
Abstract
Abstract
Hematopoietic cells (HCs) and endothelial cells (ECs) can be produced in vitro from human embryonic stem cells (hESCs), but the differentiation systems used are still inefficient. To overcome this obstacle, it is necessary to understand the differentiation process. One of the methods used to obtain HCs and ECs from hESCs is their co-culture with stromal cells. The soluble factors secreted by these cells and cell–cell contact have a great impact on the differentiation process. Here, we performed comparative proteomic analyses of proteins obtained from the total extract of OP9 stromal cells and secreted by these cells before and during in vitro generation of HCs and ECs (hematoendothelial) from hESCs. We identified a total of 83 secreted and 759 intracellular proteins during differentiation. Twenty-five secreted and 181 proteins from the total extract were more abundant. Some secreted proteins are involved in cell–matrix interactions and HC and/or EC development. Moreover, 13 proteins of the total extract from OP9 cells that were exclusive/or more abundant during differentiation are involved in the Nrf2/Nfe2l2 gene pathway, that is, they are described to have a key role in oxidative stress and in hematopoietic development and maturation. Our proteomic profiles provide valuable insight about the proteins involved in in vitro hematoendothelial cell generation and in the future they might be used to optimize the differentiation process and produce both cell types in vitro.
Introduction
E
Human embryonic stem cells (hESCs) are pluripotent stem cells characterized by their unlimited capacity for self-renewal and differentiation into all of the specialized cells of the adult (Thomson et al., 1998). These cells are an important tool for producing several cell types in vitro, including ECs and hematopoietic cells (HCs). However, the therapies based on hESCs are still a challenge. For practical applications, the residual pluripotent cells that escape differentiation are potentially able to generate teratomas in vivo (Koyanagi-Aoi et al., 2013), one ability of pluripotent stem cells (Thomson et al., 1998). To overcome this obstacle, it is necessary to understand the mechanisms that promote differentiation.
In bone marrow, the function of HSCs is controlled by specialized cells and extracellular matrix components that constitute the hematopoietic niche (Schofield, 1978); stromal cells are one cell type comprising this microenvironment. Thus, the hESC co-culture system with stromal cells is a useful system for in vitro hematopoietic differentiation (Choi et al., 2009; Vodyanik and Slukvin, 2007). This differentiation model mimics the contact of hematopoietic and stromal cells, a key regulator of the cell differentiation process, and has a direct impact on the cell phenotype.
Recently, mass spectrometry (MS)-based proteomics has offered the possibility of analyzing biological processes at the molecular level. The proteomic analysis of feeder cells has enabled the identification of proteins involved in self-renewal, growth, and maintenance of pluripotency of hESCs (Buhr et al., 2007; Prowse et al., 2007; Richards et al., 2003). However, the molecules secreted by stromal cells and the molecular mechanisms by which these cells promote hematoendothelial differentiation from hESCs need to be understood better.
The purpose of this study was to perform a detailed proteomic analysis of OP9 stromal cells before and during co-culture with hESCs, as well as the secreted proteins during the differentiation process, to identify the molecules and pathways involved in the generation of hematoendothelial cells from pluripotent stem cells. Our proteomic data showed changes in the proteomic profile in OP9 stromal cells after contact with hESCs and revealed potential candidates that can regulate in vitro differentiation. Bioinformatics analysis showed that the Nrf2 pathway can be implicated in this process. Moreover, the proteins identified could be used to optimize the generation of vascular and blood cells from pluripotent stem cells.
