
Abstract
Abstract
Bone defects are severe burdens in clinics, and thus cell therapy offers an alternative strategy exploiting the features of bone marrow stromal cells (BMSCs). Sheep are a suitable orthopedic preclinical model for similarities with humans. This study compares the influence of two phosphate sources combined with bone morphogenetic protein-2 (BMP-2) on the osteogenic potential of human and ovine BMSCs. β-Glycerophosphate (β-GlyP) and monosodium phosphate (NaH2PO4) were used as organic and inorganic phosphate sources. Osteogenic differentiation of the BMSCs was assessed by calcified matrix, alkaline phosphatase (ALP) activity, and gene expression analysis. A higher calcified matrix deposition was detected in BMSCs cultured with NaH2PO4. Although no significant differences were detected among media for human BMSCs, β-GlyP with or without BMP-2 determined a positive trend in ALP levels compared to NaH2PO4. In contrast, NaH2PO4 had a positive effect on ALP levels in ovine BMSCs. β-GlyP better supported the expression of COL1A1 in human BMSCs, whereas all media enhanced RUNX2 and SPARC expression. Ovine BMSCs responded poorly to any media for RUNX2, COL1A1, and SPARC expression. NaH2PO4 improved calcified matrix deposition without upregulating the transcriptional expression of osteogenic markers. A further optimization of differentiation protocols needs to be performed to translate the procedures from preclinical to clinical models.
Introduction
B
Many studies have reported the in vitro osteogenic differentiation of BMSCs derived from animals and humans, but differently from human BMSCs (hBMSCs), ovine BMSCs (ovBMSCs) are poorly characterized. Therefore, the in vitro behavior and osteogenic potential of ovBMSCs need to be examined deeply for the proper use of this cell source in bone cell therapy and tissue engineering.
The in vitro differentiation of BMSCs largely depends on culture conditions; in fact, bioactive agents like ascorbic acid 2-phosphate, dexamethasone, and β-glycerophosphate (β-GlyP) are able to induce osteogenic differentiation (Langenbach and Handschel, 2013). Moreover, the osteogenic differentiation of hBMSCs can be improved by bone morphogenetic protein-2 (BMP-2) (Suzuki et al., 2006; Wang et al., 2010; Wang et al., 2012). A recent study pointed out that ovBMSCs are able to produce calcified matrix only when cultured in the presence of an inorganic source of monosodium phosphate (NaH2PO4), assuming that the lack of alkaline phosphatase (ALP) in ovBMSCs impedes the release of phosphate ions from β-GlyP (Kalaszczynska et al., 2013). On the basis of these findings and because of the lack of extensive knowledge about ovBMSCs, this study attempts to identify an osteogenic medium composition able to properly differentiate hBMSCs and ovBMSCs, as different species-derived cells, by using an adequate phosphate ion source and evaluating the efficacy of BMP-2 to enhance osteogenic differentiation. We hypothesized that different phosphate ions combined or not with BMP-2 could improve the in vitro osteogenic differentiation of both hBMSCs and ovBMSCs. Supporting this hypothesis is essential in bone regenerative approaches, considering that humans are the final user of this technology and that the sheep is the elective animal model in orthopedics to bridge in vitro studies toward clinical trials.
Materials and Methods
BMSCs isolation and culture
Bone marrow aspirates were obtained from the iliac crest of six female skeletally mature sheep (mean age 5.1±2 years old) (n=6) under general anesthesia in compliance with national (Law 116/92, Italian Ministry of Health) and international laws and policies (EEC Council Directive 86/609, OJ L 358. 1, December 12, 1987; Standards for the Care and Use of Laboratory Animals–UCLA, US National Research Council, Statement of Compliance A5023-01, November 6, 1998). Human bone marrow samples were collected by written consent according to the Institutional guidelines (Galeazzi Orthopaedic Institute Scientific Committee) from the femoral canal of seven female osteoarthritic patients (mean age 55.7±13.3 years old) (n=7) who underwent hip prosthetic implantation.
