
Abstract
Abstract
Using of embryonic stem cells (ESCs) could improve production traits and disease resistance by improving the efficiency of somatic cell nuclear transfer (SCNT) technology. However, robust ESCs have not been established from domestic ungulates. In the present study, we generated goat induced pluripotent stem cells (giPSCs) and transgenic cloned dairy goat induced pluripotent stem cells (tgiPSCs) from dairy goat fibroblasts (gFs) and transgenic cloned dairy goat fibroblasts (tgFs), respectively, using lentiviruses that contained hOCT4, hSOX2, hMYC, and hKLF4 without chemical compounds. The giPSCs and tgiPSCs expressed endogenous pluripotent markers, including OCT4, SOX2, MYC, KLF4, and NANOG. Moreover, they were able to maintain a normal karyotype and differentiate into derivatives from all three germ layers in vitro and in vivo. Using SCNT, tgFs and tgiPSCs were used as donor cells to produce embryos, which were named tgF-Embryos and tgiPSC-Embryos. The fusion rates and cleavage rates had no significant differences between tgF-Embryos and tgiPSC-Embryos. However, the expression of IGF-2, which is an important gene associated with embryonic development, was significantly lower in tgiPSC-Embryos than in tgF-Embryos and was not significantly different from vivo-Embryos.
Introduction
E
The goat is an important domestic ungulate. Researchers have attempted to establish ESCs from goats and other ungulates, but they have not achieved much success because of differences in starting materials and limited knowledge about suitable culture systems. Many studies have demonstrated that somatic cells could be reprogrammed into induced pluripotent stem cells (iPSCs) using transcription factors (Takahashi and Yamanaka, 2006; Takahashi et al., 2007; Yu et al., 2007). The iPSCs of many domesticated ungulates have been obtained, including in goats (Ren et al., 2011), sheep (Bao et al., 2011; Li et al., 2011; Liu et al., 2012), pigs (Ezashi et al., 2009; Esteban et al., 2009; Montserrat et al., 2012; Wu et al., 2009), and cattle (Han et al., 2011; Huang et al., 2011). Mouse iPSCs and pig iPSCs have been used to generate corresponding clones using iPSCs (Zhou et al., 2010; Fan et al., 2013), indicating that iPSCs could be used as a substitute for ESCs to improve cloning efficiency. Therefore, iPSCs provide a valuable tool for generating transgenic cloned animals.
Transgenic nuclear transfer technology has an important role in livestock production and clinical medicine (Tachibana et al., 2013; Qin et al., 2015; De Bem et al., 2011). SCNT has been successfully used in our laboratory to produce transgenic cloned dairy goats from dairy goat fibroblasts that were transduced with the human lactoferrin (hLF) gene, but with a very low efficiency (Wan et al., 2012). In many mammalian species, the birthrate of cloned embryos transplanted into the recipients is only about 1–5% compared with a 30–60% birthrate for embryos that were fertilized in vitro (IVF) (Wilmut et al., 2002). The low viability of embryos and aberrations in neonates hamper the application of SCNT. Lactoferrin has some functions, including innate immunity, regulation of cell development, and iron metabolism. hLF is an iron-binding glycoprotein with multiple functions, which include a broad spectrum of antimicrobial, anti-inflammatory, and immunoregulatory activities. Compared with other lactoferrins, hLF is more suitable for humans.
In the present study, we generated goat induced pluripotent stem cells (giPSCs) and transgenic cloned dairy goat induced pluripotent stem cells (tgiPSCs),which were reprogrammed from dairy goat fibroblasts (gFs) and transgenic cloned dairy goat fibroblasts (tgFs) with four human genes (hOCT4, hSOX2, hMYC, and hKLF4), respectively. We used dairy goat fibroblasts tgFs and tgiPSCs as donor cells to produce hLF-transgenic dairy goat embryos, which were named tgF-Embryos and tgiPSC-Embryos. At the same time, we collected vivo-Embryos by superovulation. Normal expression of insulin-like growth factor IGF family genes is required for the early development of embryos. Using real-time polymerase chain reaction (PCR), we compared the expression profile of IGF family genes (IGF-1, IGF-2, IGF-1R, and IGF-2R) in tgF-Embryos, tgiPSC-Embryos, and vivo-Embryos.
Material and Methods
Ethics statement
Permission for all animal experiments was approved by the Nanjing Agricultural University Animal Care Committee (SYXK2011-0036) and Nanjing University Animal Care Committee (SCXK2010-0001) for scientific purposes.
Cell culture
Mouse embryonic fibroblasts (MEFs), primary goat ear fibroblasts, and 293FT cells were cultured in Dulbecco's Modified Eagle Medium (DMEM; high glucose) with 10% fetal bovine serum (FBS; Life Technologies), 50 U/mL penicillin/streptomycin (Life Technologies), and 0.1 mM nonessential amino acids (NEAA; Life Technologies). The giPSCs and tgiPSCs were cultured in KnockOut™ DMEM (Life Technologies) with 20% KnockOut™ Serum Replacement (KSR) (Life Technologies), 50 U/mL penicillin/streptomycin, 0.1 mM NEAA, 0.1 mM mercaptoethanol (Life Technologies), 2 mM glutamine (Life Technologies), and 20 ng/mL human basic fibroblast growth factor (bFGF; Peprotech Corporation).