Material and Methods
OP9 cells culture
The OP9 stromal cell line was established from C57BL/6XC3H)F2-op/op mice (Yoshida et al., 1990). In this study, we used OP9 cells modified with green fluorescent protein (GFP+). These cells were cultured in Minimum Essential Medium Alpha Medium (α-MEM) supplemented with 20% fetal bovine serum (FBS) until confluence. Then 5 × 105 cells were transferred to 100-mm3 plates coated with 0.1% gelatin and cultured for 4 days until a confluence of approximately 90%.
hESCs culture
The hESC H1 lineage derived by Thomson et al. (1998) was maintained as undifferentiated cells on mouse embryonic fibroblast (MEF) feeder cells inactivated with 10 μg/mL mitomycin C (Invitrogen). These cells were maintained in Dulbecco's F12 Modified Eagle Medium (DMEM/F-12; Invitrogen) supplemented with 15% Knockout Serum Replacement (Invitrogen), 1% nonessential amino acids (Invitrogen), 1 mM
In vitro differentiation of hESCs into endothelial and hematopoietic cells
Differentiation of hESCs into hematoendothelial cells was performed according to Vodyanik and Slukvin (2007) with some modifications. Briefly, about 1–1.5 × 106 undifferentiated hESCs were transferred to 10-cm2 plates coated with a monolayer of OP9 GFP+ cells. These cells were cultured with α-MEM medium supplemented with 10% FBS (HyClone) and 100 μM monothioglycerol (MTG, Gibco) for 9 days. The differentiation medium was replaced by fresh medium on day 1 (D + 1) and half of the medium was replaced on days 4 (D + 4) and 6 (D + 6). The entire differentiation process was performed without added cytokines.
After 9 days, all cells were harvested from the plates and digested with 0.5% trypsin-EDTA (Mediatech) solution for 10 min at 37°C and centrifuged at 300 × g for 10 min. The supernatant was discarded, and the cell pellet was treated again with trypsin-EDTA solution (0.25%) for 15 min at 37°C to individualize the cells. The OP9 GFP+ cells were separated from differentiated cells by cellular sorting using a FACSAria cell sorter (BD Biosciences). The OP9 cells were used to identify proteins involved in hematopoietic cell generation. The differentiated cells were characterized to evaluate the percentage of cells positive for endothelial and hematopoietic markers.
Characterization of differentiated cells obtained from hESCs
Phenotypic characterization of differentiated cells was performed by flow cytometry using monoclonal antibodies (BD Bioscience, MD, USA), CD34, CD43, CD45, CD31, CD144, and KDR conjugated with fluorescein isothiocyanate (FITC), phycoerythrin (PE), peridinin chlorophyll (PerCP), or allophycocyanin (APC). The control was isotype control immunoglobulin G- (γ1/γ2) conjugated probes with FITC, PE, PerCP, or APC. About 105 cells were analyzed in duplicate on a FACSCalibur Flow Cytometer (Becton Dickinson), and the events were analyzed using CELLQuestTM software (Becton Dickinson).
MS and proteomic analysis
For secretome analysis, were used 30 mL of supernatant from OP9 control cells (OP9-CTRL) and OP9 cultured with hESCs (OP9-hESCs) with 25 μL of protease inhibitor cocktail (Sigma). One day before MS analysis, the medium containing FBS was changed with medium without serum to decrease the amount of proteins from serum. The medium was concentrated to a final volume of 1 mL in an Ultracel Amicon System 15 filter unit (Millipore). For the analysis of total extract proteins from OP9-hESC and OP9-CTRL cells, 5 μL of protease inhibitor cocktail (Sigma) was added. The proteins were extracted from OP9 cells with 300 μL of lysis buffer with 0.5 M Tris-HCl (pH 6.8), 10% sodium dodecyl sulfate (SDS), and 0.1% NP-40 and subjected to three freeze–thaw cycles for cell lysis. The samples were centrifuged at 12,000 × g for 30 min at 4°C, and the supernatant was collected.
The proteins were quantified using the Bradford protein assay (Bio-Rad) and stored at −80°C; about 50 μg of protein was used for spectrometry analysis. Initially, the samples were resuspended in 40 μL of 0.1 M Tris-HCl solution (pH 8.8) containing 8 M urea and subjected to disulfide bonds reduction of proteins by the addition of 1.25 μL of dithiothreitol (DTT). The samples were incubated for 2 h at room temperature (RT) followed by alkylation with 2.5 μL of iodoacetamide and incubation for 1 h at RT in the dark. The samples were diluted five times in 0.1 M ammonium bicarbonate solution (pH ≥ 8.0) to a final volume of 290 μL and incubated with 4 μg of trypsin (Promega) at 37°C overnight. Before application in the mass spectrometer, the samples were cleaned up using an OASIS HLB cartridge 1 cc column (Waters). Briefly, the column was equilibrated with 5% acetonitrile solution containing 0.1% formic acid, and the elution was performed with 80% of acetonitrile.