Bone marrow samples were washed in phosphate-buffered saline (PBS) and centrifuged. The pellet containing ovine or human mononuclear cells was resuspended in complete medium (CM) composed of Dulbecco's Modified Eagle Medium High Glucose (DMEM-HG; Gibco), 10% fetal bovine serum (FBS; Hyclone), 100 U/mL penicillin–streptomycin, 2 mM
Osteogenic differentiation
Both hBMSCs and ovBMSCs were plated in triplicate at a density of 103 cells/cm2 and cultured in four different osteogenic media. All of the analyses were performed after 14 days of osteogenic differentiation.
The osteogenic medium was commonly composed of CM supplemented with 50 μg/mL ascorbic acid-2P, 0.1 μM dexamethasone (all from Sigma-Aldrich), and differently added with the following inducers of mineralization: (1) 3 mM NaH2PO4 (Sigma-Aldrich), (2) 10 mM β-GlyP (Sigma-Aldrich), (3) 3 mM NaH2PO4+10 ng/mL recombinant human BMP-2 (rhBMP-2; Peprotech), and (4) 10 mM β-GlyP+10 ng/mL rhBMP-2. Undifferentiated BMSCs cultured in CM were used as negative control (Ctrl).
The cell proliferation rate of hBMSCs and ovBMSCs based on DNA content was determined for each medium condition by means of a CyQUANT® Cell Proliferation Assay Kit (Life Technologies), according to the manufacturer's instruction.
Evaluation of calcified matrix deposition
For Alizarin Red-S (AR-S) staining, cells were fixed with ice-cold 70% ethanol for 30 min and stained with 40 mM AR-S (pH 4.1, Sigma-Aldrich) for 15 min. Then, samples were unstained with 10% cetylpyridinium chloride monohydrate (CPC; Sigma-Aldrich) in 0.1 M phosphate buffer (pH 7.0) for 15 min to quantify the staining. Absorbance was read at 570 nm (Lopa et al., 2014). von Kossa staining was performed to localize calcium as mineralized nodules. Cells were fixed in 10% formalin, stained with 5% silver nitrate solution (Sigma-Aldrich) under ultraviolet (UV) light for 1 h, and then counterstained with 0.1% Safranin (Bio-Optica) (Lovati et al., 2011).
ALP activity assay
ALP activity was quantified by enzymatic assay (Lopa et al., 2014). Cells were lysed in 0.1% Triton X-100 (Sigma-Aldrich). ALP was quantified by incubating cellular lysates at 37°C with 1 mM p-nitrophenylphosphate (Sigma-Aldrich) in alkaline buffer (100 mM diethanolamine and 0.5 mM MgCl2, pH 10.5). The enzymatic reaction was blocked with 1 N NaOH, and absorbance was read at 405 nm (Perkin Elmer Victor X3 microplate reader). ALP activity was normalized on total protein content, determined by a BCA Protein Assay Kit (Pierce Biotechnology), and expressed as ALP units/mg of proteins, as described elsewhere (Lopa et al., 2014; Rawadi et al., 2003).
Gene expression analysis
Gene expression was evaluated by real-time PCR (StepOne Plus, Life Technologies). Total RNA was purified using a PureLink RNA Mini Kit (Life Technologies). RNA was quantified and reverse-transcribed to cDNA using an iScript cDNA Synthesis Kit (Bio-Rad Laboratories). A total of 20 ng of cDNA was incubated with a PCR mixture including TaqMan® Universal PCR Master Mix and TaqMan® Assays-on-Demand™ Gene Expression Probes (Life Technologies). The following genes were analyzed: Glyceraldehyde 3-phosphate dehydrogenase (human GAPDH, HS99999905_m1; sheep GAPDH, Fw 5′-AGGGACTTATGACCACTGT-3′, Rv 5′-CCAGTAGAAGCAGGGATGATGT-3′), runt-related transcription factor 2 (RUNX2 HS00231692_m1), type I collagen alpha I (COL1A1, HS01076777_m1), and osteonectin (SPARC, HS00234160_m1). The expression of the analyzed markers was normalized on the expression of the housekeeping gene GAPDH.
