
Abstract
Abstract
The thymus is mainly composed of two types of epithelial cells, medullary thymic epithelial cells and cortex thymic epithelial cells (mTECs and cTECs). The tissue structure and mechanism for T cell development are complicated, with generation of the thymus regulated by complex molecular and cellular interactions of the thymic microenvironment during embryogenesis. Since the development of organ regeneration techniques became available, complete in vitro regeneration of the thymus has been attempted. Steric induction systems are thought to be optimal for tissue regeneration, but three-dimensional (3-D) induction of TECs from induced pluripotent stem cells (iPSCs) has not yet been reported. Here, we demonstrate the induction of functional TECs from iPSCs by a 3-D spheroid culture system with recruitment of robust numbers of T cells into the peripheral blood. Purified iPSC-derived TECs showed a sufficient expression level of FoxN1 comparable to TECs, and phenotypic analysis revealed that iPSC-derived TECs were expressing K5. Moreover, transplants of cell aggregations into recipient mice were not rejected and there was normal support of T cell development. Functional analysis revealed that T cells showed immune tolerance to both donor and recipient MHCs and could reject an allogeneic third party's skin graft without tumorigenesis. Taken together, these findings raised the possibility of using iPSC-derived TECs induced by 3-D spheroid culture in future regenerative therapy for patients with immunodeficiency.
Introduction
T
Previously, we reported the induction of phenotypically TEC-like cells from pluripotent stem cells by two-dimensional (2-D) culture. However, the functionality of these cells has not yet been proven (Inami et al., 2011). Because we thought that spatiotemporal cell–cell interaction was essential to inducing both mTECs and cTECs, we thought that three-dimensional (3-D) spheroid culture system might induce functional TECs. 3-D culture systems are thought to be a more optimal condition for cell differentiation and organ regeneration (Bokhari et al., 2007; Kyba et al., 2002; Saito et al., 2011; Suga et al., 2011), but induction of functional TECs by 3-D culture for organ regeneration has not been reported. In this study, we investigated the induction of TECs from iPSCs by a 3-D spheroid culture system.
Materials and Methods
Mice
Six- to 8-week-old BALB/c, KSN nude, and C57BL/6 mice were purchased from Japan SLC (Shizuoka, Japan). These mice were maintained in the Animal Research Facility at the Nagoya University Graduate School of Medicine under specific pathogen-free conditions and used for these experiments according to institutional guidelines. This study was carried out in strict accordance with the recommendations of the Regulations on Animal Experimentation at Nagoya University. The Animal Care and Use Committee of Nagoya University Graduate School of Medicine approved the protocol.
Cell culture and cell differentiation of iPSCs
Generation of mouse embryonic fibroblast-derived iPSCs (C57BL/6) was as previously described by Cheng et al. (2011). For TEC induction, the culture conditions used were according to our previous method (Inami et al., 2011) and were modified to 3-D spheroid conditions. Ten thousand iPSCs were seeded onto 96-well low-cell-adhesion plates (Lipidure Coat, NOF Corp., Japan) in SF-O3 culture medium (Sanko Junyaku, Tokyo, Japan) supplemented with 0.2% bovine serum albumin (BSA), 5% fetal bovine serum (FBS), and 0.1 mM 2-mercaptoethanol with the addition of the following materials. From days 1 to 4, these cultures were supplemented with 10 ng/mL Activin A (R&D systems, Minneapolis, MN, USA) and 5 mM LiCl (Sigma-Aldrich). At day 4, the culture supernatant was replaced with fibroblast growth factor 8 (FGF8; R&D Systems) and LiCl. At day 8, the culture supernatant was replaced with FGF7, FGF10 (R&D Systems), Bone Morphogeneic Protein (BMP4) (R&D Systems), and LiCl. At day 12, the medium was changed to the day 8 conditions supplemented with 20 ng/mL receptor activator of nuclear factor kappa-B ligand (RANKL), and cells were cultured in the medium for 4 days.
