
Abstract
Abstract
We compared the cloning efficiency of buffalo embryos produced by handmade cloning (HMC) using ear skin– and milk-derived donor cells. The blastocyst rate was lower (p < 0.05) for milk-derived than that for skin-derived embryos, whereas the total cell number and apoptotic index were similar. The global level of H3K9ac was higher (p < 0.05) in skin- than in milk-derived cells, whereas the level of H3K27me3 was similar in the two groups. The global level of H3K9ac was similar between milk-derived and in vitro–fertilized (IVF) blastocysts, which was higher (p < 0.05) than that in skin-derived blastocysts. The level of H3K27me3 was similar among the three groups. The expression level of IGF-1R and G6PD was higher (p < 0.05) in skin- than in milk-derived cells, whereas DNMT1, DNMT3a, and HDAC1 expression level was similar. In the blastocysts, the expression level of DNMT1, HDAC1, OCT4, and CDX2 was higher (p < 0.05) in skin-derived than that in IVF blastocysts. The expression level of DNMT3a and IGF-1R, was in the order (p < 0.05) skin-derived and IVF > milk-derived blastocysts and that of NANOG was (p < 0.05) IVF-> milk-derived > skin-derived blastocysts. The expression level of all these genes, except NANOG, was lower (p < 0.05) in milk- than in skin-derived or IVF blastocysts. In conclusion, milk-derived cells can be used for producing HMC embryos of quality similar to that of skin-derived embryos, although with a lower blastocyst rate.
Introduction
C
Among several biological factors that affect the cloning efficiency, the donor cell type is very important (Tian et al., 2003). A variety of donor cells from different origins (fetal, adult, etc.) and various tissues (skin, cumulus, etc.) have been used for SCNT with observed differences in the overall efficiency (for review, see Oback and Wells, 2002). Specifically, donor cells include ovarian cumulus cells, mural granulosa cells, fibroblast cells from skin, internal organs and oviduct cells, mammary epithelial cells, Sertoli cells, blood leukocytes, natural killer T cells, mature B and T cells, olfactory cells, neural stem cells and myoblasts, germ cells, embryonic stem cells (ESCs), liver cells, and colostrum-derived mammary gland epithelial cells. Recently, we have reported production of embryos by handmade cloning (HMC) from milk- (Golla et al., 2012) and semen-derived somatic cells (Selokar et al., 2014). Considerable differences have been found in the ability of different cell lines and cell types to be reprogrammed and in the developmental competence of the cloned embryos produced from them. It is still unclear which cell type is the most efficient for SCNT.
Besides cloning efficiency, another factor that needs to be considered for determining the choice of cell type is the degree of invasiveness of the procedure used for obtaining the cells from the donor animal. Because of reduced likelihood of highly invasive procedures being acceptable to the animal owner, the donor cells that can be collected in a completely noninvasive and simple manner, such as cells isolated from milk, urine, or saliva, will have an edge over other cell types that require invasive procedures for their collection. There is an urgent need to evaluate the cloning efficiency of such cell types. It is very difficult to compare the cloning efficiency of different cell types among the plethora of reports available because the biological factors, such as species, age, genotype, sex, source, cell-cycle stage, passage number of the donor cells, and technical factors, such as nuclear transfer procedure and culture conditions, all of which profoundly affect the cloning efficiency (Oback and Wells, 2002; Tian et al., 2003), have been different among these studies. To our knowledge, there is only one report in which the cloning efficiency of different cell types, all from the same donor, a 13-year-old dairy cow, was compared, keeping all the technical and biological factors constant, except the quality of recipient oocytes (Tian et al., 2003). Therefore, it is not fully understtod to what extent the donor cell influences the cloning efficiency.
Previously, we reported a method for the isolation of somatic cells from milk and their use as donor cells for production of HMC buffalo embryos (Golla et al., 2012). However, there is no information available on the epigenetic status and expression of important genes in SCNT embryos produced from milk-derived cells. Also, there is very little information available on the extent to which animal-to-animal variation affects the cloning efficiency (Shi et al., 2007). In the present study, we examined the developmental competence, quality, and global levels of important epigenetic markers, i.e., acetylation of lysine 9 at histone 3 (H3K9ac) and trimethylation of lysine 27 at histone 3 (H3K27me3), in milk cell–derived HMC embryos using ear skin–derived embryos as controls in three individual animals. We also compared the expression level of epigenetics-, pluripotency-, development-, and trophectoderm-related genes in these two types of blastocysts produced from the three individual animals, using in vitro–fertilized (IVF) blastocysts as controls.
Materials and Methods
All the chemicals and media were purchased from Sigma Chemical Co. (St. Louis, MO, USA), the disposable plasticware was obtained from Nunc (Roskilde, Denmark), and the media were purchased from GIBCO (Grand Island, NY, USA) unless otherwise mentioned. Fetal bovine serum (FBS) was obtained from Hyclone (Logan, UT, USA). Animal experiments were carried out after approval by the Animal Ethics Committee (National Dairy Research Institute, Karnal). Two types of somatic cells, i.e., ear skin–, and milk-derived cells, which had been obtained from the same animal (n = 3), and which were between passages 3–8, were used for HMC. Three healthy adult female buffaloes (Mu-5345, Mu-5365, and Mu-5476) available at the Animal Farm of the Institute were used in the present study.