Statistical analysis
All data in the graphs are reported as fold increase of treated cells with respect to undifferentiated BMSCs set as 1 (Ctrl). Statistical analyses were performed using commercially available software (GraphPad Prism 5). Normal distribution of values was assessed by the Kolmogorov–Smirnov normality test. Group comparisons were analyzed with one-way analysis of variance (ANOVA). When significant differences were detected, post hoc comparisons of means were performed using Dunnett's procedure. Data are expressed as mean±standard error of the mean (SEM). Values of p<0.05 were considered statistically significant.
Results
Characterization of BMSCs clonogenic ability, morphology, and growth capability
The mean CFU-F efficiency was 9.77%±5.52% for hBMSCs and 43.02%±12% for ovBMSCs, showing an extremely significant difference (p<0.0001). Under phase-contrast microscopy, both hBMSCs and ovBMSCs exhibited the typical fibroblast-like morphology characterized by a spindle shape. During the expansion phase, the PDT was significantly higher in hBMSCs than in ovBMSCs with values of 85.5±50.9 and 36.5±3.2 h, respectively (p<0.05), pointing out a higher proliferation rate in ovBMSCs.
After 14 days of culture in control medium or osteogenic medium, no significant differences were detected in terms of cell proliferation between undifferentiated cells compared to cells treated with the four different osteogenic formulations, as depicted in Figure 1A.
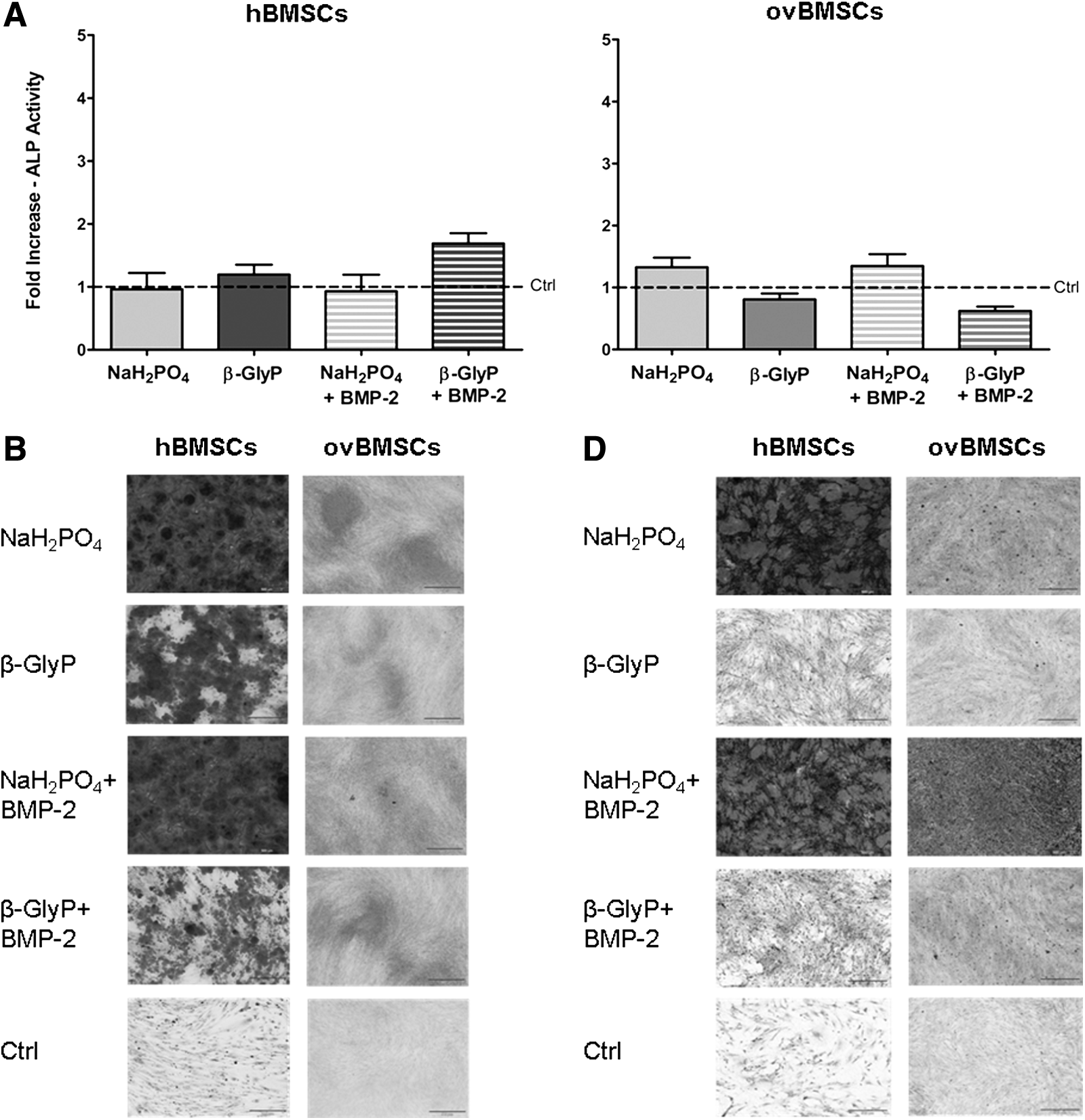
DNA quantification, qualitative and quantitative AR-S analysis, von Kossa staining, and specific ALP activity. (
Calcified matrix deposition
Overall, hBMSCs and ovBMSCs showed an increased amount of calcified matrix deposition compared to the undifferentiated cells (Fig. 1). In particular, osteogenic differentiated hBMSCs produced a greater amount of calcified matrix compared to ovBMSCs. Micrographs in Figure 1B depict a higher matrix deposition in hBMSCs when cultured in NaH2PO4 and NaH2PO4+BMP-2 compared to media containing β-GlyP or β-GlyP+BMP-2. Even though a lower amount of calcified deposits overall was observed in ovBMSCs, an initial calcified matrix deposition was present in cells treated with NaH2PO4 and NaH2PO4+BMP-2, as also observed in the experiment performed with hBMSCs.