Flow cytometric analyses
Cultured cells were trypsinized by 0.05% trypsin–EDTA (Gibco) and harvested. The single-cell suspensions were incubated with BV605-conjugated anti-CD326 (EpCAM1; BD Biosciences, San Jose, CA, USA) and purified anti-SSEA1 (Millipore) antibodies. Cells were then incubated with phycoerythrin (PE)-conjugated anti-mouse immunoglobulin M (IgM) antibody. For double negative (DN) cells and TEC purification, thymus tissue was minced and incubated with collagenase for 30 min at 37°C. Cells were stained with fluorescein isothiocyanate (FITC)-conjugated anti-CD4, PE-conjugated anti-CD8, allophycocyanin (APC)-conjugated anti-CD45, and BV605-conjugated anti-CD326 (EpCAM1). To analyze recipient-mouse peripheral blood cells, the erythrocytes were lysed, and the remaining cells were stained with FITC-conjugated anti-B220, PE-conjugated anti-CD4, and APC-conjugated anti-CD8 antibodies. Biotinylated antibodies were detected with streptavidin–APC–Cy7. Analysis and cell sorting were performed on a FACSCantoII or FACSAriaII (Becton Dickinson, Japan) and analyzed using FlowJo software (Tree Star, Ashland, OR, USA).
Subrenal capsule transplantation
Purified EpCAM1+SSEA1− cells derived from iPSCs (100,000) were cultured and reaggregated with or without CD4−CD8−CD45+ DN thymocytes (100,000) from BALB/c mice on Lipidure plates for 24 h. The reaggregates were grafted under the subrenal capsule of nude mice. Aggregations of DN cells from neonatal thymus of BALB/c or iPSC-derived TECs were used as a control. The peripheral blood of recipient mice was assessed after 6 weeks of transplantation.
Skin grafting
Skin graft analysis was performed as previously described (Oikawa et al., 2007). In brief, tail skins of C57BL/6 and C3H were cut into small pieces and placed on the shaved back of recipients. The grafts were protected by wrapping with a bandage, and rejection was assessed every 2 days after day 6 of the skin graft.
Hematoxylin & Eosin staining and immunostaining
Histological analysis was performed as previously described (Inami et al., 2011; Ito et al., 2013). In brief, the sections were fixed with 4% paraformaldehyde and stained with Hematoxylin & Eosin (H&E). For immunohistochemistry, the sections were fixed with 4% paraformaldehyde. Blocking was done in PBS containing 2% BSA for 1 h, followed by incubation with the primary antibody rabbit anti-cytokeratin5. Alexa Fluor 488–conjugated goat anti-rabbit IgG was used as a secondary antibody. Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI).
RT-PCR
Total RNA was isolated using TRIzol reagent (Invitrogen). First-strand cDNA was synthesized using the Superscript First-Strand Synthesis System (Invitrogen) for RT-PCR and oligo d(T)12–18 primers. The cDNA was diluted with DNase-free water to a concentration of 10 ng/mL. RT-PCR was performed using the Ex-Taq PCR kit (TaKaRa) according to the manufacturer's instructions.
Results
Induction of TEC-like cells from iPSCs by 3-D spheroid culture system
Since we previously reported the induction of TEC-like cells from iPSCs in 2-D culture (Inami et al., 2011), we attempted to induce TECs from iPSCs by using low-attachment 96-well plates for 3-D culture (Fig. 1A) (Suga et al., 2011; Wataya et al., 2008). In this study, we used C57BL/6-derived iPSCs (H-2b) (Cheng et al., 2011). When the 10,000 iPSCs were seeded onto low-attachment U-bottomed 96-well plates, cell aggregates were formed after 4 days of induction and became large spheroids at day 8. After 16 days of induction, a 3-D capsule-like structure was formed (3-D spheroid) (Fig. 1B). When we examined TEC marker genes in the 3-D spheroid, we could detect the expression of TEC marker genes, such as AIRE, DLL1, K5, or K8 at several time points. Furthermore, the expression of FoxN1, a key marker gene for TEC differentiation, was distinctly detected. However, we could still detect the expression of Nanog, an indicator of pluripotency, which might be the result of contamination with undifferentiated iPSC cells (Fig. 2A).