The tgF-Embryos and tgiPSC-Embryos were treated with Tissue Culture Medium-199 (TCM-199; Gibco, Grand Island, NY, USA) containing 10% FBS and 5 μM ionomycin for 5 min. The tgF-Embryos and tgiPSC-Embryos were incubated in TCM-199 containing 2 mM 6-dimethylaminopurine (6-DMAP) for 4 h in a CO2 incubator at 38.5°C in an atmosphere containing 5% CO2 (Wan et al., 2012). The reconstructed embryos were cultured with synthetic oviductal fluid supplemented with amino acids (SOFaa) medium (Holm et al., 1999).
Lentivirus production and fibroblast infection
We plated 293FT cells (8 × 106 cells per T75 tissue culture flask) (Corning Corporation) and incubated them overnight in a CO2 incubator. The next day, lentiviruses encoding human OCT4, SOX2, MYC, and KLF4 were generated according to standard procedures (Sun et al., 2009). The viral supernatant was used to infect 5 × 104 gFs and tgFs, respectively.
Immunofluorescence and alkaline phosphatase assay
After fixing, permeabilizing, and blocking, cells were incubated with primary antibodies (Millipore, Billerica, MA, USA) and secondary antibodies (ImmunoReagents, Raleigh, NC, USA). The activity of alkaline phosphatase (AP) was detected using the Alkaline Phosphatase Detection Kit (EMD Millipore, Billerica, MA, USA) according to the manufacturer's instructions.
In vitro differentiation of giPSCs and teratoma formation
Colonies were detached from the MEFs by manual dissection and transferred to petri dishes (Corning Corporation) containing medium lacking human FGF-2. After 3 days of suspension culture, embryoid bodies (EBs) were transferred to gelatin-coated plates and cultured for additional 8 days. Six male severe combined immunodeficient (SCID) mice were injected with 2 × 106 tgiPSCs in dorsal flanks. One month after injection, the teratomas were excised, dissected, and fixed with 4% paraformaldehyde. Sections of paraffin-embedded tissue were stained with Hematoxylin & Eosin.
PCR analysis of tgFs, tgiPSCs, and teratomas derived from tgiPSCs
Genomic DNA was extracted from tgFs, tgiPSCs, and teratomas derived from tgiPSCs (tgiPSCs-teratomas). Genomic DNA (1 μL) was amplified using PCR in a 25-μL reaction mixture. The reaction conditions were as follows: 94°C denaturation for 5 min; 30 cycles each at 94°C for 30 sec, 62°C for 30 sec, and 72°C for 45 sec; and final extension at 72°C for 10 min (Table 1).
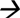 (3′)
(3′)
Nuclear transfer
Appropriate cumulus–oocyte complexes (COCs) (Chen et al., 2007) were sorted using a stereomicroscope and cultured for 20–23 h in a CO2 incubator at 38.5°C in an atmosphere containing 5% CO2 for maturation in vitro. Granulosa cells were removed from mature oocytes (Wan et al., 2012). The tgiPSCs were presumed to be at the G1 stage and selected as nucleus donors (Kou et al., 2010). The tgFs were cultured in DMEM with 0.5% FBS for 3 days to achieve synchronization. Then, one tgiPS cell nucleus and one tgF cell nucleus, respectively, were injected into two enucleated oocytes. After electrofusion, reconstructed embryos were incubated in TCM-199 containing 10% FBS at 38.5°C in an atmosphere of 5% CO2 (Wan et al., 2012).
Vivo-Embryo Collection
Estrus was synchronized in six adult Saanen and Saanen-cross goats for 10 days using intravaginal sponges with 60 mg of medroxyprogesterone acetate (XINYI Corporation); injection with 125 mg of cloprostenol (XINYI Corporation, Yancheng City, China) was administered at the time of sponge removal. Superovulation was induced with a total dose equivalent to 160 mg of follicle-stimulating hormone (FSH; XINYI Corporation) and administered twice daily in decreasing doses over 4 days. Administration was initiated at 48 h before sponge removal. A 100-mg dose of gonadotropin-releasing hormone (GnRH; XINYI Corporation) was given 36 h after sponge removal, followed by artificial insemination with fresh semen 12 h and 24 h later. Embryos were collected using surgical oviduct flushing under general anesthesia at 2 days after breeding.
RNA isolation, reverse transcriptase PCR, and real-time PCR (qPCR)
Total RNA was extracted from cells and embryos using the RNeasy Micro Kit (QIAGEN Corporation), and reverse transcription was performed using the PrimeScript™ RT reagent Kit with gDNA Eraser (Takara Corporation). The expression of genes was detected by qPCR using SYBR® Premix Ex Taq™ II (Takara Corporation). The expression of the genes was normalized to that of the internal control β-actin. The data were analyzed by analysis of variance (ANOVA) with multiple comparisons of the means in SPSS (v. 17.0, SPSS Inc. Chicago, IL, USA). p values less than 0.05 were considered significant differences statistically. The results of reverse transcriptase PCR (RT-PCR) were shown as mean ± the standard error of measurement (SEM).