The samples were dried in a SpeedVac and applied in the mass spectrometer. Proteins were identified with a mass spectrometer LTQ-Orbitrap (Thermo-Finnigan) coupled to a nanoflow liquid chromatography (LC) system (LC-MS/MS). The raw data acquired were automatically processed by the Computational Proteomics Analysis System (CPAS; Rauch et al., 2006). The peptides identified that met the criteria minimum quality (error rate <1.0%) were grouped in proteins, using the “Protein Prophet” algorithm and a list of identifications with rate <2.0% error was generated. The proteomic data were analyzed using the LABKEY Server platform and X!Tandem (UniProt database). The statistical analysis was performed using PeptideProphet and ProteinProphet software, and each sample was analyzed in triplicate. The OP9-hESC/OP9-CTRL ratio was calculated by the ratio of the mean of the normalized counts.
Bioinformatic analysis
The proteins identified were characterized using Ingenuity Pathways Analysis (IPA, Ingenuity Systems) and MetaCore™ commercial software to evaluate the biological process and pathways during hematoendothelial differentiation from hESCs.
Results
In vitro hematoendothelial differentiation from hESCs by co-culture with OP9 stromal cells
The method for hESCs differentiation into hematoendothelial cells was previously described by Vodyanik and Slukvin (2007), and it was detailed in Material and methods. The schematic representation of the differentiation process is summarized in Figure 1A.

The in vitro hematoendothelial differentiation from human embryonic stem cells co-cultured with OP9 stromal cells. (
Generation of hematopoietic progenitor cells (HPCs) and endothelial progenitor cells (EPCs) showed a complex process that involved the emergence of proliferating cell clusters and vascular tube-like formations. To start the differentiation on day 0, the H1 cells were plated on the OP9 GFP+ cells (Fig. 1B, D + 0), and on the first day, small aggregates of hESCs adhered to stromal cells (Fig. 1B, D + 1). On day 4, half of the culture medium was replaced to prevent the death of cells and to avoid differentiation of the hESCs into mesenchymal stem cells. Moreover, at this time, we could see the first colonies with mesodermal morphology (Fig. 1B, D + 4). After 2 days, the cells proliferated and formed colonies full of round cells. It was possible to see the vascular tube–like structures (Fig. 1B, D + 6). At the peak of differentiation on day 9 (D + 9), the colonies reached a larger size, and it was possible to see typical hematopoietic colonies (Fig. 1B, D + 9, left) and in highlight the vascular tube–like structures (Fig. 1B, D + 9, right). After this time, the number of HPCs and EPCs decreased due to excessive cell density and overall decline in cell viability.
The differentiated cells were collected on D + 9, and cytometric analysis showed that they were positive for hematopoietic CD34 (26%), CD43 (40%), CD45 (4.5%), and KDR (24%) and endothelial markers CD31 (18.45%) and CD144 (22%) (Fig. 1C).
Analysis and characterization of proteins found in supernatant and in cellular extract on OP9 cells during hematoendothelial differentiation
The supernatant and OP9 stromal cells were collected at the peak of differentiation on day 9. A proteomic approach based on LC-MS/MS was used to identify qualitative and quantitative differences between supernatants and cellular extracts of OP9-CTRL and OP9-hESCs during the differentiation process. In this analysis, a less than 1% false discovery rate was obtained.
We investigated possible functions of all identified proteins during hematoendothelial differentiation secreted by OP9 cells and activated in these stromal cells that can promote the generation of hematopoietic and endothelial cells. The protein list was subjected to MetaCore™ software analysis, and the results for the major biological functions are shown below. Some proteins showed more than one function.