Isolation and culture of milk-, and ear skin–derived somatic cells
Milk-derived somatic cells were isolated from buffalo milk as described by Golla et al. (2012) with some modifications. Briefly, milk was collected from buffaloes in sterilized plastic bottles and kept at 4°C for 1 h. The cell pellet obtained after centrifugation at 500 × g for 25 min was resuspended in 10 mL of Dulbecco's phosphate-buffered saline (DPBS) and was washed three times with DPBS containing 100 μg/ mL−1 gentamicin, 100 IU/mL−1 penicillin, 100 μg/mL−1 streptomycin, and 5 μg/mL−1 amphotericin B. The pellet was then washed twice with the culture medium, which consisted of DMEM/F12, 20% FBS, 10 ng/mL−1 epidermal growth factor (EGF), 5 μg/mL−1 insulin, 0.5 μg/mL−1 hydrocortisone, 100 μg/mL−1 gentamicin, 100 IU/mL−1 penicillin, 100 μg/mL−1 streptomycin, and 5 μg/mL−1 amphotericin B. The cells were cultured in a four-well dish in a CO2 incubator (5% CO2 in air) at 38.5°C with a change of medium every 2–3 days. Around day 7, the concentrations of gentamicin, penicillin, and streptomycin were reduced by 50% and amphotericin B was completely omitted from the culture medium. The cells were cultured with a change of medium every 2–3 days until they reached confluence, which usually took 25–30 days. Surplus cells were frozen in DMEM, 20% FBS, and 10% dimethyl sulfoxide (DMSO) and stored in liquid nitrogen for future use.
Collection of skin biopsies from the ear pinna of the animals, establishment of primary cell cultures, and donor cell preparation for HMC were performed as reported earlier (Selokar et al., 2012a).
Immunocytochemical characterization of somatic cells
The expression of cytoskeletal proteins (cytokeratin-18, keratin, vimentin, and tubulin) was examined by immunofluorescence staining. Cells that had been cultured in 96-well plates were washed three times with DPBS and then permeabilized with 0.5% Triton X-100 in DPBS for 1 h. After thorough washing with DPBS, the cells were incubated with the blocking solution (1.5% BSA) for 1 h, followed by an overnight incubation at 4°C with the primary antibody which included anti-cytokeratin-18 (1:200; cat. no. SC-32329, Santa Cruz Biotechnology, Dallas, TX, USA), anti-keratin (1:500; cat. no. MAB1611, Milipore, Temecula, CA, USA), anti-vimentin (1:200; cat. no. V6630, Sigma), and anti-tubulin (1:400; cat. no. T8328, Sigma).
For negative controls, the entire procedure was followed except that the primary antibody was replaced with mouse immunoglobulin G (IgG). After three washings with DPBS containing 0.1% Triton X-100 (0.1% DPBST), the cells were incubated with the appropriate fluorescein isothiocyanate (FITC)-labeled secondary anti-mouse antibody (1:1000; cat. no. F0257-.5ML, Sigma) for 1 h. The cells were washed three times with 0.1% DPBST followed by nuclear staining with either Hoechst 33342 or propidium iodide (PI). The cells were then examined under a fluorescence microscope (Diaphot, Nikon, Tokyo, Japan) after addition of antifade solution. Each experiment was repeated at least three times.
Effect of matrix on the growth of milk-derived cells
To compare the efficacy of different attachment factors for supporting the proliferation of milk-derived somatic cells cultured in vitro, the cells were divided into three groups and were cultured for 120 h in a 24-well dish coated with collagen matrix or MaxGel™ ECM or no attachment factor (controls). Collagen matrix and MaxGel ECM were prepared as per the manufacturer's (Sigma) protocol. Briefly, a mixture of 25 μL rat tail collagen type I/mL−1 of Dulbecco's Modified Eagle Medium (DMEM)/F12 or 10 μL of MaxGel ECM/mL−1 of DMEM was pipetted out in a 48-well plate (400 μL/well), which was incubated for 2 h at 37°C in a CO2 incubator. The medium was removed after 2 h, and fresh culture medium was added before culture of milk-derived cells. The cells were cultured for 120 h and harvested every 24 h for determination of cell number. The experiment was repeated three times.
Production of IVF embryos
Cumulus-oocyte complexes collected from abattoir buffalo ovaries were subjected to in vitro maturation (IVM) and IVF as described earlier (Sharma et al., 2011). For in vitro culture (IVC), the presumed zygotes were washed several times with Research Vitro Cleave Medium (K-RVCL-50, Cook®, Queensland, Australia) supplemented with 1% fatty acid-free bovine serum albumin (BSA) and were cultured in this medium for up to 8 days afer insemination in a CO2 incubator (5% CO2 in air) at 38.5°C.
Handmade cloning and assessment of embryo development and quality
Somatic cells were synchronized in G1 stage of the cell cycle by growing them in culture to full confluence for contact inhibition as described previously (Selokar et al., 2012a). HMC, which included IVM, cumulus/zona removal, manual enucleation, fusion, activation, and culture, was performed as described earlier (Selokar et al., 2012b). The blastocyst rate recorded on day 8 of IVC was taken as a measure of the developmental competence of embryos. For examining the quality of blastocysts, their total cell number (TCN) and level of apoptosis were determined by terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining as described earlier (Selokar et al., 2014). Cell counting was performed from the digital images obtained on an inverted Nikon fluorescence microscope. Each experiment was repeated at least three times. Apoptotic index (AI) = (number of TUNEL-positive nuclei in the blastocyst/total number of nuclei counted in that blastocyst) ×100.
Immunofluorescence staining for epigenetic markers in somatic cells and embryos
The global level of H3K9ac and H3K27me3 was examined by immunofluorescence staining in the two types of somatic cells and the blastocysts derived from them, as described earlier (Selokar et al., 2014). Briefly, somatic cells/HMC blastocysts were fixed with 4% paraformaldehyde, washed three times with DPBS and 0.3% poly(vinyl alcohol) (PVA; DPBS-P), permeabilized with 0.5% Triton X-100, and then blocked with 3% BSA. The cells were then incubated overnight at 4°C with the respective primary antibody (anti-H3K9ac, 1:1000; cat. no. H9286, Sigma; and anti-H3K27me3, 1:1500; cat. no. ABE44, Millipore) diluted in 3% BSA. After washing five times with DPBS, 0.3% PVA, and 0.1% Triton X-100 solution (DPBS-PT), the cells/blastocysts were incubated for 90 min with FITC-conjugated goat anti-rabbit secondary antibody (Sigma) diluted 1:700 in DPBS-P. After washing five times with DPBS-PT, the nuclei were counterstained with Hoechst 33342 (10 μg/mL) and rinsed with DPBS-PT. The stained cells/embryos were mounted on slides in mounting medium (glycerol with 2.5% DABCO; Sigma), and were observed under a Nikon fluorescence microscope. NIS-Elements Basic Research image processing software (Nikon, Tokyo, Japan) equipped with a microscope was used for image acquisition and quantitative measurements of the mean pixel intensity emitted by each individual nucleus. At least 10 images (50 nuclei from each image) were analyzed for each epigenetic marker.