Results
Derivation of giPSCs from ear fibroblasts of a dairy goat and a transgenic cloned dairy goat
Common gFs were obtained from the ear of a 3-month-old dairy goat (no. 1). The tgFs were obtained from the ear of a 6-month-old transgenic cloned dairy goat (no. 12001) (Fig. 1B, C).
Generation of giPSCs from skin fibroblasts of the dairy goat and the transgenic cloned dairy goat. (
We reprogrammed gFs and tgFs into giPSCs and tgiPSCs using lentiviruses; chemicals were not used. The time schedule of reprogramming is shown in Figure 1A. After two rounds of overnight infection with lentiviruses, the cells were split onto MEF feeders and cultured using ESC medium. Six to 7 days after transduction, two kinds of goat iPSCs colonies appeared. ESC-like colonies grew large enough for mechanical isolation from 15 to 18 days and were transferred onto 24-well plates with MEF feeders (tgiPSCs, Fig. 1D, E; giPSCs, Fig. 1F). After 5–7 days of culture, secondary colonies were passaged.
We have generated iPSCs from a normal goat but not from no. 1 (Hui et al., 2013). Here, we used giPSCs as a control to measure the reprogramming effect of tgFs. A total of 5 × 104 infected gFs and tgFs were plated onto mitomycin C–treated feeder cells. Each cell was repeated five times. The average number of colonies generated from gFs and tgFs was 82.8 ± 6.83 and 76.4 ± 8.53. The number of colonies generated from tgFs did not differ significantly from that generated from gFs (p > 0.05). The average number of AP-positive colonies generated from gFs and tgFs was 49.4 ± 2.70 and 13.8 ± 3.11, respectively. The number of AP-positive colonies generated from tgFs was significantly less than that generated from gFs (p < 0.01) (Table 2 and Fig. 1H, I).