LC-MS/MS identified 83 secreted proteins: Four were found exclusively during differentiation and 32 were found only in OP9-CTRL; 47 proteins were common before and after differentiation, in which 21 were more abundant and 26 less abundant during hematoendothelial differentiation (Fig. 2A). All of the secreted proteins are detailed in Table S1 (Supplementary Data are available at www.liebertpub.com/cell/). The analysis showed that the main biological functions of the more abundant proteins and the number of proteins are: Cell adhesion (11), biological adhesion (11), extracellular matrix organization (10), extracellular structure organization (10), and cell–substrate adhesion (6) (Fig. 2B). On the other hand, the less abundant proteins are associated with actin filament organization (15), regulation of cell death (12), exocytosis (10), platelet activation (10), and platelet degranulation (9) (Fig. 2C). The biological functions of more and less abundant secreted proteins are detailed in Table 1.
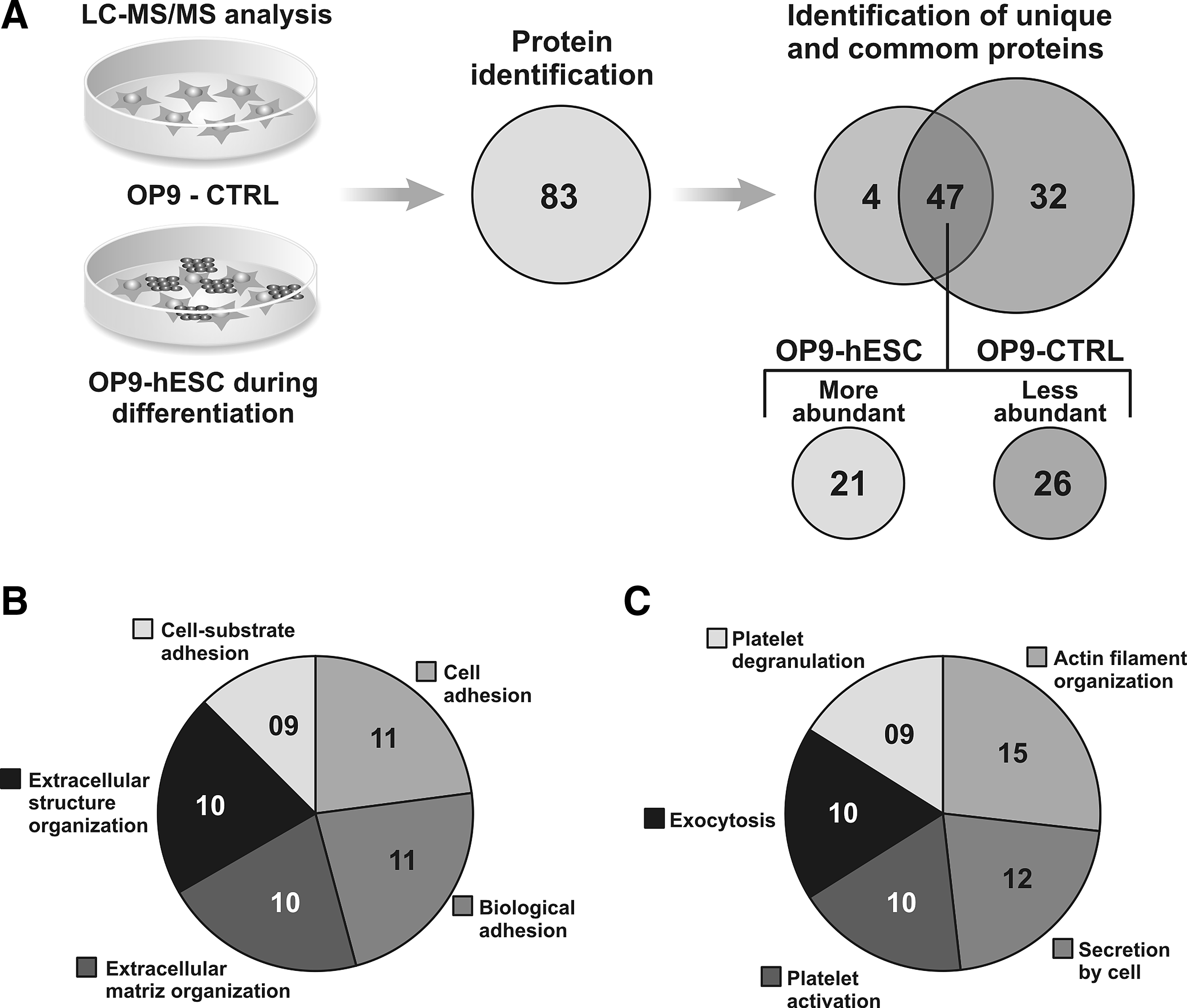
Analysis of secreted proteins by OP9 stromal cells before (OP9-CTRL) and during HC and EC generation (OP9-hESCs). OP9 GFP+ cells were separated from differentiated cells by cell sorting. (
A total of 759 intracellular proteins were identified in OP9 cells (OP9-CTRL and OP9-hESCs): 106 proteins were identified exclusivelyi OP9-hESCs, 370 proteins were found only in OP9-CTRL, and 283 proteins were common to OP9-CTRL and OP9-hESCs (75 were more abundant, 27 less abundant, and 181 unchanged) (Fig. 3A). The intracellular proteins are detailed in Table S2. The top biological functions revealed that the most abundant intracellular proteins are associated with cell-to-cell signaling and interaction, nervous system development and