Gene expression analysis in donor cells and embryos
Quantitative real-time PCR (qRT-PCR) was performed as described earlier (Singh et al., 2014). Briefly, RNA was isolated from the two types of donor cells and from pools of blastocysts using RNAqueous-Micro Kit (Ambion, Austin, TX, USA) as per the manufacturer's protocol. Following DNase treatment, a reverse transcription (RT) reaction was performed for cDNA synthesis using superscript reverse transcriptase III (Invitrogen). Quantification of mRNA was carried out by qRT-PCR using CFX 96 I Cycler (Bio-Rad, Hercules, CA, USA). The reaction mixture (10 μL) contained 5 μL of SYBR Green Master Mix (Maxima SYBR Green Mastermix, Thermo Scientific), 0.2 μL of 10 μM of each primer, and 1:3 diluted cDNA that had been obtained from randomly selected cells/blastocysts.
Thermal cycling conditions for all genes, were as follows: Initial denaturation at 95°C for 3 min, 40 cycles (denaturation at 95°C for 10 sec, corresponding annealing temperature and 15 sec at 72°C, and extension at 72°C for 10 sec), and melting cycle starting from 65°C up to 95°C with a 0.5°C/sec transition rate. All primer pairs used were confirmed for their PCR efficiency, and specific products were checked by melt curve analysis and for the appropriateness of size by 2% agarose gel electrophoresis. Relative levels of expression were determined using the 2−ΔΔCt method, where DCt = Ct (target gene) − Ct (internal reference), and ΔΔCt = ΔCt sample − ΔCt calibrator. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) served as the internal reference gene. Skin-derived cells/IVF blastocysts served as the calibrator for each gene. The experiment was repeated three times, each time using two replicates of the two types of donor cells/blastocysts (n = 10).
Embryo transfer and detection of pregnancy
Cycling buffaloes possessing a functional corpus luteum were treated with prostaglandin F2α (PGF2α) analog (cloprostenol sodium, 500 mg) intramuscularly. Those exhibiting estrus 72 h after the treatment were selected as recipients. Day 7/8 blastocysts produced using donor cells obtained from ear skin (n = 6) or milk (n = 4) were transferred, two to each recipient. Pregnancies were examined by transrectal palpation between days 60 and 90.
Experimental design and statistical analyses
In experiment 1, somatic cells were isolated from ear skin and milk, established in culture, and characterized. Also, the effect of matrix on the growth of milk-derived cells was investigated. In experiment 2, the two types of cells were used as donor cells for producing HMC embryos, which were compared for their developmental competence, as indicated by the blastocyst rate and quality as indicated by TCN and AI. Also, the in vivo developmental competence was examined following transfer of HMC embryos to recipients. In experiment 3, the global levels of H3K9ac and H3K27me3 were compared between donor cells of the two types and between blastocysts produced from them using immunofluorescence staining. In experiment 4, the relative mRNA abundance of some epigenetic- (DNMT1, DNMT3a, and HDAC1), pluripotency- (OCT4 and NANOG), development (IGF-1R and G6PD), and trophectoderm-related (CDX2) genes was compared between blastocysts produced from the two types of donor cells to IVF control embryos. To compare the means of different groups in the epigenetics, gene expression, and embryonic development experiments, a Student's t-test and one-way analysis of variance (ANOVA) was performed using SPSS 15 software. The post hoc test (least significant different [LSD]) was carried out to study the significant differences among means. The percentage values were analyzed after arcsine transformation. The differences were considered to be statistically significant at p < 0.05. Data were presented as mean ± standard error of the mean (SEM).
Results
Experiment 1: Establishment and characterization of skin- and milk-derived cells
The morphological appearance of ear skin– and milk-derived somatic cells is presented in Figure 1. The ear skin–derived cells expressed vimentin but not keratin and cytokeratin-18, whereas the milk-derived cells expressed all three (Fig. 2). In the experiment aimed at examining the effect of matrix on the growth of milk-derived cells, the collagen-coated surface was found to be superior to the MaxGel ECM-coated surface and uncoated plastic surface (Fig. 3). The milk-derived cells were, therefore, cultured on collagen-coated surface for all subsequent experiments.