The number of colonies generated from tgFs did not significantly differ from those generated from gFs (p > 0.05). The number of AP-positive colonies generated from tgF was significantly lesser than that generated from gF (p < 0.01).
iPSCs, induced pluripotent stem cells; AP, alkaline phosphatase; gF, dairy goat fibroblasts; tgF, transgenic cloned dairy goat fibroblasts.
The timelines for deriving giPSCs from gFs did not differ significantly from those for tgFs. No significant differences were observed between the number of ESC-like colonies generated from gFs and tgFs. However, the number of AP-positive colonies generated from tgFs was significantly fewer than that generated from gFs (p < 0.01). During the process of generating giPSCs, many partial reprogrammed colonies with loose structures appeared that lacked the capability for further expansion (Fig. 1G). Therefore, we used the tgiPSCs for further experimentation and focused on conducting a detailed characterization of tgF-derived tgiPSCs.
Expression of pluripotent genes in goat iPSCs
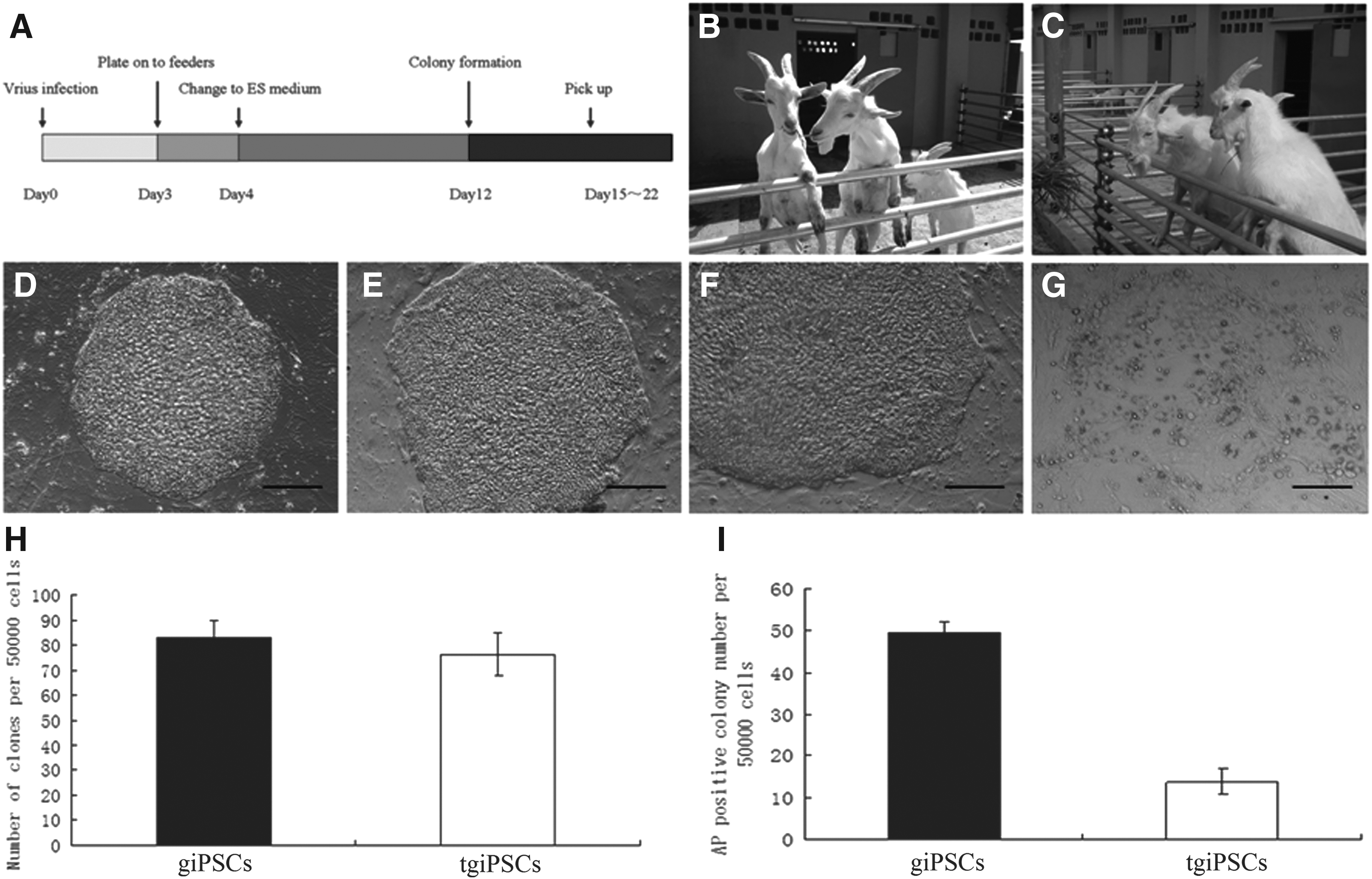
We used gFs and tgFs as negative controls and used eight- to 16-cell goat embryos (vivo-Embryos) as positive controls because of the lack of goat ESC lines. RT-PCR analyses of three giPSCs and three tgiPSCs revealed that the expression levels of the goat endogenous pluripotency genes NANOG and KLF4 were comparable to those of vivo-Embryos. The endogenous goat