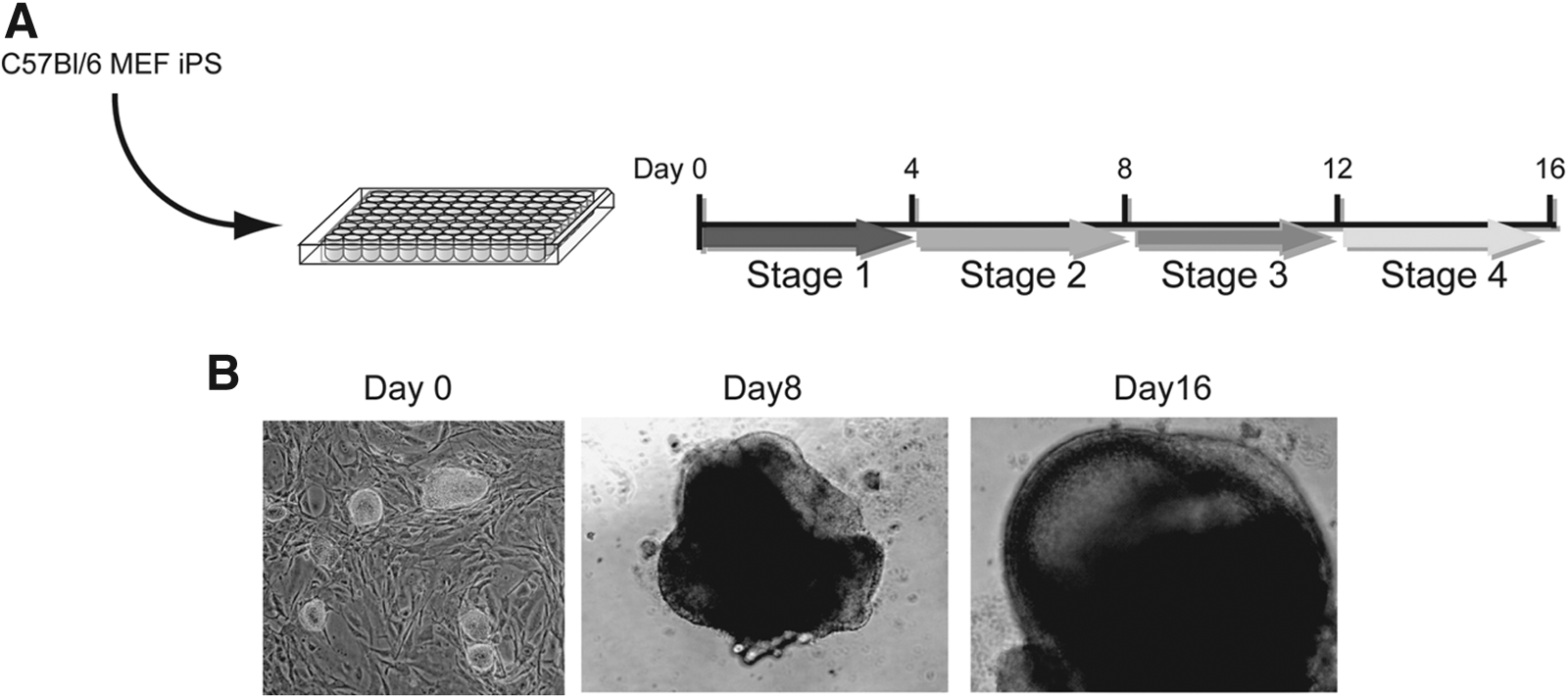
Induction of TECs from iPSCs by 3-D culture system. (

Induction of TECs from iPSCs by a 3-D culture system. (
To examine the contamination with undifferentiated iPSCs in spheroids, we performed flow cytometry analysis. Although a previous study showed that EpCAM1 could be used as a TEC marker (Rossi et al., 2006), we found that EpCAM1 was also expressed in pluripotent stem cells (data not shown). Therefore, we added the marker SSEA1+ to EpCAM1+ spheroids to exclude iPSC contamination. When the frequency of EpCAM1+SSEA1− cells was assessed on days 7, 10, and 13 of TEC induction by flow cytometry with or without FBS, we found that the day-10 spheroids with 5% FBS yielded the largest number of EpCAM1+SSEA1− cells (∼17% of culture cells) (Fig. 2B). Furthermore, 3-D spheroid cultures with 5% FBS highly expressed K5 (Fig. 2C).