function, behavior (20), free radical scavenging, molecular transport, cellular development (16), protein synthesis, hereditary disorder (15), cell signaling, lipid metabolism, small-molecule biochemistry (14) and cellular movement, hematological system development and function, and immune cell trafficking (13) (Fig. 3B). The less abundant proteins identified in OP9 cells after cell–cell contact are involved in cellular assembly and organization, cellular function and maintenance, cell-to-cell signaling and interaction (15) and cellular development, cellular function and maintenance, and cellular growth and proliferation (5) (Fig. 3C). The biological functions of more and less abundant intracellular proteins are detailed in Table 2.

Analysis of intracellular proteins from OP9 stromal cells before (OP9-CTRL) and during HC and EC cells generation (OP9-hESCs). OP9 GFP+ cells were separated from progenitor hematoendothelial cells by cell sorting. (
Signaling pathways activated in OP9 cells during hematoendothelial differentiation
IPA was used to identify known regulators of identified proteins in OP9 cells and pathways associated with these molecules that could promote endothelial and hematopoietic differentiation. We found 13 more abundant or exclusive proteins during the differentiation process—Catalase (Cat), glutathione S-transferase (Gstm5), heat shock protein 90 (Hsp90b1), lamin A (Lmna), poly binding protein 1 (Pcbp1), protein disulfide isomerase associated 4 (Pdia4), protein disulfide isomerase associated 6 (Pdia6), peroxiredoxin 1 (Prdx1), Sec23a (unknown), triosephosphate isomerase 1 (Tpi1), aldehyde dehydrogenase family 3, subfamily A1 (Aldh3a1), aldolase A (Aldoa), and ATPase, alpha 1 polypeptide (Atp1a1)—that are regulated by the Nrf2/Nfe2l2 gene (Fig. 4).
