
Abstract
Abstract
Histone modification plays an important role in maintaining pluripotency and self-renewal of embryonic stem cells (ESCs). The histone acetyltransferase MOF is a key regulator of ESCs; however, the role of MOF in the process of reprogramming back to induced pluripotent stem cells (iPSCs) remains unclear. In this study, we investigated the function of MOF on the generation of iPSCs. We show that iPSCs contain high levels of MOF mRNA, and the expression level of MOF protein is dramatically upregulated following reprogramming. Most importantly, overexpression of MOF improves reprogramming efficiency and facilitates the formation of iPSCs, whereas small hairpin RNA (shRNA)-mediated knockdown of MOF impairs iPSCs generation during reprogramming. Further investigation reveals that MOF interacts with the H3K4 methyltransferase Wdr5 to promote endogenous Oct4 expression during the reprogramming process. Knockdown of MOF reduces H4K16ac and H3K4me3 modification at the Oct4 promoter. In conclusion, our data indicate that MOF is an important epigenetic regulator that is critical for efficient reprogramming.
Introduction
I
Histone H3 and H4 acetylation levels increase in the promoter region of pluripotent genes during the reprogramming process, which may be one of the key epigenetic factors controlling the reprogramming of somatic cells (Mattout et al., 2011). The reprogramming factor c-Myc could regulate the chromosome structure by recruiting histone acetyltransferase in iPSCs generation. In addition, low levels of histone H3K9 acetylation (H3K9ac) could reduce the efficiency in cell fusion-mediated reprogramming (Hezroni et al., 2011). Histone deacetylase (HDAC) inhibitors, including valproic acid (Huangfu et al., 2008a, b) and butyrate (Liang et al., 2010; Mali et al., 2010), could enhance reprogramming efficiency by promoting epigenetic remodeling and the expression of pluripotency-associated genes. Despite this evidence demonstrating that histone acetylation plays an important role in reprogramming, it is surprising that few reports have identified the function of histone acetylation in regulating pluripotency.
MOF (males absent on the first; also known as MYST1 or KAT8) belongs to the histone acetyltransferase MYST (MOZ-Ybf2/Sas3-Sas2-Tip60) family. The family contains a highly conserved MYST domain, which is responsible for the acetyltransferase activity (Awakumov and Côté, 2007). MOF as a catalytic subunit could cause histone H4K16 acetylation (Neal et al., 2000). MOF resides in two distinct complexes—the MOF–MSL complex and the MOF–NSL complex. The MOF–NSL complex could interact with the MLL complex through the commonly shared component Wdr5 and strengthen transcriptional activation of the MLL complex (Dou et al., 2005; Li et al., 2009; Li and Dou, 2010). In mammals, MOF is essential for vertebrate development. Ablation of MOF leads to peri-implantation lethality in mouse embryos (Gupta et al., 2008; Thomas et al., 2008). A recent study shows that MOF deletion leads to loss of ESCs self-renewal and defects in embryoid body (EB) formation (Li et al., 2012). However, the current study did not further study the role of MOF in the reprogramming process.
In this study, we investigated the impacts of MOF on iPSCs generation. We show that MOF is highly enriched in iPSCs and ESCs, and MOF expression is upregulated during the reprogramming process. The ectopic expression of MOF promotes reprogramming, whereas knockdown of MOF suppresses iPSCs generation, possibly by affecting the H3K4 methyltransferase Wdr5 and endogenous Oct4 expression. The results demonstrate that MOF may be a potential epigenetic regulator that facilitates iPSCs generation.