pluripotency gene KLF4 was highly expressed in gFs and tgFs. The endogenous pluripotency genes OCT4 and SOX2 were expressed in giPSCs and tgiPSCs, but at lower levels than those of vivo-Embryos (p < 0.01). The goat endogenous pluripotency gene MYC was expressed in giPSCs at the same level as that of vivo-Embryos, but the expression of MYC in tgiPSCs was lower than that of vivo-Embryos (p < 0.01). Moreover, the pluripotency genes SOX2 and NANOG were expressed at very low levels in gFs and tgFs, respectively. These observations suggest that endogenous pluripotency genes, including gOCT4, gSOX2, gMYC, and gNANOG, could be reactivated by reprogramming (Fig. 2).
RT-PCR analyses of endogenous OCT4, SOX2, cMYC, NANOG, and KLF4 expression in giPSCs, tgiPSCs, and vivo-Embryos. Transcript levels were normalized to those of β-actin. Error bars represent standard deviations of relative values compared to vivo-Embryos (n = 3). (*) p < 0.05; (**) p < 0.01.
Immunocytochemistry, AP assay, and karyotype analysis
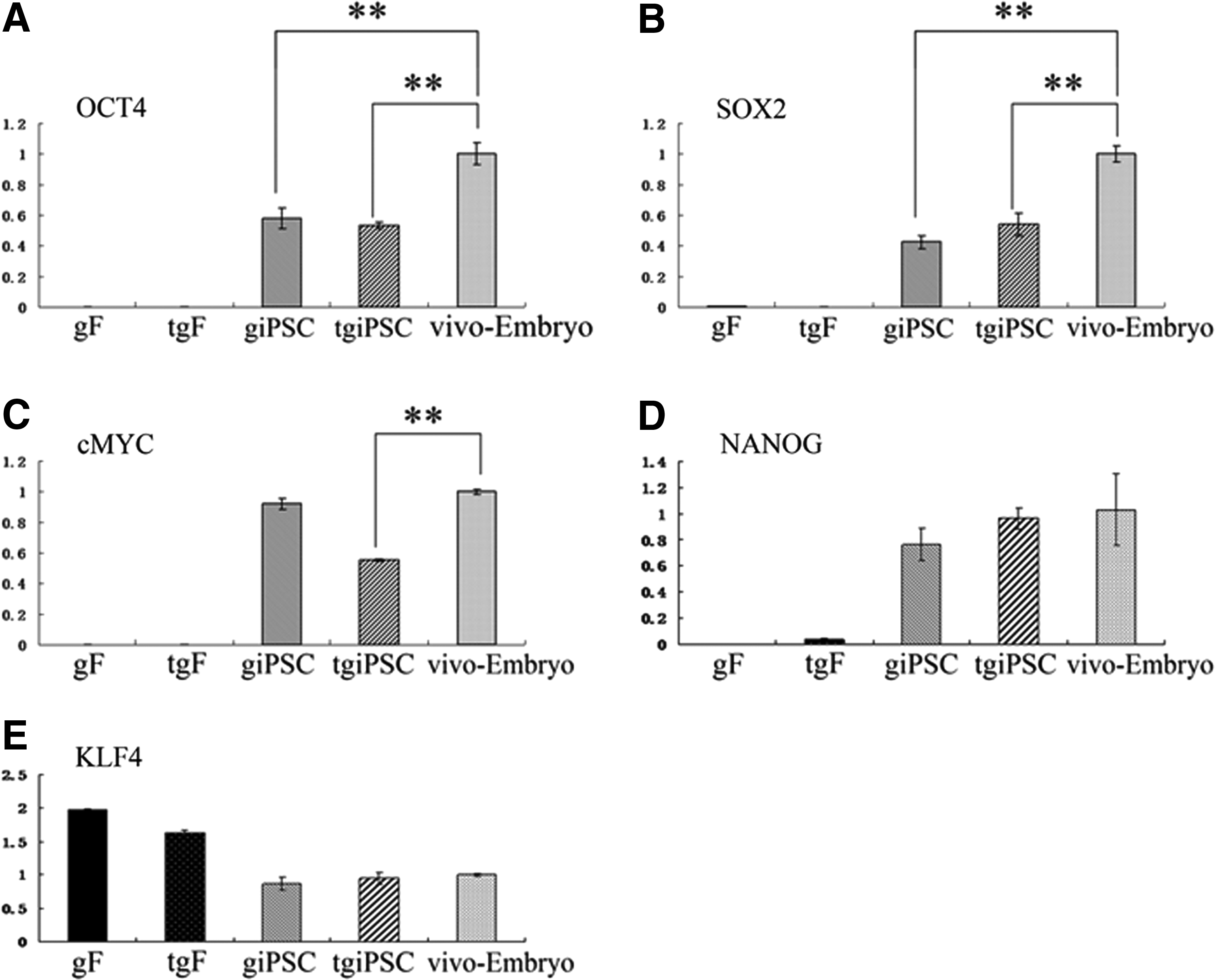
The result of immunofluorescence revealed that giPSCs and tgiPSCs expressed OCT4, NANOG (giPSCs, Fig. 3A, C; tgiPSCs, Fig. 3E, G), and AKP (Fig. 3M, N). giPSCs and tgiPSCs were incubated only with a secondary antibody, which did not show florescence (Fig. 3I, K), and possessed normal karyotypes (60XY) after 18 passages (Fig. 3O, P).
giPSCs and tgiPSCs express pluripotency markers. Immunofluorescence staining shows the expression of pluripotent markers OCT4 (
giPSCs spontaneously differentiate into three germ layers
