et al. (2012) report in buffalos.
The levels of histone H3K18 acetylation and global DNA methylation are normally used as the marker of epigenetic reprogramming in SCNT embryos. In the present study, treatment of buffalo SCNT embryos with Scriptaid resulted in an increase in H3K18 acetylation and a similar acetylation pattern to the IVF group. It has been reported that treatment of porcine SCNT embryos with Scriptaid resulted in an increase in their overall cloning efficiency and histone H4K8 acetylation level (Zhao et al., 2010). Meanwhile, the acetylation levels of H3K4, H3K9, H3K14, H3K18, H4K5, H4K8, and H4K16 in SCNT embryos were also increased following Scriptaid treatment (Cervera et al., 2009; Iager et al., 2008; Wang et al., 2007; Xu et al., 2012; Yamanaka et al., 2009). These reports are consistent with our results. Furthermore, treatment of buffalo SCNT with Scriptaid also resulted in a decrease in the level of global DNA methylation and a similar DNA methylation pattern in blastocysts to the IVF group, which was consistent with the report in porcine SCNT embryos (Whitworth et al., 2011). The DNA methylation level and histone acetylation may be related each other, and the molecular mechanism linked them will be studied further.
It is well known that acetylation and deacetylation are catalyzed by HATs and HDACs, respectively, which are closely related to transcription activity (Grunstein, 1997). HATs are divided into two types, A and B. Type A HATs are found in the nucleus and mainly regulate gene expression by functioning as transcriptional co-activators (Brownell and Allis, 1996). Type B HATs are located mainly in the cytoplasm and may function in chromatin assembly (Ruiz-García et al., 1998). Many HATs have a co-activating role for different transcriptional factors, including the CREB-binding protein (CBP) and p300 (Dokmanovic and Marks, 2005). It has been reported that transcriptional repression of selective cellular genes was released via HDAC mechanisms, which include recruitment of the co-activator CBP/p300 and P300/CBP-associated factor (PCAF) (Hernandez et al., 2006). Li et al. (2008b) reported that the expression levels of CBP and p300 in cloned mouse blastocysts were higher than in the in vivo and IVF embryos. In our study, treatment of buffalo SCNT embryos with Scriptaid decreased the expression profiles of CBP, p300, and HAT1 at the eight-cell stage, which was similar to the IVF embryos. However, the expression level of HAT1 at the blastocyst stage in Scriptaid-treated embryos was significantly higher than in the nontreated embryos. This result indicates that HAT1 may be associated with the increase in metabolism of embryos at the blastocyst stage.
DNA methylation is regulated by two types of DNA methyltransferases—Dnmt1 and Dnmt3a and Dnmt3b. Dnmt1 is mainly involved in maintaining the methylation pattern during DNA replication (Bestor, 2000). Dnmt3a and Dnmt3b, de novo methyltransferases, are responsible for establishing the initial patterns of methylation during embryogenesis (Okano et al., 1999). In cloned mouse blastocysts, the expression of Dnmts was higher than the in vivo embryos (Li et al., 2008b). Thus, decline the expression level of Dnmts in cloned embryos to the level of in vivo embryos may improve their developmental competence. In the present study, the expression of Dnmt3a in SCNT embryos treated with Scriptaid significantly decreased at the eight-cell and blastocyst stage in comparison with the control group and nearly reached to the levels of the IVF group. This result was coincident with the report by Whitworth et al. (2011), who observed that porcine cloned embryos treated with Scriptaid displayed significantly lower global DNA methylation at the blastocyst stage compared to the control. The low expression profile of Dnmt3a caused by Scriptaid treatment may result in the activation of genes that is essential for embryonic development.
Although more buffalo recipients that received blastocysts developed from SCNT embryos treated with Scriptaid became pregnant (22.9%) in comparison with SCNT untreated embryos (11.1%), the statistical difference was not significant (p=0.3459). Thus, more buffalo SCNT embryos treated with Scriptaid should be transferred into recipients to further verify the beneficial effect of Scriptaid on the development of buffalo SCNT embryos in future.
In conclusions, treatment of buffalo SCNT embryos with Scriptaid can improve their developmental competence. This action is mediated by increasing the H3K18 acetylation level and decreasing the global DNA methylation level, resulting in a similar expression pattern of genes related to histone acetylation and DNA methylation compared to IVF embryos.
Footnotes
Acknowledgments
This work was funded by the China High Technology Development program (grant no. 2011AA100607), China Transgenic Project (grant no. 2014ZX08007-001), Guangxi Bagui Scholar Program, China Natural Science Foundation (grant no. 31072033), and Guangxi Science Foundation (grant no. 2012GXNSFFA060004).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.