Materials and Methods
Cells, media, and plasmids
Human primary foreskin fibroblasts (hFFs) were obtained from Millipore. Mouse embryonic fibroblasts (MEFs) were isolated from CD-1 (ICR) mice. hFFs, MEFs, and 293T cells were cultured in growth medium consisting of Dulbecco's Modified Eagle Medium (DMEM; Invitrogen), 10% fetal bovine serum (FBS), 50 U/mL penicillin, and 50 mg/mL streptomycin. Human ESC (hESC) line H1 and iPSCs were maintained on mitomycin C (MMC)-inactivated MEFs feeder layers in hESCs medium. The hESCs medium was Knockout-DMEM (Invitrogen), supplemented with 10% Knockout Serum Replacement (KSR), 10 ng/mL basic fibroblast growth factor (bFGF; Millipore), 1 mM
The hESCs medium was refreshed daily, and cells were passaged every 5–7 days enzymatically with 1.5 mg/mL collagenase type IV (Invitrogen). pMIG retrovirus vectors with reprogramming factors Oct4, Sox2, Klf4, and c-Myc were used for iPSCs generation (Park et al., 2008). The pMIG vector has an internal ribosome entry site–green fluorescent protein (IRES-GFP) cassette, which could express GFP. Plasmids encoding Oct4, Sox2, Klf4, and c-Myc, MOF, and Wdr5 were obtained from Addgene. Retroviral small hairpin RNA (shRNA) vectors based on pQsupR were obtained from Addgene. The nucleotide sequences of MOF shRNA were AAAGACCAUAAGAU UUAUU.
All animal experiments were approved by the Animal Welfare and Research Ethics Committee of Jilin University. Mice were housed in a temperature-controlled room with proper dark–light cycles, fed with a regular diet. All efforts were made to minimize suffering.
Virus packaging and iPSCs induction
For virus packaging, retrovirus expressing Oct4, Sox2, Klf4, c-Myc, MOF, Wdr5, or MOF shRNA were produced by transfecting retroviral vectors with packaging plasmids (pUMVC and VSVG) into 293T cells using Lipofectamine™ LTX Reagent (Invitrogen) according to the manufacturer's recommendations. After 48 h, the viral supernatants were collected and filtered through a 0.45-μm filter. The premade viral supernatants were titrated in 293T cells and supplied at 1×106 IU/mL in DMEM medium. For hFFs, 2 mL/per well of viral supernatants in a six-well plate could reach the optimal multiplicity of infection.
For iPSCs induction, 5×104 hFFs (P3) were seeded in a six-well dish 24 h before transduction. hFFs were infected with the respective retroviruses supplemented with 8 μg/mL Polybrene (Sigma). After infection twice, the cells were passaged onto MEF feeders and the medium was changed to hESCs medium. The medium was changed every other day. Colonies with ESCs-like morphology became visible about 3 weeks after infection. Colonies were picked and expanded on MEFs feeder layers using standard procedures.
Alkaline phosphatase staining
The ESCs-like colonies that formed were fixed in 4% paraformaldehyde (PFA; Sigma) for 15 min at room temperature. Cells were washed with phosphate-buffered saline (PBS) and incubated with the staining solution according to the manufacturer's instructions (Beyotime). The number of alkaline phosphatase–positive (AP+) colonies was scored.
Embryoid body and teratoma formation from human iPSCs
For in vitro differentiation, iPSCs were harvested and incubated in hESCs medium without bFGF for 9 days to permit EB formation. The medium was changed every other day. Then EBs were plated onto gelatin-coated plates to facilitate adherent culture. After 8 days, differentiation was determined by immunostaining with antibodies against Pdx1 (1:100; Millipore), troponin (1:100; Millipore), or nestin (1:100; Millipore).
For teratoma formation assay, 1×106 iPSCs were injected subcutaneously into nude mice. Approximately 4 weeks later, teratomas were surgically dissected, fixed in 4% PFA, and embedded in paraffin. Then paraffin-embedded tissues were sliced and subjected to Hematoxylin & Eosin (H&E) staining to detect in vivo differentiation.