The giPSCs and tgiPSCs derived from gFs and tgFs have the potential to form EBs containing three germ layers, which were indicated by the results of qRT-PCR (Fig. 4A–C). Furthermore, tgiPSCs could form teratomas in vivo comprising derivatives from three germ layers—gut-like epithelium (endoderm, Fig. 4D), adipose tissue (mesoderm, Fig. 4E), cartilage (mesoderm, Fig. 4F), and glycogenated squamous epithelium (ectoderm, Fig. 4G).
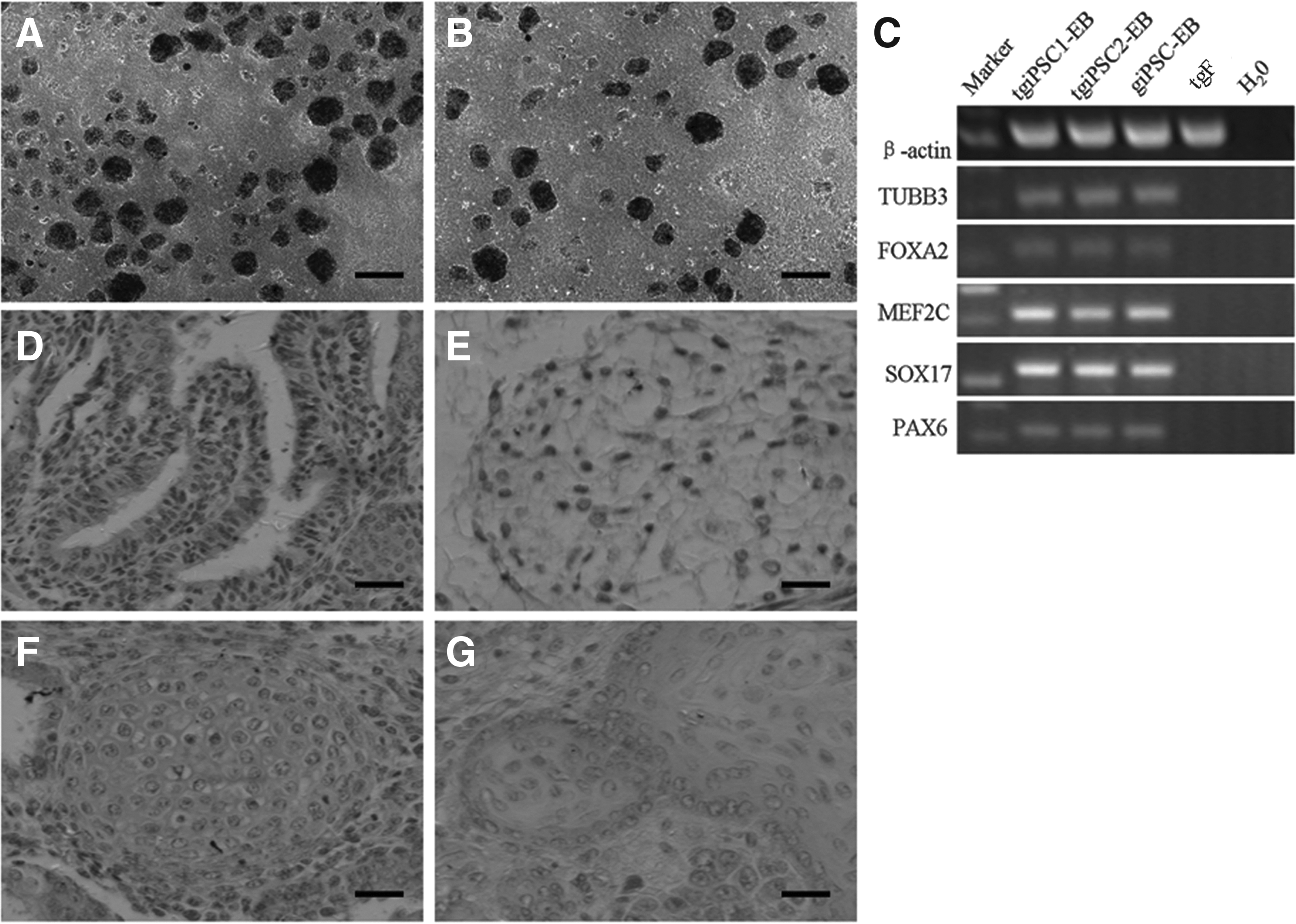
tgiPSCs and giPSCs spontaneously differentiate into three germ layers. tgiPSCs (
Effect of tgFs and tgiPSCs on the production of embryos in vitro
The tgFs and tgiPSCs were used as donor cells to produce NT embryos (Fig. 5A, B). Overall fusion rates of tgF-Embryos and tgiPSC-Embryos were 86.3 ± 2.7 (119/138) and 87.1 ± 3.6 (138/158), respectively. Overall cleavage rates of tgF-Embryos and tgiPSC-Embryos were 86.6 ± 1.2 (103/119) and 87.7 ± 1.3 (121/138), respectively. Thus, fusion rates and cleavage rates had no significant differences between the two kinds of embryos (p > 0.05; Fig.5G and Table 3). We analyzed the expression of IGFs (IGF-1, IGF-1R, IGF-2, and IGF-2R) among the tgF-Embryos, tgiPSC-Embryos, and vivo-Embryos using RT-PCR. At the eight- to 16-cell embryo stage, the expression of IGF-1, IGF-1R, and IGF-2R was not significantly different among tgF-Embryos, tgiPSC-Embryos, and vivo-Embryos. In contrast, the expression of IGF-2 in tgiPSC-Embryos was significantly lower than that in tgF-Embryos (p < 0.01). However, the expression of IGF-2 was not significantly different between tgiPSC-Embryos and vivo-Embryos (Fig. 5C–F).