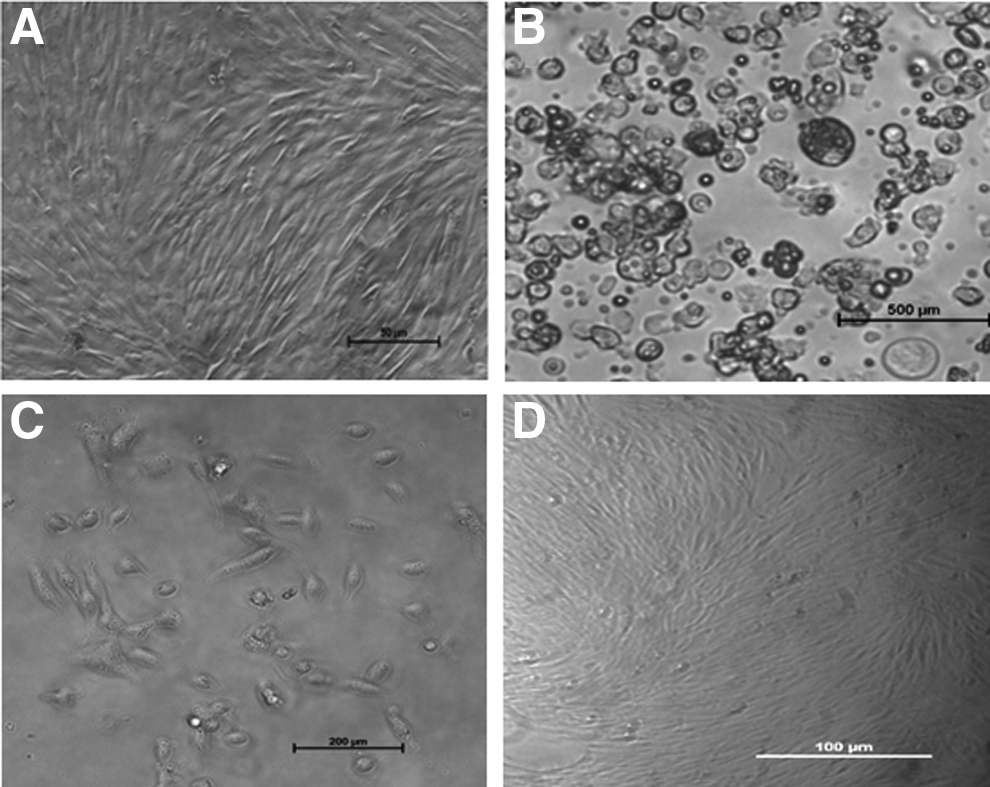
(
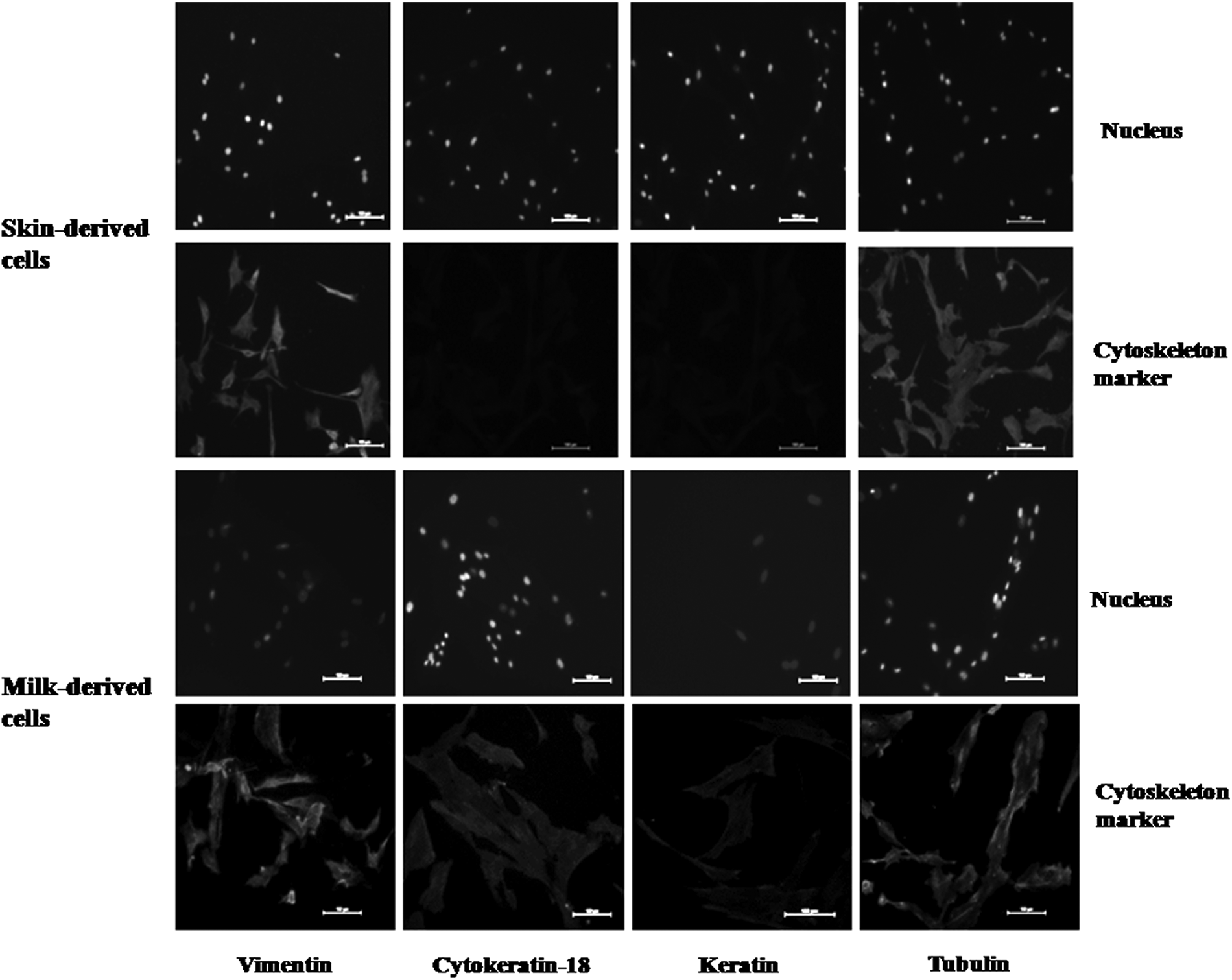
Expression of cell-specific markers in ear skin– and milk-derived somatic cells examined by immunofluorescence staining. Nucleus, nuclei of cells labeled with Hoechst 33342 stain; cytoskeleton marker, cells stained with anti-vimentin, anti-cytokeratin-18, anti-keratin and anti-tubulin (positive controls) and then with FITC-labeled secondary antibody. Expression of vimentin but not that of cytokeratin-18 and keratin showed that the skin-derived cells were of fibroblast nature; expression of all the three markers in milk-derived cells showed that they were of epithelial nature. Magnification, 200×; scale bars, 100 μm.