RNA isolation and RT-PCR analysis
RNA was extracted using TRIzol reagent (Invitrogen) and purified using an RNeasy Kit (Qiagen). Quantified RNA (1 μg) was used to produce cDNA in the presence of oligo(dT). cDNA was then used for PCR amplification by Taq DNA polymerase (TaKaRa). The mRNA levels of the target genes were quantified by real-time PCR using SYBR Premix Ex Taq™ II, as described (TaKaRa), in a Mastercycler ep realplex2S Real-time PCR system (Eppendorf). PCR reactions were performed under the following program: 95°C for 2 min, 40 cycles of 95°C for 30 sec, 58°C for 30 sec, and 72°C for 30 sec, and then a melting curve of the amplified DNA was acquired. Primers are listed in supporting information Table S1 (Supplementary Data are available at www.liebertpub.com/cell/). Quantification of target genes was normalized with β-actin.
Chromatin immunoprecipitation assay
Cells were crosslinked with 1% formaldehyde for 15 min at room temperature. Fixed cells were lysed in lysis buffer [1% sodium dodecyl sulfate (SDS), 10 mM EDTA, 50 mM Tris-HCl, pH 8.0] and sonicated to an average size of 0.5–1 kb. Chromatin extracts containing DNA fragments were immunoprecipitated using anti-hMOF antibodies (1:400, Abcam). Purified chromatin immunoprecipitation (ChIP) DNA and input DNA were analyzed by real-time PCR using SYBR Premix Ex Taq™ II, as previously described. Primer sequences are shown in supporting information Table S1.
Western blotting
Cells were washed with PBS and lysed in 2% SDS on ice for 30 min. Protein concentration was measured using a bicinchoninic acid (BCA) protein assay kit (Beyotime). A 20-μg amount of protein lysate was resolved by SDS polyacrylamide gel electrophoresis (SDS-PAGE), followed by transfer onto a polyvinylidinedifluoride (PVDF) membrane (Millipore). After blocking in PBS with 5% skim milk overnight at 4°C, the membrane was incubated with primary antibodies (anti-MOF, 1:400, Abcam; anti-H4K16ac, 1:5000, Abcam) at room temperature for 2 h, followed by fluorescence-conjugated secondary antibodies. Blots were washed and then scanned with the Odyssey Infrared Imaging System (LI-COR Biosciences).
Co-immunoprecipitation
Cells were collected and lysed in RIPA buffer (150 mM NaCl, 1% IGEPAL® CA-630, 0.5% sodium deoxycholate, 0.1% SDS, 50 mM Tris-HCl, pH 8.0) with protease inhibitor (TaKaRa). The protein extract was precleared with protein A–Sepharose beads (GE Healthcare) at 4°C for 2 h and incubated overnight at 4°C with anti-MOF antibody (Abcam) or rabbit immunoglobulin G (IgG) antibody (Millipore). Protein A–Sepharose beads were added for 2 h, and then washed with RIPA and TSE buffer (2 mM EDTA, 150 mM NaCl, 20 mM Tris-HCl, pH 8.0). A western blot analysis was performed to detect the proteins present in the isolated immunocomplexes.
Immunofluorescence
iPSCs were fixed in 4% PFA for 15 min and permeabilized with 0.1% Triton X-100 in PBS for 30 min. Then the cells were washed and blocked with 5% bovine serum albumin (BSA) for 30 min. Cells were incubated with the primary antibodies overnight at 4°C. The antibodies used were anti-OCT4 (1:100, Millipore), anti-SSEA4 (1:100, Millipore), anti-NANOG (1:100; Millipore), and anti-TRA-1-60 (1:100; Millipore). Then, the cells were washed and incubated with corresponding secondary antibodies for 1 h at room temperature. 4′,6-Diamidino-2-phenylindole (DAPI; 1:10,000, Beyotime) was used for nucleus staining. Images were detected using an IX5-RFACA Olympus fluorescence microscope.
Statistical analysis
Data were expressed as means±standard deviation (SD) and statistically analyzed by a Student t-test for paired observations. p values of less than 0.05 were considered statistically significant.