Purification of TEC-like cells from the 3-D spheroid induction culture
We then decided to enrich TEC-like cells from spheroids. We purified EpCAM1+SSEA1− cells on day 10 of induction with 5% FBS. As we expected, EpCAM1+SSEA1− cells expressed FoxN1 preferentially at a comparable level to wild-type thymus cells, whereas EpCAM1+SSEA1− cells did not express Nanog. We could also detect the expression of DLL4, a key molecule for T cell development that expresses on TECs (Fig. 2D). These results showed that iPSCs were differentiated into TECs using the 3-D spheroid culture system. As we have previously demonstrated in a 2-D culture system, we found that stepwise differentiation of TECs via definitive endoderm and anterior foregut endoderm was also necessary in 3-D spheroid culture by stimulating pluripotent stem cells consecutively with Activin A, FGF8, BMP4, FGF7, and FGF10 (Figs. 1 and 2).
Transplantation of iPSC-derived TECs enhances T cell development in nude mice
To analyze the functionality of iPSC-derived TECs in vivo, 100,000 purified EpCAM1+SSEA1− cells were reaggregated for 24 h in low-attachment 96-well plates with equivalent number of CD4−CD8−CD45+ DN thymocytes derived from the neonatal thymus of BALB/c mice and transplanted into subrenal capsule of nude mice (Fig. 3) (Anderson et al., 1993). Six weeks after transplantation, the peripheral blood of the recipient mice was analyzed to assess whether iPSC-derived TECs could contribute to T cell development. CD8+ T cells were detected in the peripheral blood of recipient mice, which were transplanted with iPSC-derived TECs mixed with DNs, but not with TECs alone or DNs alone (Fig. 4A, B).
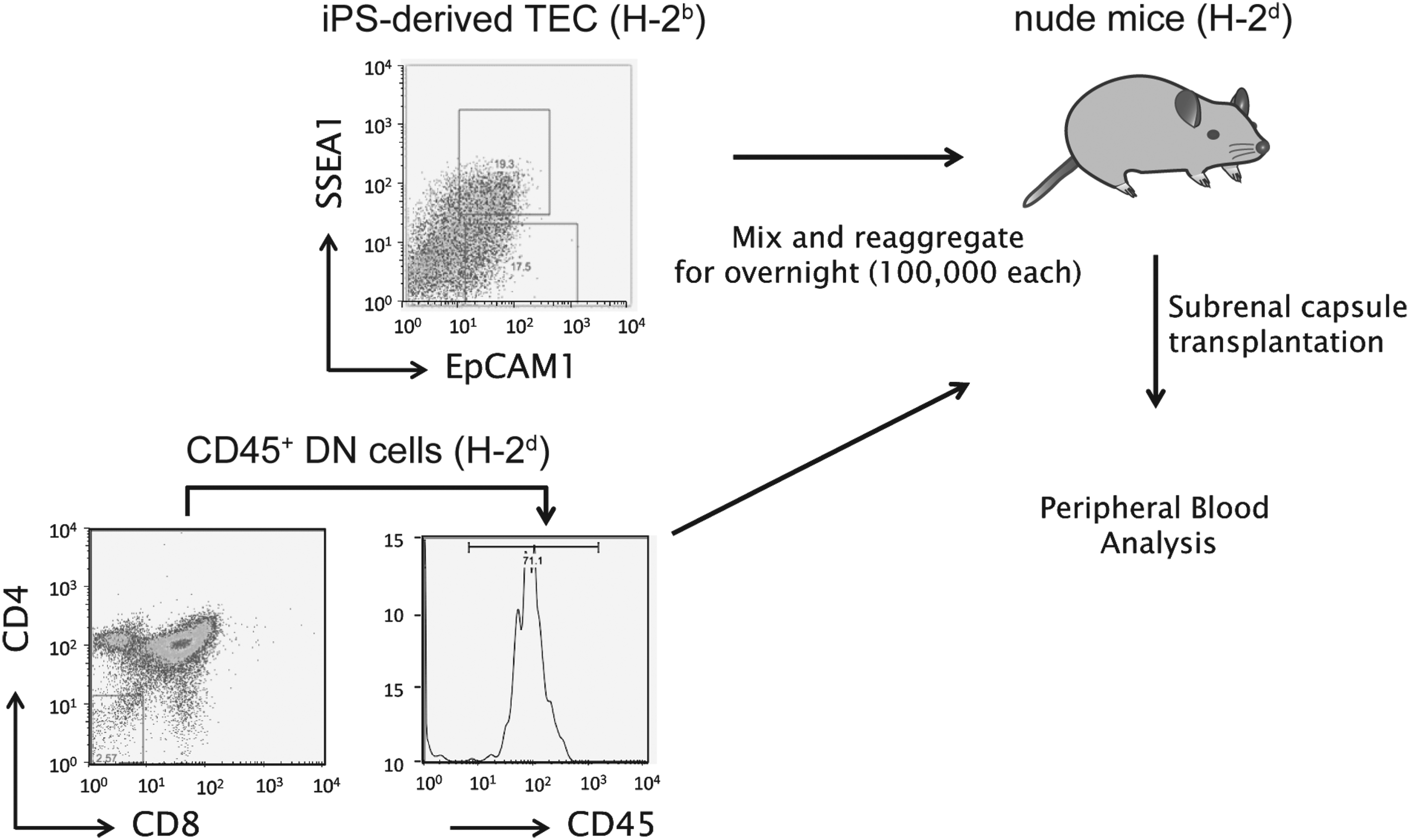
Schematic diagram for assessment of iPSC-derived TECs. One hundred thousand EpCAM1+SSEA1− iPSC-derived TECs were purified and reaggregated with 100,000 CD4−CD8−CD45+ DN cells derived from BALB/c mice in low-attachment, U-bottomed 96-well plates overnight. One day after reaggregation, cells were transplanted into the subrenal capsule of nude mice. Reaggregations of EpCAM1+SSEA1− iPSC-derived TECs (TEC only) or CD4−CD8−CD45+ DN cells (DN only) were used as a control.