AR-S quantification reported in Figure 1C shows an increased deposition both in differentiated hBMSCs and ovBMSCs compared to the undifferentiated controls. In particular, in hBMSCs, NaH2PO4 induced a significant increase of calcified matrix deposition compared to β-GlyP and β-GlyP+BMP-2. Similarly, hBMSCs treated with NaH2PO4+BMP-2 deposited significantly higher levels of matrix compared to β-GlyP and β-GlyP+BMP-2 (p<0.05). The addition of BMP-2 to NaH2PO4 promoted a significantly greater deposition of calcified matrix in hBMSCs compared to the other media, although not significantly compared to the medium containing NaH2PO4 alone. On the other hand, the addition of BMP-2 was not able to enhance the calcified matrix deposition in hBMSCs cultured with β-GlyP. Accordingly with the qualitative assessment of matrix deposition, the AR-S quantitative analysis showed a lower fold increase in ovBMSCs compared to hBMSCs. Similar to hBMSCs, ovBMSCs displayed higher levels of calcified matrix when treated with NaH2PO4 compared to β-GlyP or β-GlyP+BMP-2 (p<0.05). In ovBMSCs, the addition of BMP-2 to both NaH2PO4 and β-GlyP had a poor influence and did not increase the amount of calcified matrix. von Kossa staining revealed an increased presence of mineralized deposits in differentiated hBMSCs and ovBMSCs compared to the undifferentiated cells in which spots were barely present (Fig. 1D). Confirming the previous results, the medium containing NaH2PO4 and NaH2PO4+BMP-2 better enhanced calcified matrix deposition compared to β-GlyP and β-GlyP+BMP-2 media for both cell sources.
Specific ALP activity
Specific ALP activity showed significantly different mean values for hBMSCs (from 0.02 to 0.06 mU/μg) and for ovBMSCs (from 0.00006 to 0.0002 mU/μg) (p<0.0001) in all the tested media. Furthermore, ovBMSCs displayed a lower response to osteogenic induction respect to hBMSCs, as demonstrated by an inferior increase in ALP activity compared to undifferentiated cells.
Although no significant differences were detected among various formulations for hBMSCs, the medium containing β-GlyP with or without BMP-2 seemed to determine a higher fold increase compared to the media containing NaH2PO4 supplemented or not with BMP-2 (Fig. 1E). Overall, the medium containing β-GlyP+BMP-2 induced a greater increase of ALP activity in hBMSCs with respect to the other medium formulations. ovBMSCs showed a higher response when treated with NaH2PO4 compared to the other media, although the difference was significant only for medium containing β-GlyP+BMP-2 (p<0.01).