Results
Increased MOF expression during iPSCs generation
To investigate the role of MOF in iPSCs generation, we first monitored the level of mRNA encoding MOF in human hESCs and hFFs-derived iPSCs. The level of mRNA encoding MOF increased in both iPSCs and hESCs compared with hFFs (Fig. 1A). Moreover, western blotting experiments showed that the expression level of MOF protein was highly upregulated from hFFs to iPSCs (Fig. 1B). To explore the relationship between MOF expression and the acquisition of pluripotency during the reprogramming process, we infected hFFs with retroviruses encoding Oct4, Sox2, Klf4, and c-Myc (OSKM), and RNA was extracted in regular time intervals. The dynamic expression of MOF during reprogramming was analyzed by quantitative real-time polymerase chain reaction (qRT-PCR) and related to the expression of Nanog. MOF mRNA gradually increased, which was similar to the expression of Nanog during the period of iPSCs formation, and thus established iPSCs showed high MOF expression (Fig. 1C, D). The positive correlation between MOF and pluripotency led us to investigate whether MOF plays an active role in iPSCs generation.

Increased MOF expression during iPSCs generation. (
Ectopic overexpression of MOF improves the reprogramming efficiency
To further investigate whether ectopic MOF expression could promote iPSCs generation, hFFs were infected with retroviruses encoding OSKM together with MOF. Three weeks later, MOF-iPSCs displayed typical ESCs morphology and properly reprogrammed cells displayed the loss of GFP fluorescence as a result of retroviral transgene silencing (Fig. 2A). Immunofluorescence studies showed that they expressed the pluripotency markers OCT4, SSEA-4, NANOG, and TRA-1-60 (Fig. 2B). RT-PCR experiments revealed that MOF-iPSCs expressed ESCs markers, including endogenous Oct4, Sox2, and the other markers Nanog, Rex1, Esg1, and Gdf3 (Fig. 2C). Moreover, MOF-iPSCs were capable of forming EBs, differentiated into tissues originating from all three germ layers, and gave rise to teratomas (Fig. 2D, E). AP staining was performed. We counted the number of colonies with positive AP staining and typical ESCs-like morphology as the AP+ colonies (Fig. 2F). The number of AP+ colonies was markedly increased in the case of MOF overexpression. Interestingly, ectopic MOF expression caused three-fold higher frequency of AP+ colonies compared to the control OSKM group (Fig. 2G, H). In summary, overexpression of MOF could improve the four-factor induced iPSCs generation.

Ectopic overexpression of MOF enhances the efficiency of iPSCs generation. HFFs were transduced with the four reprogramming factors [Oct4, Sox2, Klf4, and c-Myc (OSKM)] plus MOF, or empty vector control. (
Overexpression of MOF cannot compensate for c-Myc in iPSCs generation
c-Myc is important for efficient iPSCs generation, but it could increase tumorigenicity in the chimeric mice (Nakagawa et al., 2008). Here, we examined whether MOF could compensate for c-Myc in iPSCs generation. To this end, iPSCs were generated from hFFs with the three reprogramming factors, Oct4, Sox2, and Klf4 (OSK), with or without MOF. The number of AP+ colonies at day 21 after transduction was reduced without c-Myc (Fig. 3A, B). The frequencies of AP+ colonies were only slightly increased with forced MOF compared to OSK. This indicates that overexpression of MOF could not substitute for c-Myc in efficient iPSCs generation.