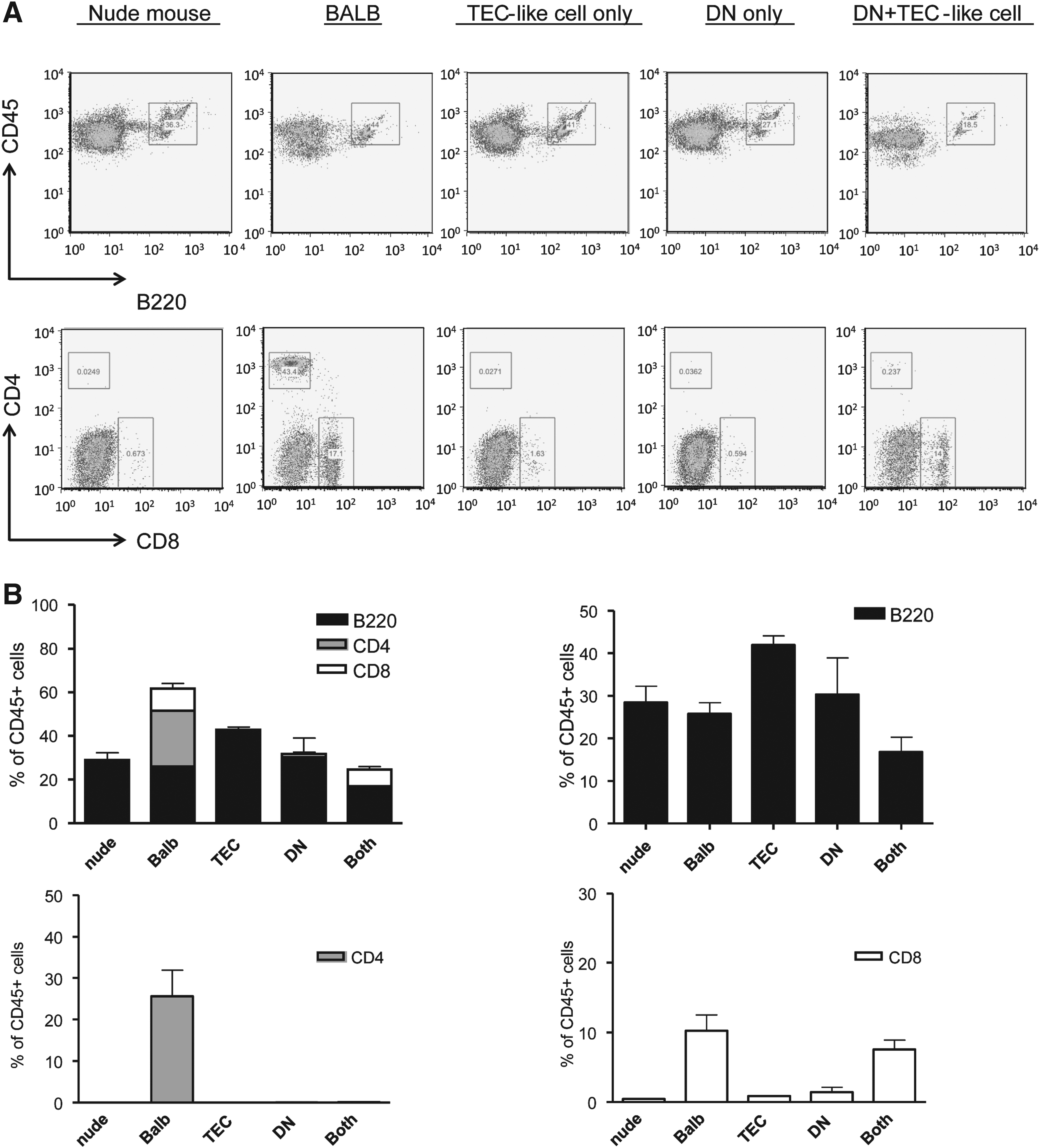
Analysis of T cell reconstitution after transplantation of iPSC-derived TECs. (
When the kidneys of the recipient mice were assessed histologically, the thymus-like structure was detected from recipient mice that were transplanted with iPSC-derived TECs mixed with DNs. No thymus-like structure was detected from the recipient mice, which were transplanted with DNs only (Fig. 5A). Interestingly, we detected all four T cell populations, such as CD4−CD8−, CD4+CD8+, CD4−CD8+, and CD4+CD8−, in thymus-like structures in kidney grafts transplanted with iPSC-derived TECs mixed with DNs. Additionally, we found that CD4+ T cells were higher than CD8+ T cells in this kidney transplant, resembling the normal thymus (Fig. 5B). These results indicate that iPSC-derived TECs have ability of supporting differentiation of both single CD4+ cells and CD8+ cells, but migration of CD4+ T cells to blood vessels was impaired for unknown reasons. To elucidate this question with further analysis, such as optimization for induction of thymic microenvironment, assessment of precise functionality of iPSC-derived TECs, and long-term observation after transplantation, are needed.