Gene expression analysis
The gene expression of all the tested osteogenic markers was increased in differentiated hBMSCs compared to undifferentiated cells, whereas ovBMSCs poorly responded to the osteogenic stimuli (Fig. 2). Indeed, hBMSCs responded to any osteogenic formulations expressing three-fold higher levels of RUNX2 compared to controls, whereas in ovBMSCs RUNX2 expression was similar to controls.
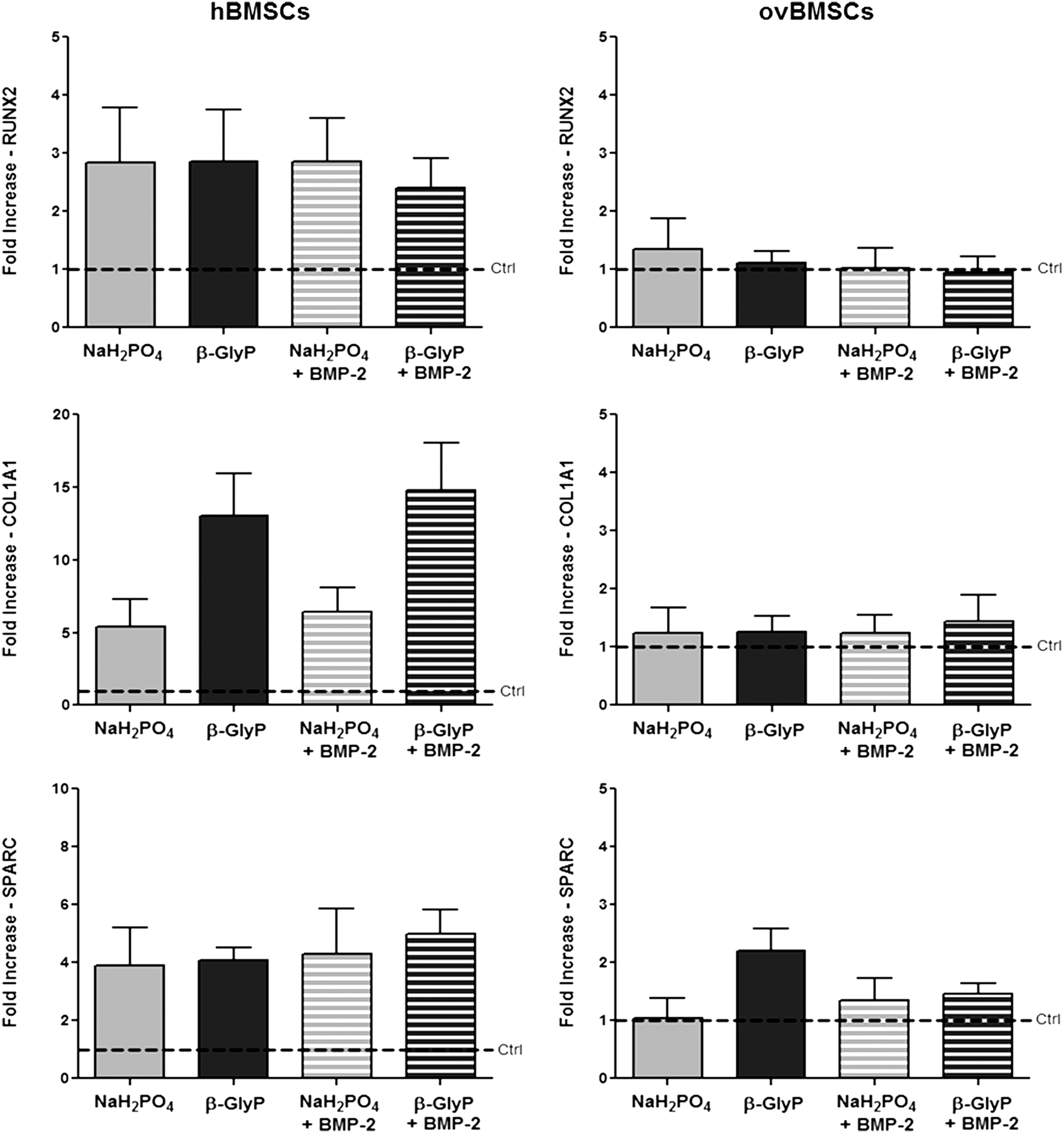
Osteogenic gene expression. RUNX2, COL1A1, and SPARC gene expression in hBMSCs and ovBMSCs cultured in four different osteogenic media. Data are reported as fold increase with respect to the undifferentiated BMSCs (dotted line) (mean±SEM).