MOF cannot replace c-Myc during iPSCs generation. hFFs were transduced with three reprogramming factors (OSK) with or without MOF. (
Ablation of endogenous MOF reduces reprogramming efficiency
We used shRNA-mediated knockdown of MOF to determine whether MOF is required for iPSCs generation. Retrovirus-mediated shRNA targeting of MOF (shMOF) caused effective downregulation of MOF expression in hFFs, as determined by qRT-PCR (data not shown). hFFs were infected with retroviruses containing OSKM together with or without MOF shRNA. At day 21, knockdown of MOF caused a strong reduction of AP+ colonies compared with the OSKM group (Fig. 4A, B). Meanwhile, this result was not due to an adverse effect of MOF depletion on the proliferative capacity of hFFs (Fig. 4C). To find whether MOF is required for the early/initiation phase or the later/expansion phase of reprogramming, we used a doxycycline-inducible pLKO-Tet-On lentiviral vector encoding MOF shRNA. hFFs were transfected with MOF shRNA. The level of hMOF mRNA was significantly decreased when 2 μg/mL doxycycline (Sigma) was added for 24 h (data not shown). OSKM-infected hFFs were treated with doxycycline at various time points (days −3, 0, 3, 6, and 9) to induce MOF shRNA expression. The most obvious reduction in iPSCs formation was observed when MOF was depleted during the initial stages of reprogramming (Fig. 4D). Taken together, these data suggest that MOF is required for efficient iPSCs generation.

Knockdown of MOF impairs iPSCs generation. (
MOF triggers establishment of euchromatic features at the promoter of pluripotency genes
Because Nanog, Sox2, and Oct4 form the core regulatory circuitry in ESCs (Boyer et al., 2005), we reasoned that MOF might be required for the reactivation of these three pluripotency genes and other markers of ESCs. To test this prediction, we examined the impacts of MOF depletion on the activation of ESCs pluripotency markers during reprogramming. qRT-PCR analysis on day 14 revealed that knockdown of MOF substantially reduced the expression of Nanog, Oct4, Sox2, Esg1, Cripto, and Rex1, which were highly activated in OSKM group (Fig. 5A). To identify direct target genes of MOF among core pluripotency genes, ChIP was carried out to examine the occupancy of MOF at these