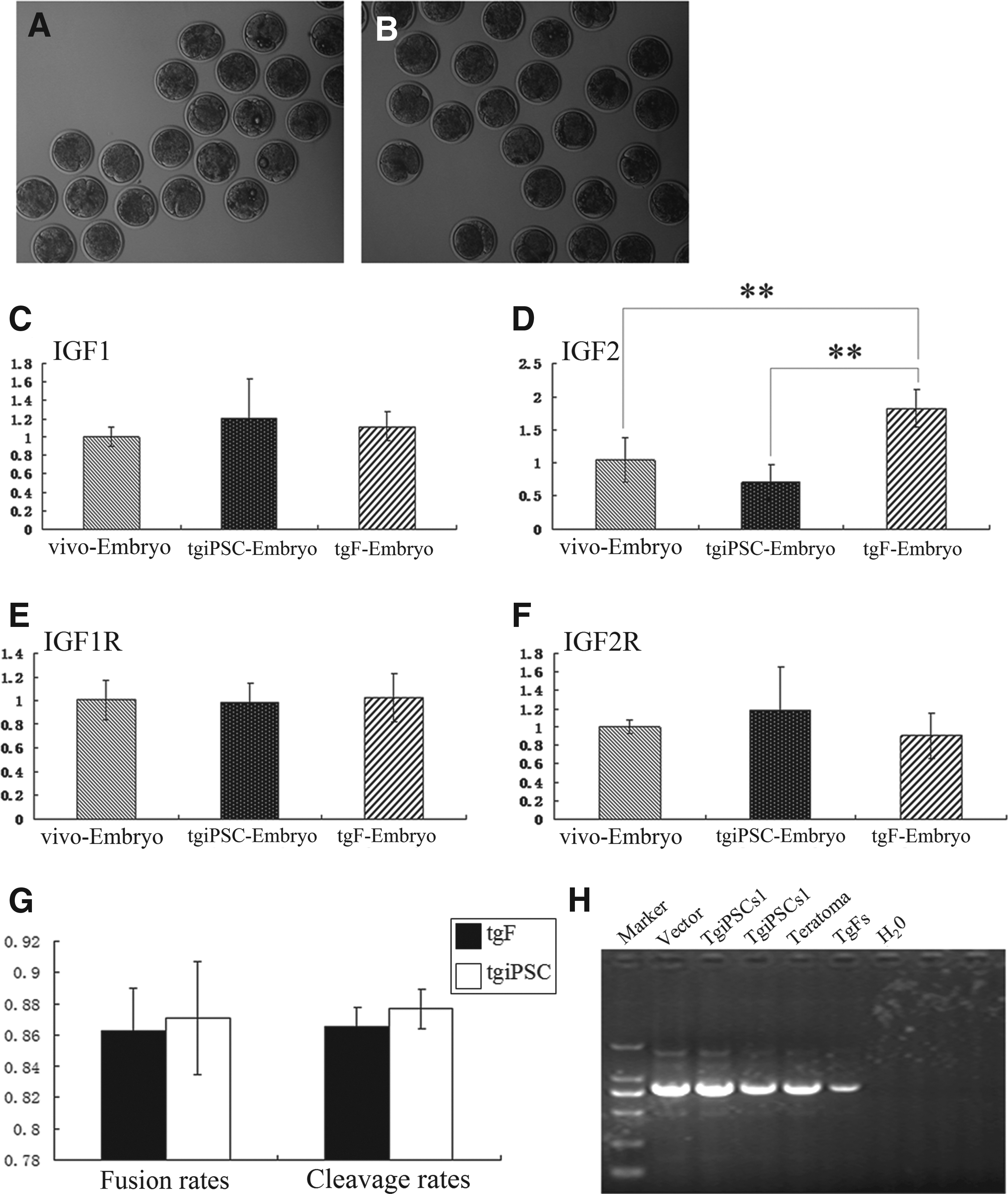
tgFs (
The overall fusion rates and cleavage rates of tgF-Embryos were not significantly different from those for tgiPSC-Embryos (p > 0.05 for both).
NT, nuclear transfer; tgF, transgenic cloned dairy goat fibroblast; tgiPSC, transgenic cloned dairy goat-induced pluripotent stem cell.
PCR analysis of tgFs, tgiPSCs, and teratoma
Genomic DNA was extracted from tgFs, tgiPSCs, and teratomas and analyzed using PCR, which demonstrated the presence of the hLF transgene in the genomes of tgFs, tgiPSCs, and teratoma cells (Fig. 5H).
Discussion
ESCs could be used in genetic engineering for improving the efficiency of SCNT technology, which would help to improve the production traits of domesticated ungulates (Donovan et al., 2005; Keefer, 2004; Rideout et al., 2000; Wall et al., 2005). However, efforts to establish robust ESCs of domesticated ungulates have been hampered because of difficulties in isolating and culturing stable ESC lines from domesticated ungulates (Keefer et al., 2007). iPSCs offer a good alternative to ESCs in this regard, and the generation of stable iPSCs of domesticated ungulates is a novel approach for establishing pluripotent stem cells.
SCNT techniques have been used successfully to produce cloned embryos and offspring in several species, but efficiencies of the procedures are low extremely (Bavister, 2004). Accumulating evidence suggested that epigenetic reprogramming in the SCNT embryo was defective (Dean et al., 2001; Mizutani et al., 2012; Kang et al., 2001; Inoue et al., 2006; Ohgane et al., 2004; Santos et al., 2003). These abnormal epigenetic modifications were likely associated with insufficient reprogramming of somatic nuclei, resulting in the overall low success rate of cloning (Fukuda et al., 2010; Egli et al., 2011; Kishigami et al., 2006; Noggle et al., 2011; Kohda et al., 2012).
Domesticated ungulates such as pigs (Ezashi et al., 2009; Esteban et al., 2009; Montserrat et al., 2012; Wu et al., 2009), cattle (Han et al., 2011; Huang et al., 2011), sheep (Bao et al., 2011; Li et al., 2011; Liu et al., 2011), and goats (Ren et al., 2011) have been used as sources for iPSCs. However, goat iPSCs lost pluripotency after transgene silencing (Ren et al., 2011), and derivation of stable giPSCs is still required.