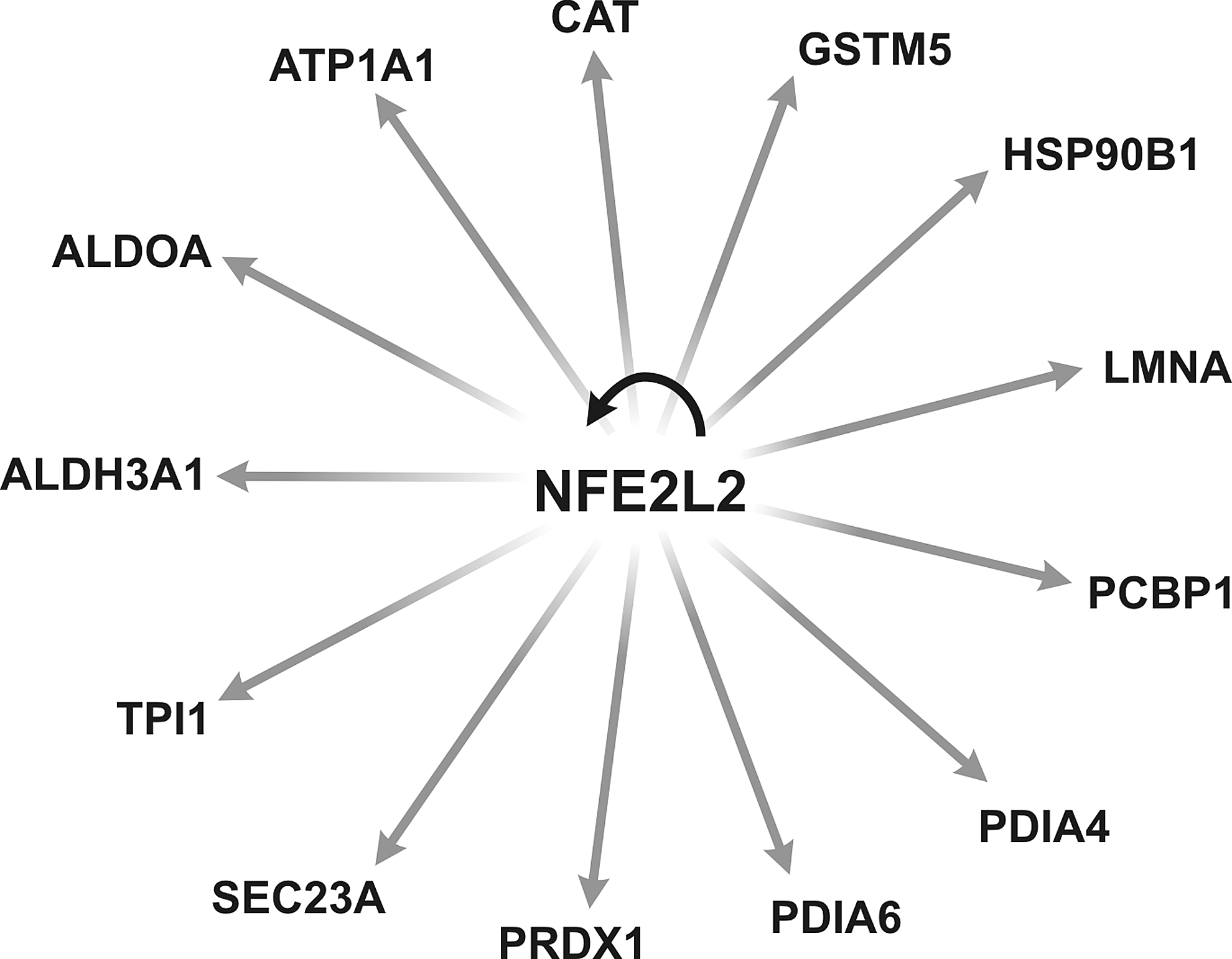
Thirteen more abundant proteins identified in OP9 cells during in vitro hematoendothelial differentiation are regulated by the Nfe2l2 gene. The list of more abundant proteins was subjected to IPA for molecular pathway analysis.
Discussion
Differentiation models from hESCs are tools for studying the mechanisms that regulate human endothelial and hematopoietic cell lineage commitment as well as the identification and function of molecules responsible for human hematoendothelial development. Stromal cells are one of the cell types that comprise the hematopoietic niche in bone marrow (Mendez-Ferrer et al., 2010). Although OP9 stromal cells are commonly used in the differentiation of pluripotent stem cells into HSCs and ECs, no study has demonstrated the molecular pathways that are activated in these cells, as well as the proteins that are secreted during hematoendothelial cells generation.