Increases in COL1A1 expression in hBMSCs ranged from five- to 15-fold compared to controls and, in particular, media containing β-GlyP and β-GlyP+BMP-2 induced the highest levels of COL1A1 expression, although differences were not significant respect to media containing NaH2PO4. As previously observed for RUNX2, ovBMSCs did not show any increase respect to controls. Upregulation of SPARC expression was detected in hBMSCs exposed to any medium with about four-fold increase respect to controls. Once again, ovBMSCs did not show a relevant increase in SPARC levels except for cells treated with the medium containing β-GlyP.
Discussion
Because hBMSCs are a suitable cell source in bone tissue engineering and regenerative medicine approaches (Murray et al., 2014) and ovBMSCs are involved in preclinical investigation, in the present study, we hypothesized that the combination of an adequate phosphate ion source supplemented or not with BMP-2 could represent a valid strategy to identify an osteogenic differentiation protocol useful both for hBMSCs and ovBMSCs. This work aimed to compare the osteogenic capability of BMSCs derived from two different species as final user and preclinical model in orthopedics, extending the recent findings published by Kalaszczynska and colleagues (Kalaszczynska et al., 2013).
The ovBMSCs showed a typical spindle-shaped, fibroblast-like morphology very similar to hBMSCs, although a smaller diameter was detected in ovBMSCs compared to hBMSCs, as confirmed by others (Rentsch et al., 2010). During in vitro culture, hBMSCs displayed precocious signs of senescence that can be explained by cell origin. Indeed, in our study, we used cells derived from elderly osteoarthritic patients, whereas ovBMSCs were harvested from adult and healthy sheep. According to these cell source characteristics, the proliferation of ovBMSCs was faster compared to hBMSCs, showing a significantly lower PDT. Supporting our results, similar PDT values for ovBMSCs and hBMSCs have been reported elsewhere (Rentsch et al., 2010). Higher PDT values observed in hBMSCs are associated with an inferior clonogenic ability that is potentially correlated to the advanced age of human donors, as recently demonstrated by others (Li et al., 2014).
In the present study, we applied various osteogenic differentiation protocols employing different sources of phosphate in combination with a well-recognized osteoinductive factor, BMP-2. The concentration of phosphate sources was chosen on the basis of recently published studies (Hoemann et al., 2009; Kalaszczynska et al., 2013).
AR-S analysis demonstrated the ability of BMSCs to produce calcified matrix. We observed that NaH2PO4 constantly represented the favorite source of phosphate ions used by cells to deposit calcium phosphate both in hBMSCs and in ovBMSCs. Matrix deposition is known to be affected by the duration of the osteogenic differentiation process (Hoemann et al., 2009; Lopa et al., 2014), as well as by donor-specific features (Janicki et al., 2011; Kim et al., 2012). In fact, in our study, we detected a high interdonor variability in terms of calcified matrix deposition in both cell sources. The strong dependence of calcified matrix deposition on culture time can explain the mild AR-S staining observed in ovBMSCs, whereby the osteogenic differentiation had to be interrupted early (10 days) to prevent monolayer detachment, according to the higher proliferation rates detected in ovBMSCs. Indeed, it is well-recognized that the main stages commonly occur in the osteogenic differentiation of BMSCs, whereby an initial phase of proliferation is followed by the expression of ALP and collage type I supporting the mineral deposition, and by the detection of late osteogenic genes, such as osteocalcin and osteopontin (Birmingham et al., 2012; Hoemann et al., 2009).