pluripotency genes in hESCs. The binding of MOF could be mainly detected at the promoter of Oct4 and Nanog, but not Sox2 (Fig. 5B). This result suggests that binding of MOF may promote Oct4 and Nanog transcription, making them acquire induced pluripotency during reprogramming. In support of this view, depletion of MOF led to decreased MOF occupancy at the Oct4 and Nanog promoters in hFFs transduced with retroviruses containing OSKM and shMOF compared with OSKM group at day 14 (Fig. 5C). Consequently, the levels of H4K16ac and H3K4me3 at the Oct4 and Nanog promoters were decreased in hFFs transduced with retroviruses containing OSKM and shMOF compared with the OSKM group (Fig. 5D)
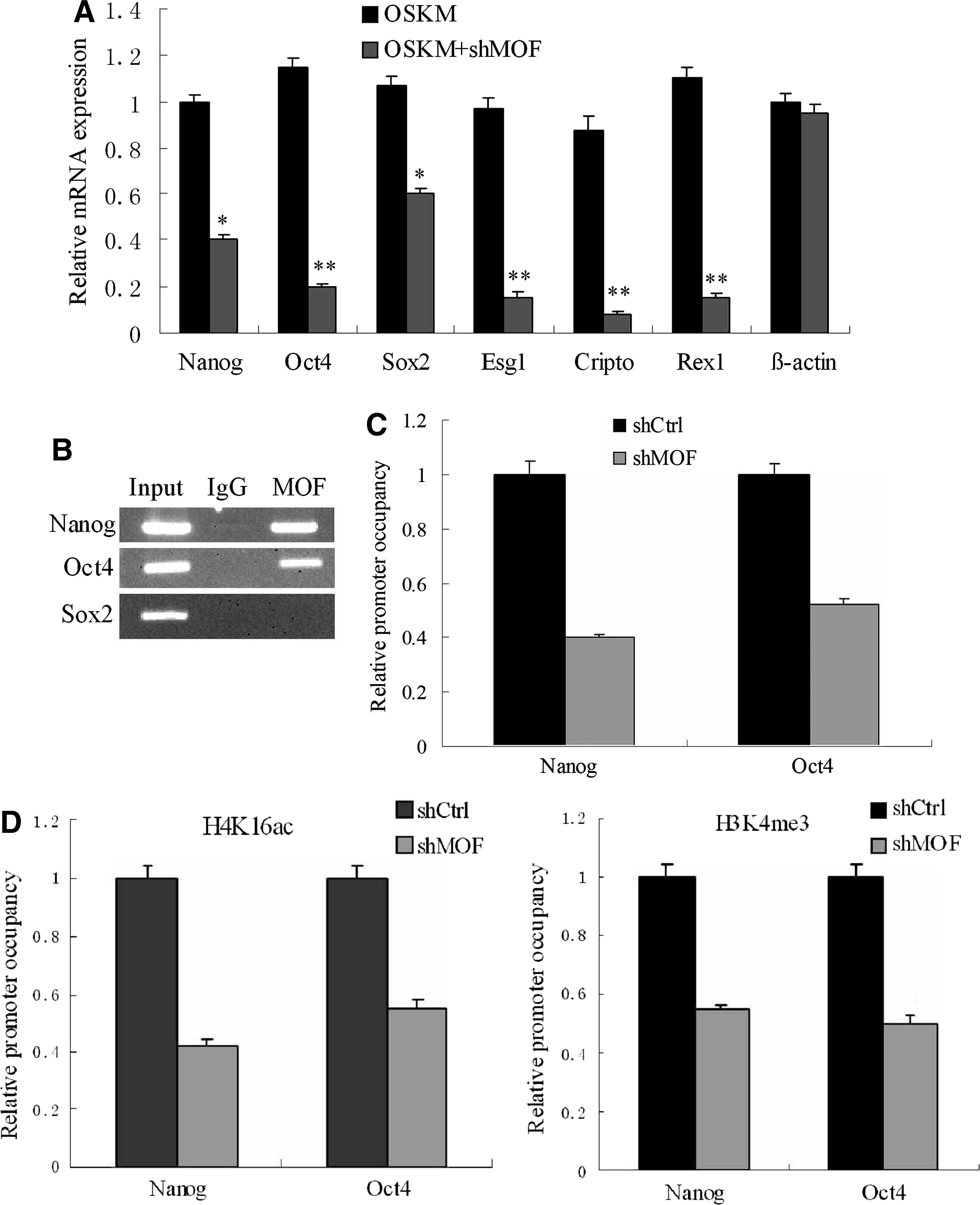
MOF triggers establishment of euchromatic features at the promoter of pluripotency genes. (
MOF acts partly through regulating H3K4 methyltransferase Wdr5 during iPSCs generation
Given the direct interaction between MOF and Wdr5 as well as the recent demonstration that Wdr5 partnered with Oct4 to reset the epigenome during reprogramming (Ang et al., 2011; Dou et al., 2005; Wysocka et al., 2005), we predicted that MOF might interact with Wdr5 to promote endogenous Oct4 expression during reprogramming. A co-immunoprecipitation (Co-IP) assay was performed in hESCs. We found that Oct4 and Wdr5 were detected in a MOF immunoprecipitated sample but not in an IgG control sample, confirming that MOF could interact with Oct4 and Wdr5 in hESCs (Fig. 6A). Next, ChIP was carried out to determine the binding of Wdr5 at the Oct4 promoter in hFFs and MOF-deficient hFFs. The ability of Wdr5 to bind to the Oct4 promoter was markedly reduced in hFFs transduced with retroviruses containing OSKM and shMOF compared with OSKM group at day 14, suggesting that MOF recruits Wdr5 to the Oct4 promoter (Fig. 6B). hFFs were co-transfected with OSKM, MOF shRNA, and with or without Wdr5. Efficiency of reprogramming was measured at day 21 by AP staining. As expected, knockdown of MOF decreased the reprogramming efficiency without Wdr5, and this was partially rescued by simultaneous overexpression of Wdr5 (Fig. 6C, D). Taken together, these results suggest that MOF may recruit Wdr5 to bind to the Oct4 promoter and reactive endogenous Oct4 expression during reprogramming.