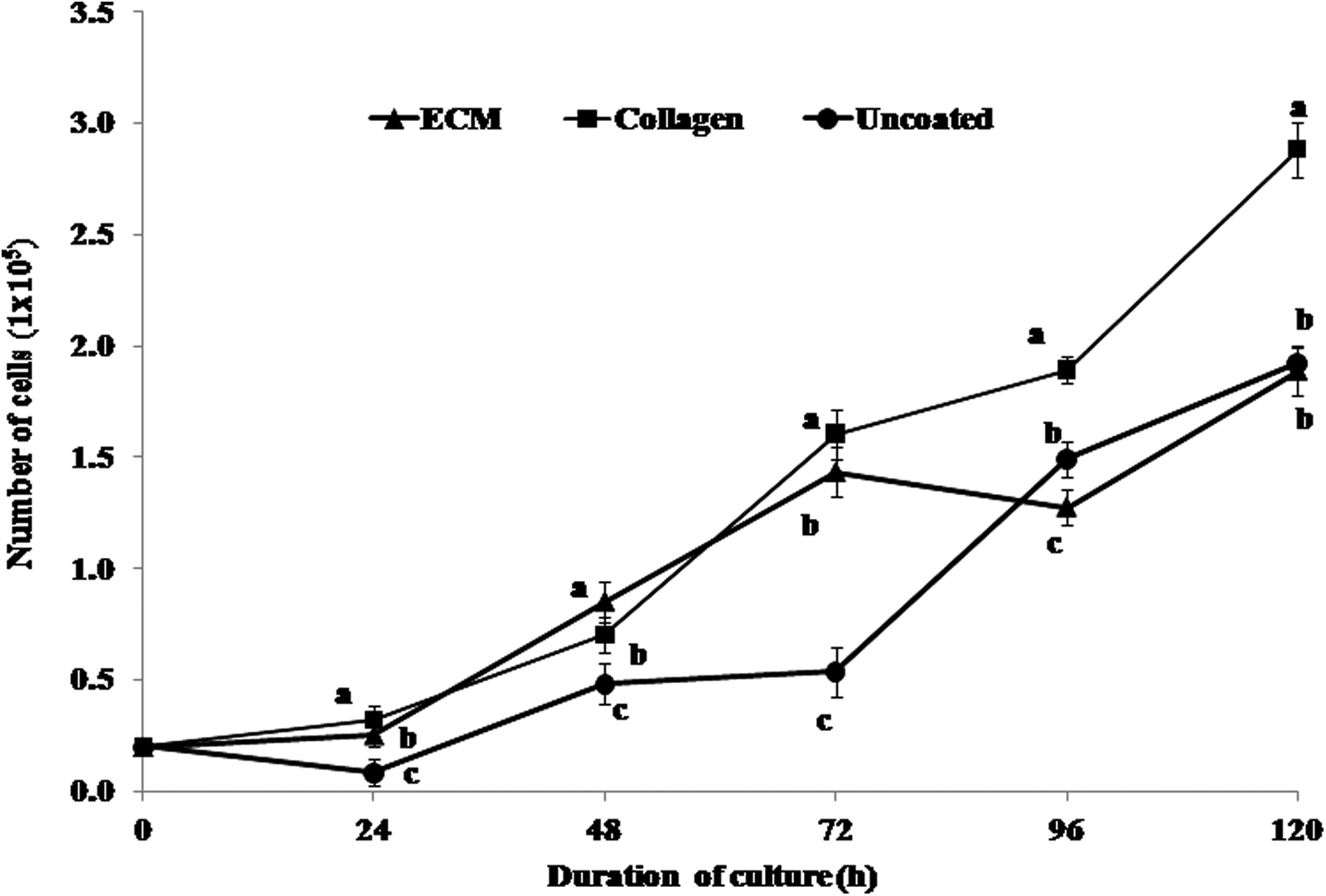
Proliferation rate of milk-derived cells following culture for 120 h in a 24-well dish coated with collagen matrix or MaxGel ECM (ECM) or no attachment factor (controls). Collagen-coated surface was found to be superior to others. Values are mean ± standard error of the mean (SEM). Points with different letters differ significantly (p < 0.05).
Experiment 2: Developmental competence and quality of HMC embryos
As shown in Table 1, the blastocyst rate was significantly lower (p < 0.05) for milk- than for ear skin–derived embryos (16.6 ± 1.91 vs. 28.8 ± 2.93%) although the cleavage rate was not significantly different (92.1 ± 2.40 vs. 90.0 ± 2.13%). TUNEL assay revealed that the TCN (153.0 ± 24.27 vs. 162.1 ± 14.07) and the AI (6.73 ± 0.75 vs. 9.01 ± 1.16) were not significantly different between the blastocysts derived from milk and ear skin cells. Neither the cleavage nor the blastocyst rate was significantly different among different individual animals for milk- and ear skin–derived embryos (Table 2).
Blastocyst rate data are from 25 trials. TUNEL data is from three trials.
Values are mean ± SEM.
Values with different superscript within the same column differ significantly (p < 0.05).
HMC, handmade cloned; TUNEL, terminal deoxynucleotidyl transferase dUTP nick end labeling; SEM, standard error of the mean.
Values are mean ± SEM.
HMC, handmade cloned; SEM, standard error of the mean.
Transfer of HMC embryos derived from ear skin cells (n = 2 each) from the three animals to recipients resulted in live birth of a healthy calf named “Lalima,” which is currently 11 months of age (Fig. 4). The genetic make-up of the cloned calf born, the donor animal, and donor cells used for producing this calf was verified by microsatellite markers analysis (Table 3). Transfer of fresh (n = 1) and vitrified-warmed (n = 3) cloned embryos produced from milk-derived cells did not result in any pregnancy.