BMP-2 promoted a significant increase in calcified matrix deposition in hBMSCs enhancing the free phosphate ion transport, as also reported in other studies on human cells (Suzuki et al., 2006; Wang et al., 2010; Wang et al., 2012). On the other hand, the addition of BMP-2 did not ameliorate the matrix deposition in ovBMSCs, as also demonstrated for in vivo bone healing in sheep (Egermann et al., 2006). This result is also supported by Blum et al. (2004), who found a positive effect of BMP-2 on the osteogenic differentiation of ovBMSCs when transfected into the cells but not when added to the culture medium. The von Kossa staining provided a further confirmation of these results. In fact, calcium phosphate deposition was abundant in both hBMSCs and ovBMSCs differentiated with NaH2PO4 compared to media enriched with β-GlyP, and no positive effect was observed for BMP-2 on ovBMSCs.
Our results are in contrast to those presented by Kalaszczynska et al. (2013), in which NaH2PO4 did not enhance matrix deposition in hBMSCs. Conversely, our findings are supported by other studies on hBMSCs osteogenic differentiation in which NaH2PO4 was considered a better phosphate ion source with respect to β-GlyP (Boskey and Roy, 2008; Schäck et al., 2013). In particular, Schäck et al. (2013) have demonstrated that the hBMSCs interdonor variability of ALP activity influences the release of β-GlyP during osteogenic differentiation, whereas NaH2PO4 provides cells with a constant amount of free phosphate ions.
ALP activity is commonly used as an osteogenic differentiation marker and is an enzyme essential for the release of phosphate ions (Bellows et al., 1991). In our study, ALP activity detected in hBMSCs was significantly higher compared to that of ovBMSCs under all the conditions, accordingly with recently published data by Kalaszczynska et al. (2013). The normalization of ALP levels found in undifferentiated cells revealed that, despite being low, ALP levels were increased upon osteogenic stimulation, even in ovBMSCs. In particular, a higher fold increase of ALP activity was found in ovBMSCs treated with NaH2PO4, compared to the other treatments, confirming the AR-S results. hBMSCs showed a higher specific ALP activity when cultured in media containing β-GlyP instead of NaH2PO4. This result is in agreement with the physiological role of ALP, which is known to be strictly implicated in the beginning of the mineralization process by means of the phosphate ion release from β-GlyP (Bellows et al., 1991). Once again, BMP-2 did not increase ALP levels in ovBMSCs, whereas we observed a positive trend in ALP levels in hBMSCs when both β-GlyP and NaH2PO4 media were supplemented with BMP-2.
The osteogenic markers analyzed were upregulated in differentiated hBMSCs compared to the controls, whereas ovBMSCs showed a lower response to differentiation compared to hBMSCs. Overall, we can assert that the addition of β-GlyP better supported the expression of COL1A1 in hBMSCs, whereas all of the tested media indiscriminately enhanced the expression of RUNX2 and SPARC. Because RUNX2 is a transcriptional factor involved in the early phase of osteogenic differentiation (Dalle Carbonare et al., 2012), we reasonably supposed that possible variations in RUNX2 expression, depending on the different media, were not appreciable at the time we performed the gene expression analysis.
Our study demonstrated that osteogenic media supplemented with NaH2PO4 better supported calcified matrix deposition with β-GlyP in both hBMSCs and ovBMSCs. This result is in contrast with recent data from Kalaszczynska et al. (2013), who demonstrated that NaH2PO4 is a suitable phosphate source for ovBMSCs but not for hBMSCs. Moreover, our deeper insight into gene expression of osteogenic markers revealed a poor response in ovBMSCs toward osteogenic differentiation induced by NaH2PO4, which, to the best of our knowledge, has never been reported before. The choice of a single time point for all the analyses may have been detrimental to the detection of time-related variations in osteogenic markers and represents a major limitation of this study. Further investigations will be needed to assess the osteogenic response at different time points and to evaluate the combination of the two ion phosphate sources in ovBMSCs, to define a suitable medium formulation to improve their osteogenesis, and to implement their use in translational preclinical models.
Footnotes
Acknowledgments
This study was supported by the Italian Ministry of Health (grant no. RF-IOG-2010-2322039).
Authors Disclosure Statement
The authors declare that no conflicting financial interests exist.