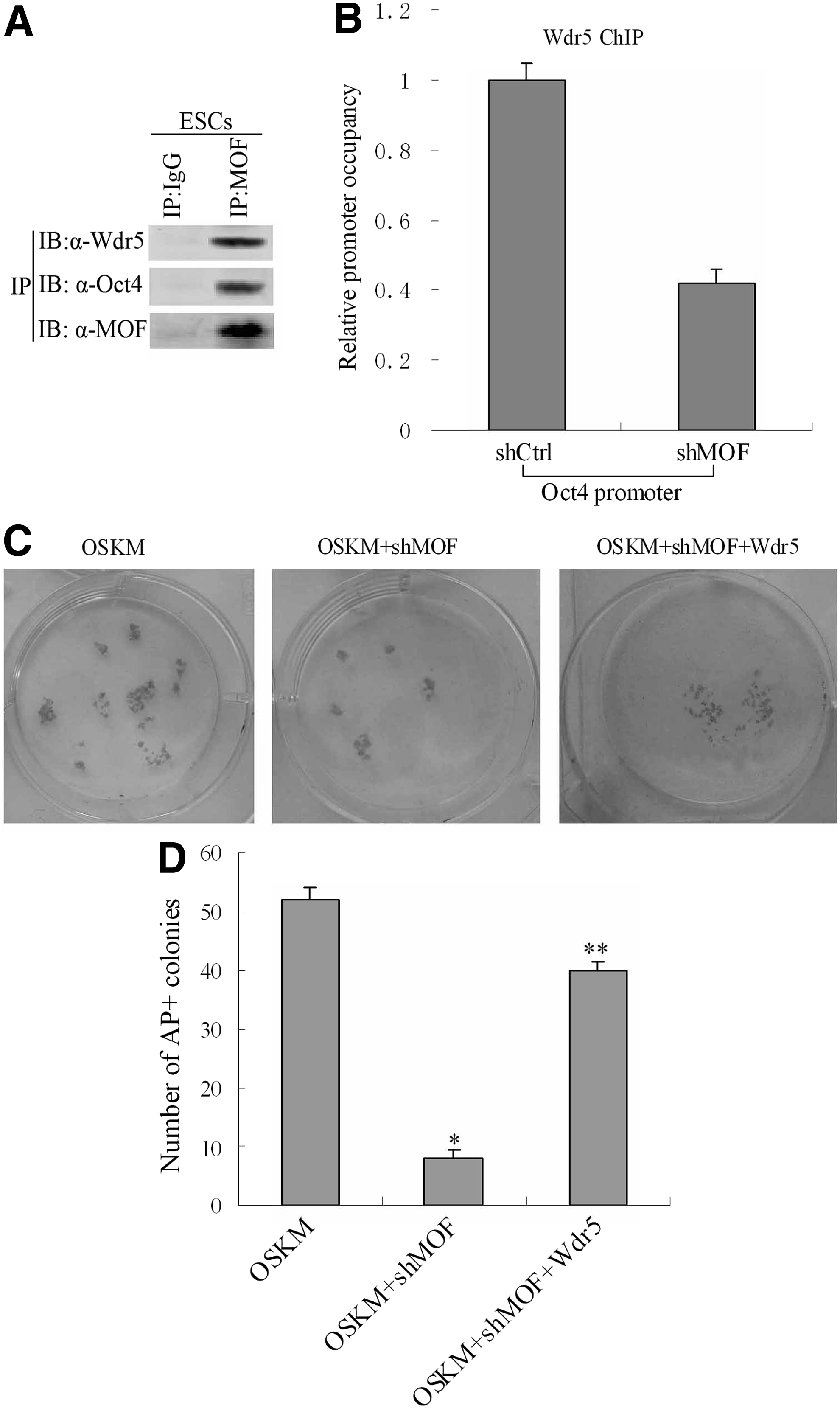
MOF acts partly through regulating H3K4 methyltransferase Wdr5 during iPSCs generation. (
Discussion
Generation of iPSCs by somatic cell reprogramming involves global epigenetic remodeling (Takahashi and Yamanaka, 2006). Some epigenetic factors and chemical modulators of epigenetic modifications are capable of affecting reprogramming efficiency. Consistent with this notion, many studies focused on examining the histone modification changes in the reprogramming process (Ang et al., 2011; Huangfu et al., 2008a, b; Liang et al., 2010; Mali et al., 2010; Singhal et al., 2010). However, the role of specific histone modifying enzymes in reprogramming remains to be determined. In this study, we examined whether MOF participates in the reprogramming process or plays a positive function in iPSCs generation. We show that MOF is efficiently induced during the reprogramming process, and the level of mRNA encoding MOF increases gradually along with the expression of the pluripotency gene Nanog. By performing a western blot analysis, we show that iPSCs contain high levels of MOF protein, which are similar to the levels in ESCs. Importantly, we show that depletion of MOF impairs iPSCs generation, and thus, ectopic expression of MOF can significantly increase the reprogramming efficiency. Meanwhile, MOF is required for the early/initiation phase of somatic cell reprogramming.
To further investigate the possible mechanisms by which MOF affects reprogramming efficiency, we examined the relationship between MOF and the H3K4 methyltransferase Wdr5. Fully reprogrammed iPSCs are associated with reactivation of endogenous Oct4 and several other ESCs marker genes (Plath and Lowry, 2011; Stadtfeld Hochedlinger, 2010). A recent study shows that the reprogramming factors OSKM could induce Wdr5 expression and promote formation of Wdr5–Oct4 complexes that bind to the Oct4 promoter, leading to reactivation of endogenous Oct4 expression. This study posited that Wdr5 partnered with Oct4 to reset the epigenome and promote reprogramming (Wysocka et al., 2005).