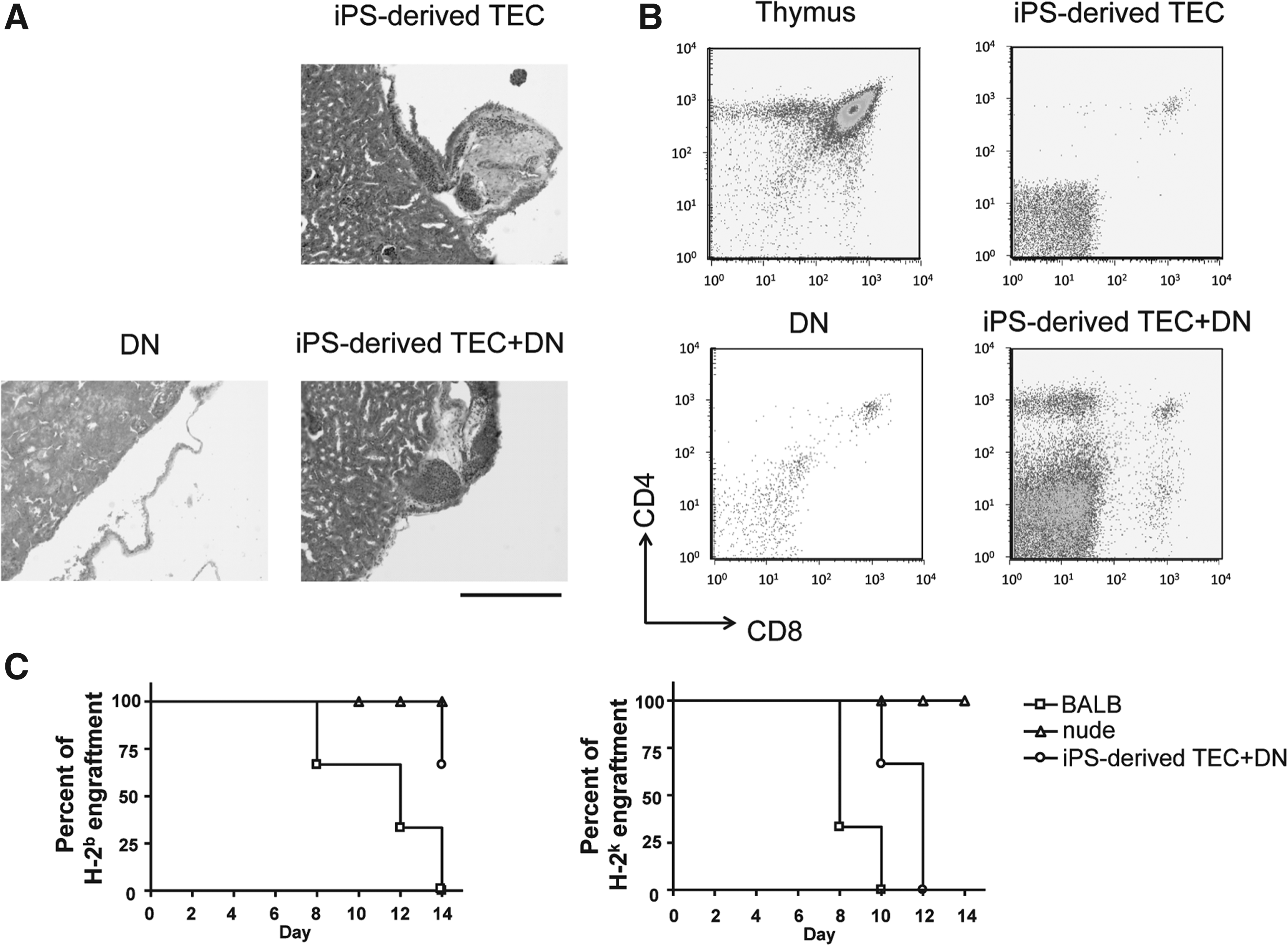
Functional analysis of reconstituted T cells in nude mice. (
T cells developed by iPSC-derived TECs were immunologically functional
Finally, we examined the immune responses of recipient mice to allogeneic skin grafts. Because allogeneic thymic grafts can induce immune tolerance to allogeneic major histocompatibility complexs (MHCs), we used the tail skin of C57BL/6 mice (H-2b), which is syngeneic to iPSCs, to assess whether T cells developed within iPSC-derived TECs are immunologically tolerant to H-2b, and we used that of C3H mice (H-2k) for assessment of rejection response to third party against nude mice (H-2d). The recipient mice showed the rejection of allogeneic C3H skin grafts within 2 weeks after transplantation, whereas grafts of C57BL/6 were not rejected, suggesting that iPSC-derived TECs induce immune tolerance to donor MHCs but not to a third party (Fig. 5C). These results indicate that our 3-D spheroid culture system can induce TECs from iPSCs that are able to develop functional T cells in nude mice.
Discussion
Although several studies succeeded in inducing TECs by 2-D culture (Hidaka et al., 2010; Lai and Jin, 2009; Parent et al., 2013; Sun et al., 2013), there are no reports to succeed in functional TEC induction from iPSCs using a 3-D culture system. Here, we reported the induction of TECs from iPSCs using a 3-D spheroid culture system, which induces development of functional T cells in allogeneic nude mice. We assessed: (1) Expression of FoxN1; (2) T cell development; and (3) functionality of T cells in recipient mice. Because the 2-D culture system can stimulate cultured cells homogeneously, it may be not sufficient to induce functional tissue, which is composed of several kinds of cells. Our culture system showed the robust expression of FoxN1 at day 10 of induction after differentiation of iPSCs; thus, we suggest that spatiotemporal cell–cell interaction in the 3-D spheroid culture may mimic