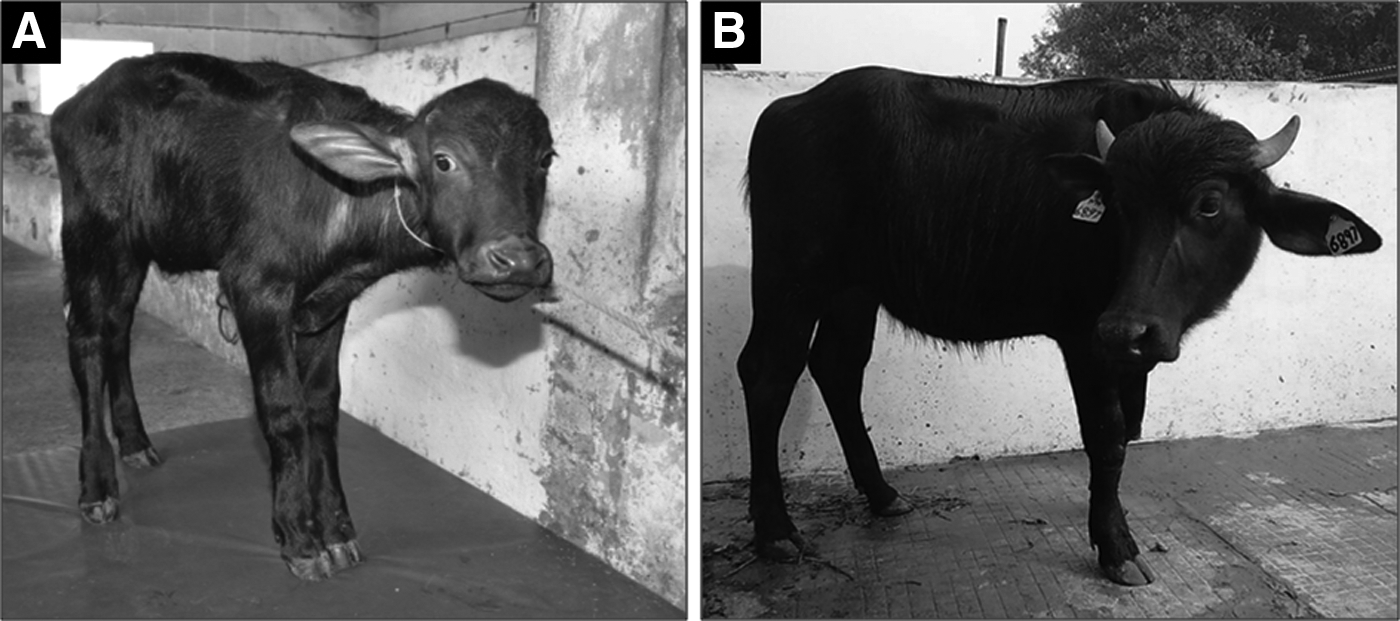
HMC calf produced using skin-derived somatic cells. (
Experiment 3: Epigenetic status of donor cells and HMC embryos produced from them
The global level of H3K9ac was significantly higher (p < 0.05) in ear skin– than in milk-derived cells, whereas there was no significant difference in the global level of H3K27me3 between the two groups (Fig. 5A). The global level of H3K9ac was similar between milk-derived HMC blastocysts and those produced by IVF, which was higher (p < 0.05) than that in ear skin–derived HMC blastocysts, whereas the level of H3K27me3 was not significantly different among the three types of blastocysts (Fig. 5B).
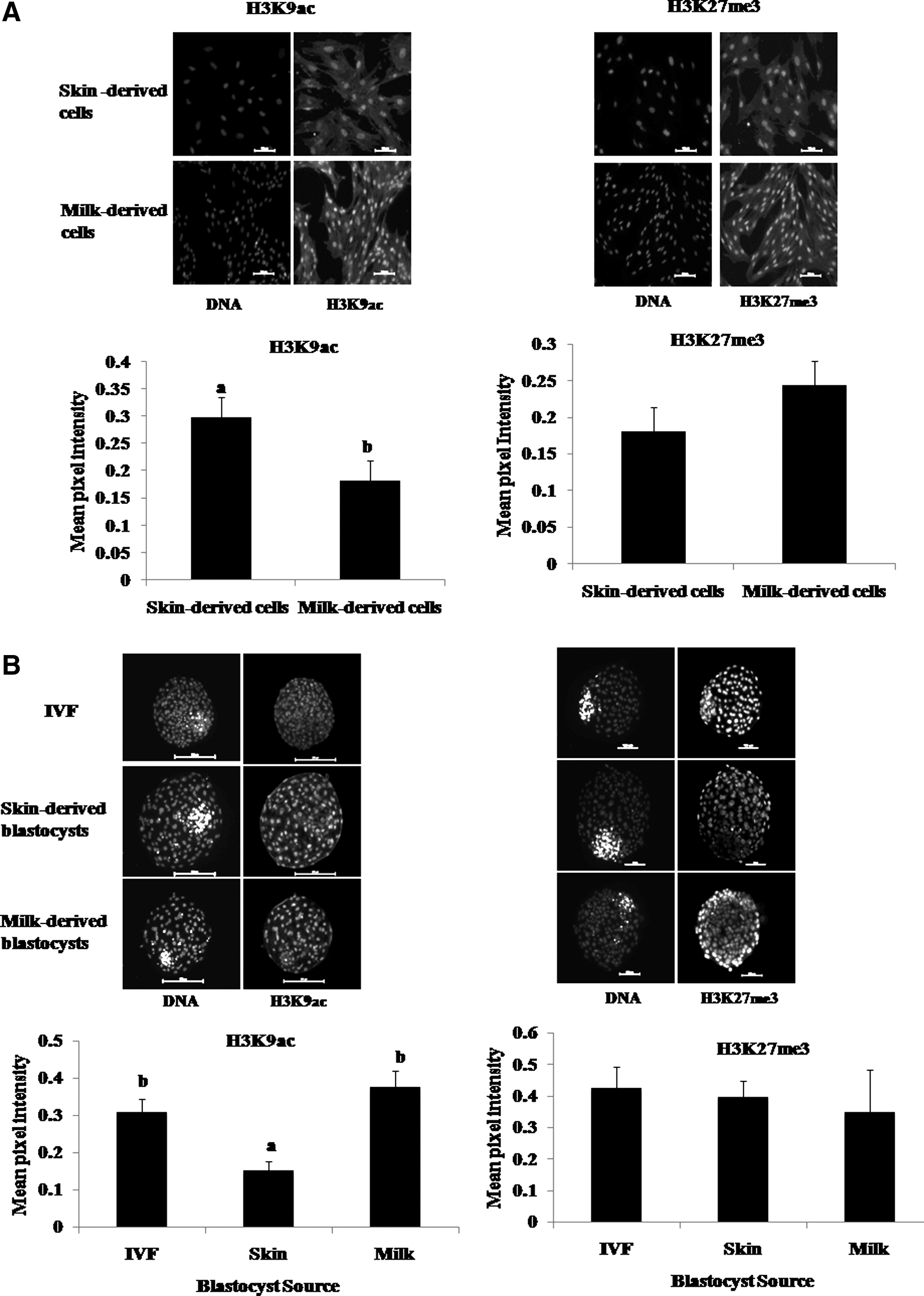
Global level of H3K9ac and H3K27me3 examined by immunofluorescence staining in ear skin– (
Experiment 4: Gene expression in donor cells and HMC embryos
The relative expression level of IGF-1R and G6PD was significantly higher (p < 0.05) in ear skin cells compared to that in milk-derived cells whereas that of DNMT1, DNMT3a, and HDAC1 was not significantly different (Fig. 6A). In the embryos derived from these cells, the relative mRNA abundance of epigenetics-related genes DNMT1 and HDAC1, pluripotency-related gene OCT4, and trophectoderm-related gene CDX2 was higher (p < 0.05) in skin-derived HMC blastocysts than that in IVF blastocysts, which, in turn, was higher (p < 0.05) than that in milk-derived HMC blastocysts (Fig. 6B). The expression level of epigenetics-related gene DNMT3a and development-related gene IGF-1R, which was similar in skin-derived HMC blastocysts and IVF blastocysts, was higher (p < 0.05) than that in milk-derived HMC blastocysts. The expression level of pluripotency-related gene NANOG was higher (p < 0.05) in IVF than in milk-derived blastocysts, which, in turn, was higher (p < 0.05) than in skin-derived blastocysts.