Another study shows that regulation of the ESC core transcription network by Wdr5 could be partially mediated by MOF, and binding of Wdr5 at selected pluripotent gene targets was MOF dependent (Li et al., 2012). This result suggests that MOF is an important upstream regulator of Wdr5 at ESCs pluripotent loci such as Oct4. However, it is unclear whether the effects of MOF on iPSCs generation were Wdr5 dependent. To link MOF directly to Oct4 expression, we first showed that MOF not only binds to the Oct4 promoter region but also specifically interacts with Wdr5 and Oct4, suggesting a critical requirement for MOF in reactivation of endogenous Oct4 expression. Deletion of MOF reduces the ability of H4K16ac, H3K4me3, and Wdr5 to bind to the Oct4 promoter, which is consistent with the results of a recent report showing that H3K4 methylation is closely associated with H4K16 acetylation (Wang et al., 2009).
The results further demonstrate that knockdown of MOF decreases the reprogramming efficiency without Wdr5, which is rescued by simultaneous overexpression of Wdr5, suggesting that MOF increases the reprogramming efficiency partially through regulating Wdr5 during iPSCs generation.
In summary, the work presented here demonstrates that MOF could promote iPSCs generation. MOF protein regulates Wdr5 during reprogramming and then leads to the expression of endogenous Oct4, thereby improving reprogramming efficiency. Further work is needed to elucidate the exact mechanisms by which MOF affects reprogramming. The study emphasizes the importance of epigenetic modification in cellular reprogramming toward pluripotency. In addition, it adds to our understanding of the underlying molecular mechanisms ongoing during iPSCs induction.
Footnotes
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.