The differentiation system used here was based on the Vodyanik and Slukvin model (Vodyanik and Slukvin, 2007), which was highly reproducible and allowed generation of similar cell populations in independent experiments. Differentiated cells obtained from hESCs at the peak of differentiation on day 9 were positive for the hematoendothelial markers CD43, CD34, CD45, CD31, KDR, and CD144. Several studies have demonstrated that these markers are widely used to characterize and isolate hematopoietic and endothelial progenitors (Bhatia et al., 1998; Choi et al., 2012; Slukvin, 2013; Vodyanik et al., 2005; Vodyanik and Slukvin, 2007), with some differences in the percentage of cells obtained.
Currently, large-scale studies based on genomics and proteomics combined with bioinformatics are new in the area of hematoendothelial cell development and could provide insight into the generation of blood and vascular cells from pluripotent stem cells. In the present study, we used a sensitive and quantitative proteomic approach, LC-MS/MS analysis, to identify and quantify proteins that were changed during hematoendothelial differentiation. Our results showed the identification of secreted and intracellular proteins that were changed during the differentiation process. We identified proteins that were secreted only during endothelial and hematopoietic generation, such as angiopoietin-1 (Angp1) and Sparc-like protein 1 (Sprl1). Suri and colleagues (1996) demonstrated that Angp1 is a binder of the Tie2 receptor and one of the key regulatory molecules in the development of hematopoietic and vascular systems.
Corroborating our findings, a recent study examined the effect of various growth factors on the differentiation of murine ESCs and demonstrated that Angp1 plays a central role in the differentiation of pluripotent cells into endothelial cells (Joo et al., 2011). The Sprl1 protein is part of the extracellular matrix in the bone marrow hematopoietic niche and is critical to the matrix remodeling in niche (Delany et al., 2003). However, the role of this protein in the biology of HSCs is not completely understood. A recent study demonstrated that in the absence of Sparcl1, the HSC niche remains in a quiescent state (Ehninger et al., 2014). Our results suggest that the Sparcl1 protein is important for the generation of hematoendothelial progenitor cells in vitro, even though this protein was not necessary for the functions of murine hematopoietic cells (Siva et al., 2012). Moreover, other proteins were more abundant during differentiation, such as Fibulin2 (Fbln2), growth arrest-specific protein 6 (Gas6), versican (Vcan), fibronectin1 (Fn1), and nidogen-2 (Nid2). These proteins are involved in cell adhesion and cell–extracellular matrix interactions that have a great impact on hematopoietic cells properties in their niche (Hoggatt and Pelus, 2011; Lam and Adams, 2010; Mendez-Ferrer et al., 2010).
In a previously study, Ueno and colleagues (2003) used OP9 cells to identify important genes that were able to support HSCs culture in vitro. The authors demonstrated that the mKirre expression is important to maintaining hematopoiesis, mainly by acting directly on HSCs. In a recent study, Uenishi and colleagues (2014) cultured human pluripotent stem cells in a chemically defined medium without serum components and xenogeneic proteins to identify genes that are able to support hematopoietic differentiation. The authors showed that tenascin-C, an extracellular matrix protein, is able to promote the development of hematoendothelial and T lymphoid cells from hESCs. In our study, we used proteomics analysis to identify proteins from OP9 cells that, after hESCs contact, could be important to inducing the hematopoietic development.
The MS technique is highly sensitive and allowed identification of secreted and intracellular proteins during differentiation. The proteins of the total extract that were exclusive or more abundant during differentiation are involved in the Nrf2/Nfe2l2 gene pathway, and the proteins identified in supernatant are mainly involved in cell matrix interactions and hematoendothelial development. The differences found in these studies can be attributed to the techniques employed, time of analysis, and/or culture conditions. Also, it is important to note that different from our work, both studies mentioned above evaluated gene expression from OP9 cells and not protein.
Likewise, for secreted proteins, we performed a refined analysis of the intracellular proteins from OP9-hESCs produced during hematoendothelial differentiation to identify molecules and pathways that could be associated with in vitro endothelial and hematopoietic cells generation. After cell–cell contact, we observed a change in the protein profile that activates primordial pathways for HPC and EPC generation and not only for maintenance of the cells. IPA analysis showed that 13 proteins of the total extract from OP9 cells that were exclusive/or more abundant during differentiation from hESCs—Cat, Gstm5, Hsp90b1, Lmna, Pcbp1, Pdia4, Pdia6, Prdx1, Sec23a, Tpi1, Aldh3a1, Aldoa, and Atp1a1—are regulated by the Nrf2/Nfe2l2 gene. It was demonstrated that Nrf2/Nfe2l2 (nuclear factor-erythroid 2 or nuclear-related factor 2) is a transcription factor that has a pivotal role in the maintenance of cellular homeostasis and is considered a key regulator of antioxidant response (Dinkova-Kostova et al., 2002).