Relative expression level of some important genes examined by real-time quantitative PCR in milk- (
Discussion
To our knowledge, this is the first report in which the cloning efficiency, quality, epigenetic status, and gene expression levels were compared between embryos derived from two different types of donor cells, keeping the animal source constant. We found that the developmental competence of milk-derived embryos was lower than that of embryos derived from ear skin cells. However, the TCN, level of apoptosis, and the global level of H3K27me3 were similar between the blastocysts derived from these two types of donor cells. Also, there was no significant between-animal variation in the blastocyst rate for both milk- and ear skin–derived cells.
In the first part of the study, the efficacy of three matrices for supporting the growth of milk-derived cells cultured in vitro was compared. Because the collagen-coated surface was found to be superior to MaxGel ECM or uncoated surface in terms of the cell proliferation rate, the cells were cultured on a collagen-coated surface in all subsequent experiments. When the cultured cells were characterized, ear skin–derived cells were found to be of fibroblast nature because they expressed vimentin but not keratin and cytokeratin-18, whereas a majority of milk-derived cells were of an epithelial nature because they expressed all the three markers.
We had found in our earlier study that, although the cell pellet obtained after centrifugation of milk contained a mixed population of epithelial- and fibroblast-like cells, the cells were of fibroblast type following culture on uncoated plastic surface (Golla et al., 2012). Presence of epithelial cells following culture of the mixed population of cells on a collagen-coated surface in the present study suggests that collagen-coating is capable of supporting the growth of epithelial cells, which are otherwise more difficult to maintain in culture compared to fibroblasts (Freshney and Freshney, 2002). Epithelial cells obtained from sheep milk could be maintained in culture on collagen-coated dishes (Nel-Themaat et al., 2007). We found collagen-coated surfaces to be better than laminin-coated or uncoated surfaces for the culture of epithelial cells obtained from buffalo seminal plasma (Selokar et al., 2014).
In the present study, the developmental competence, as indicated by the blastocyst rate, was found to be lower for milk- than that for skin-derived embryos. This was also observed in our previous study (Golla et al., 2012). This agrees with our observation of a lower expression level of the development-related gene IGF-1R in milk- than in skin-derived HMC blastocysts. However, it is difficult to compare our results due to nonavailability of any other report on the cloning efficiency of milk-derived cells. Of the more than 200 distinct mammalian cell types known, less than 5% have been tested as nuclear donors, and all of them have been found to support development to blastocyst stage (Oback and Wells, 2002). In all the studies in which the cloning efficiency of different cell types was compared within a species, it was found to vary substantially in cattle (Kato et al., 2000), swamp buffalo (Srirattana et al., 2010), and pig (Zhang et al., 2009).
However, the nature of biological factors that determine the “clonability” of cells is currently not known. Oback and Wells (2002) attributed the following three principal reasons to the differences in the clonability of different types of donor cells: (1) Genetic or epigenetic errors in the donor genome, (2) faulty or incomplete genetic or epigenetic reprogramming, or (3) a combination of both and suggested that all the three factors may be important, but their relative roles depend on the choice of the nuclear donor.
In our study, the origin of donor cells could be an important factor that affected the developmental competence of the two types of donors. Whereas the ear skin–derived cells were of a fibroblast nature, the milk-derived cells were predominantly of epithelial origin. Skin fibroblasts have been reported to offer better cloning efficiency and live birth rate compared to mammary epithelial cells in cattle (Tian et al., 2003) and oviductal epithelial cells in pig (Zhang et al., 2009). Recently, we have shown that for the production of interspecies SCNT embryos using donor cells from wild buffalo (Bubalus arnee) and cytoplasts from domestic buffalo (Bubalus bubalis), the blastocyst rate was nearly 2.5-fold higher (p < 0.05) for fibroblasts than that for epithelial cells isolated from ear skin (Saini et al., 2015). We found that the TCN and the AI were not significantly different between milk- and ear skin–derived HMC blastocysts. Both of these are considered to be important criteria for evaluation of embryo quality because apoptotic cell death plays an important role in embryo development by eliminating cells with nuclear or chromosomal abnormalities. An increase of apoptosis in cloned blastocysts is correlated with a decrease in the TCN (Yu et al., 2007).
We found that the blastocyst rate for milk- and ear skin–derived cells was not significantly different among the three individual animals taken in the present study, suggesting that the between-animal variation may not be a major factor influencing the blastocyst rate. However, in the only report available, to our knowledge, in which the cloning efficiency of the same cell type was compared among different individual animals, the blastocyst rate of buffalo fetal fibroblasts was found to vary among the three animals examined (Shi et al., 2007).