Here, we generated giPSCs and tgiPSCs from the ear fibroblasts of a dairy goat and a transgenic cloned dairy goat. The number of AP-positive colonies generated from tgFs was significantly lower than those from gFs (p < 0.01). Thus, it is more difficult to generate tgiPSCs than giPSCs. It was also difficult to determine the gap between the pluripotency of iPSCs and corresponding totipotency in many studies because of the lack of a good positive control (Bao et al., 2011; Li et al., 2011; Liu et al., 2012; Ren et al., 2011). Here, we used goat embryos at the eight- to 16-cell stage produced in vivo as a positive control. The results of RT-PCR indicated that the expression of the endogenous pluripotency genes OCT4 and SOX2 in giPSCs and tgiPSCs was lower than that of vivo-Embryos (p < 0.01). The expression of the endogenous pluripotency gene MYC in tgiPSCs was lower than that of vivo-Embryos (p < 0.01).
The expression of other endogenous pluripotency genes was not significantly different among giPSCs, tgiPSCs, and vivo-Embryos (p > 0.05 for all). The pluripotent giPSCs and tgiPSCs were defective in their ability to acquire totipotency. This finding is consistent with the results of many reports (Aasen et al., 2008; Balasubramanian et al., 2009; Kim et al., 2009; Li et al., 2009; Tan et al., 2011). For example, significant differences in expression of endogenous pluripotency genes between iPSCs and the corresponding ESCs were not found (Hiratsuka et al., 2011; Kane et al., 2010). However, ESCs were considered to be totipotent only in some cell lines (Beddington and Robertson, 1989; Ralston and Rossant, 2005). Whether mouse and primate ESCs could be considered truly totipotent is debatable, depending on their capability to form all cell types, including placental tissue. Most ESCs do not serve as rigorous positive controls for measuring the expression of endogenous pluripotency genes in the corresponding iPSCs. However, immunofluorescence staining of giPSCs and tgiPSCs demonstrated that they expressed AKP, OCT-4, and NANOG. TgiPSCs could differentiate into three germ layers in vivo and in vitro.
We detected hLF in the genome of tgiPSCs, and thus tgiPSCs could be directly used as donor cells to produce transgenic cloned dairy goats harboring hLF to avoid re-transgenics. The tgFs and tgiPSCs were used as donor cells for producing hLF-transgenic dairy goat embryos (tgiPSC-Embryos). We collected tgF-Embryos, tgiPSC-Embryos, and vivo-Embryos at the eight- to 16-cell stage for two reasons. First, we wanted to use the vivo-Embryos as a positive control to detect the expression of endogenous pluripotency genes in tgiPSCs and giPSCs. Each cell in the vivo-Embryos at the eight- to 16-cell stage exhibited robust totipotency. Second, the expression of IGFs and their receptors decreased significantly during early development until activation of the embryonic genome at the eight- to 16-cell stage, when an increase was observed (Pandey et al., 2009; Yaseen et al., 2001).
We compared the expression profile of growth-promoting genes (IGF-1 and IGF-2) and their receptor genes (IGF-1R and IGF-2R) in tgF-Embryos, tgiPSC-Embryos, and vivo-Embryos using RT-PCR. IGF-1 regulates the growth-promoting activity of growth hormone through IGF-1 receptors. The bioactivities of IGF-2 are similar to those of IGF-1 (Kim et al., 2005; Shen et al., 2013). The expression of IGF-1 and IGF-2 significantly affects the reprogramming of the derived embryos and their subsequent development (Kim et al., 2006). At the eight- to 16-cell stage, the expression of IGF-1, IGF-1R, and IGF-2R was not significantly different among tgF-Embryos, tgiPSC-Embryos, and vivo-Embryos, but the expression of IGF-2 was significantly lower in tgiPSC-Embryos than that in tgF-Embryos. However, there were no significant differences in the expression of IGF-2 between tgiPSC-Embryos and vivo-Embryos.
The expression of IGF-2 in embryos that were generated using SCNT was relatively higher than that in vivo-Embryos. This finding could account for the abnormal development of SCNT embryos and the production of large offspring [large offspring syndrome (LOS)] (Han et al., 2003; Miyoshi et al., 2010). The expression profile of IGFs family genes (IGF-1, IGF-2, IGF-1R, and IGF-2R) in tgiPSC-Embryos was closer to that of vivo-Embryos than tgF-Embryos.
In the present study, IGF-I mRNA was detected in tgF-Embryos, tgiPSC-Embryos, and vivo-Embryos. This result was consistent with the observations for preimplantation bovine and mouse embryos (Doherty et al., 1994; Lonergan et al., 2000; Schultz et al., 1992; Watson et al., 1992; Yoshida et al., 1998) in contrast to other reports in which IGF-I mRNA was not detected in preimplantation embryos of mice (Rappolee et al., 1992), rats (Zhang et al., 1994), water buffaloes (Daliri et al., 1994), and humans (Lighten et al., 1997).
Footnotes
Acknowledgments
This study was supported by grants from the National Major Special Projects on New Cultivation for Transgenic Organisms (grant no. 2014ZX08008-003) and National Natural Science Foundation of China (grant no. 31272443)
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.