In addition to its main cytoprotective function (Motohashi and Yamamoto, 2004), Nrf2 has also been associated with hematopoiesis (Mignotte et al., 1989), controlling the production of reactive oxygen species (ROS) and as a critical mediator of cytokine signaling. Both are particularly important in our study, whereas the hematopoietic/endothelial progenitor cells are sensitive to oxidative stress (Tothova et al., 2007) and responsive to several cytokines. Although is clear Nrf2 contributes to cellular homeostasis, it is not known exactly if this gene could promote endothelial and hematopoietic differentiation from pluripotent cells, especially in models of gene differentiation in vitro based on the use of stromal cells.
Some proteins regulated by Nrf2 are important to free radical scavenging, cell signaling, lipid metabolism, small-molecule biochemistry and cellular movement, hematopoietic system development and function, and immune cell trafficking. Furthermore, Shin and colleagues demonstrated that laminin A (Lmna) contributed to cellular trafficking and erythroid differentiation (Shin et al., 2013). The proteins disulfide isomerases (PDI) on the surface of cells have the ability to rearrange, generate, or reduce disulfide bonds in proteins (Wilkinson and Gilbert, 2004). In platelets, these isomerases are implicated in aggregation, secretion, and fibrinogen binding (Burgess et al., 2000). The Cat gene encodes catalase, a key antioxidant enzyme and is associated with growth of cells, including T and B cells (Takeuchi et al., 1995). Aldehyde dehydrogenase (Aldh) activity is high in hematopoietic stem cells. Gasparetto and colleagues demonstrated that mice that lack Aldh3a1 exhibited multiple abnormalities in hematopoiesis that could be attributed to insufficient metabolism of ROS (Gasparetto et al., 2012).
Merchant and colleagues showed that Nrf−/− mice showed defective differentiation, decreased survival, and impaired engraftment of hematopoietic progenitor cells after bone marrow transplantation (Merchant et al., 2011). Furthermore, it has been recently described that Nrf2 is involved in the regulation of hematopoiesis in mice and modulates migration as well as the retention of hematopoietic cells in their niche (Tsai et al., 2013). Our data suggest the role of this gene as a regulator of hematoendothelial development from pluripotent stem cells. A detailed investigation of the role of Nrf2 as well as identified proteins during the differentiation process will be necessary to verify whether they can be used effectively to induce or regulate in vitro HPC and EPC generation. Interestingly, Nrf2 was not identified in OP9 cells-hESCs, probably due to basal conditions in which this transcription factor is quickly degraded by the ubiquitin–proteasome complex, resulting in minimal levels in the cytoplasm (Kobayashi et al., 2013). It is important to note that the proteome is highly dynamic and can vary depending on the genetic background of the cells, the method and time culture employed, and other parameters (Loring and Rao, 2006; Wagner et al., 2006).
This is the first work detailing the proteins secreted by OP9 cells and pathways activated in these cells that could be involved in hESC–hematoendothelial differentiation. The proteomic approach revealed the complexity of the in vitro generation of HPCs and EPCs revealed by identification of several proteins involved in this biological process. These proteins allowed the identification of Nrf2 pathway, but the functional role of this gene as well as the other proteins should be further evaluated to validate its importance on blood and vascular lineage commitment. The proteins identification from in vitro differentiation methods may be useful to improve the efficiency and/or generation of specific and mature cell types to be used in future in cell therapy.
Footnotes
Acknowledgments
This study was supported by Center for Cell-based Therapy (CTC) and Fundação à Pesquisa do Estado de São Paulo (FAPESP).
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.