the stepwise embryonic development of thymus and enhance induction of functional TECs by stimulation with activin A, BMP4, FGF8, FGF7, and FGF10. Several reports also performed the enrichment of differentiated cells from iPSC-derived spheroids for use as a disease therapeutic model (Hidaka et al., 2010; Kyba et al., 2002; Matsumoto et al., 2009), suggesting purification of target cells can be used for one of the strategic technologies to remove contamination of unexpected factors for regenerative therapy.
In addition, we could detect the both CD4+ and CD8+ T cells in the kidney graft of iPSC-derived TECs recipient mice. The reason for the lower number of CD4+ T cells in the peripheral blood of the recipient mice remains unknown. We speculate that the reaggregates of iPSC-derived TECs was partially functional to support T cell recruitment into peripheral blood. To answer this question, precise analysis of the architecture and cell population distributing the reaggregation is essential. However, it is important that the T cells differentiated by iPSC-derived TECs could reject allogeneic skin grafts with acquiring immune tolerance to donor MHC, indicating that these T cells are functional. This may enhance the future clinical applications of pluripotent stem cells to acquire cell-mediated immunity in patients with both primary and secondary immunodeficiencies. Further analysis is needed to confirm normal T cell functionality and development by assessing the T cell receptor (TCR) rearrangement, cytokine production, and differentiation of other T cell subsets.
Footnotes
Acknowledgments
We thank M. Tanaka and I. Mizuguchi for technical support, N. Oiwa for administrative assistance, and M.A. Malek for critical review of the manuscript. This work was supported by Micro-Nano GCOE program provided by the Ministry of Education, Science, Technology, Sports and Culture, Japan.
Author Disclosure Statement
The authors declare that they have no competing interests.