Because epigenetic reprogramming during early development is linked to the developmental potential of cloned embryos (Santos et al., 2003), abnormality of cloned embryos is considered to be due to aberrant reprogramming (Dean et al., 2001; Kang et al., 2001). Histone modifications are an important part of reprogramming, not only because of their direct role in influencing gene expression but also because of their role in DNA methylation and the consequent effects on gene repression (Cedar and Bergman, 2009). Among histone modifications, those at histone H3 are of particular importance. In normal embryos, H3K9 methylation is reprogrammed in parallel with DNA methylation, whereas in cloned embryos it exhibits hypermethylation, which is associated with DNA hypermethylation (Santos et al., 2003). Alternatively, H3K9 can be acetylated, which is associated with active chromatin configurations (Rice and Allis 2001). H3K9ac, one the most studied epigenetic markers, is enriched in the promoter region of activated genes (Nishida et al., 2006). H3K27me3 is involved in X- chromosome inactivation in placental tissues in addition to its role in gene silencing (Kalantry et al., 2006).
We found that the global level of H3K9ac was higher in ear skin– than in milk-derived cells, whereas that of H3K27me3 was similar between the two, suggesting that the two types of cells differed epigenetically. The global level of H3K9ac was lower in ear skin– than in milk-derived HMC blastocysts and IVF blastocysts, whereas that of H3K27me3 was not significantly different among the three types of blastocysts. This suggests that the milk-derived HMC blastocysts were normal, at least in terms of the global level of these two markers. However, further studies are required to compare the global histone modifications at other positions and global DNA methylation in milk-derived cloned embryos with those produced by IVF.
The ultimate test of the quality of an embryo is its ability to result in pregnancy resulting in live birth. Because a very limited number of milk-derived embryos were transferred in our study, it is not possible to comment upon their in vivo developmental competence. In view of the ease in obtaining milk from donor animals, milk-derived cells hold a lot of potential for use as a source of donor cells for producing cloned animals. However, further work is required on transfer of a large number of milk-derived cloned embryos to recipients to determine their offspring rate. We were able to get one live birth following transfer of six embryos derived from ear skin. The calf is now 11 months old and is healthy.
One of the consequences of incomplete epigenetic reprogramming in SCNT embryos is aberrant gene expression compared to that in their IVF counterparts (for review, see Niemann et al., 2002), which results in adverse effects on their development (Santos et al., 2003). DNA methylation is one of the most important epigenetic events because genes can be repressed reversibly by a high level of methylation. It is carried out by DNA methyl transferases (DNMTs), which include DNMT1, a maintenance methyltransferase that methylates the newly formed DNA strand during S phase of the cell cycle, and DNMT3s, which are responsible for the establishment of de novo methylation during gametogenesis and early embryonic development (Bestor, 2000).
Histone acetylases (HDACs) and histone acetyl transferases, which are a part of the network that controls the acetylation status of a particular chromatin domain, play an important role in epigenetic reprogramming because the level of acetylation is correlated with the transcriptional activity of chromatin. The hyperacetylated histones are associated with transcriptionally active domains, whereas hypoacetylated histones are associated mainly with silenced chromatin regions (Morgan et al., 2005). We found the relative expression level of DNMT1 and HDAC1 to be higher in skin-derived HMC embryos than in IVF embryos, whereas that of DNMT3a was not significantly different. This is in partial agreement with an earlier report in which the relative expression of HDAC1 was observed to be higher and DNMT1 to be lower in skin-derived cloned buffalo embryos produced by conventional micromanipulation, whereas that of DNMT3a was similar between the two groups (Pandey et al., 2010).
We found that the pattern of relative expression level of all the epigenetics- (DNMT1, DNMT3a, and HDAC1) and development-related (IGF-1R) genes examined in the milk- and ear skin–derived cells was not reflected in the respective HMC blastocysts produced from these donor cells. This, combined with the fact that the global level of H3K9ac in two types of donor cells were not reflected in the respective HMC embryos produced from them, suggests that reprogramming is able to override the epigenetic status and gene expression pattern of donor cells, which may not be correlated with that of cloned embryos produced from them. The relative expression level of epigenetics-related genes DNMT1 and DNMT3a was lower in milk- than that in ear skin–derived HMC blastocysts or that in IVF blastocysts. However, further studies are required to confirm whether the lower expression level of these DNMTs is translated into a low DNA methylation level of milk-derived cloned blastocysts. Although the pattern of high expression level of DNMT1 and DNMT3 in skin- than in milk-derived HMC blastocysts was not reflected in the global level of H3K27me3, which was similar in the two types of embryos, the high expression level of HDAC1 in skin- than in milk-derived blastocysts was in agreement with the lower global level of H3K9ac in skin- than that in milk-derived blastocysts.
Proper expression of the pluripotency-related genes OCT4 and NANOG, which are expressed in the inner cell mass of blastocysts, and that of CDX2, which is expressed in the trophectoderm, is necessary for successful embryonic development. We found that the relative expression level of OCT4, NANOG, and CDX2 was different in cloned blastocysts produced from skin- and milk-derived cells than that in IVF blastocysts, suggesting that these genes are expressed in an aberrant manner in cloned embryos. This agrees with earlier reports in which the expression pattern of Oct-4 and Oct-4–related genes has been shown to be abnormal in somatic cell clones compared to that in IVF embryos (Sebastiano et al., 2005; Beyhan et al., 2007).
In conclusion, this study demonstrates that milk-derived cells can be used for producing HMC embryos of quality similar to that of ear skin–derived embryos, although with lower blastocyst rate. The epigenetic status and gene expression pattern were different among skin- and milk-derived HMC and IVF-derived blastocysts.
Footnotes
Acknowledgments
This work was funded by the Department of Biotechnology, Ministry of Science & Technology, Government of India. A.K. Raja and A.A. Sahare were supported by University Grants Commission (UGC) fellowships.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.