
Abstract
Abstract
Intrastriatal transplantation of dopaminergic neurons has been shown to be a potentially very effective therapeutic approach for the treatment of Parkinson's disease (PD). With the detection of induced pluripotent stem cells (iPSCs), an unlimited source of autologous dopaminergic (DA) neurons became available. Although the iPSC-derived dopaminergic neurons exhibited most of the fundamental dopaminergic characteristics, detailed analysis and comparison with primary DA neurons have shown some aberrations in the expression of genes involved in neuronal development and neurite outgrowth. The limited outgrowth of the iPSC-derived DA neurons may hamper their potential application in cell transplantation therapy for PD. In the present study, we examined whether the forced expression of L1 cell adhesion molecule (L1CAM) and polysialylated neuronal cell adhesion molecule (PSA-NCAM), via gene transduction, can promote the neurite formation and outgrowth of iPSC-derived DA neurons. In cultures on astrocyte layers, both adhesion factors significantly increased neurite formation of the adhesion factor overexpressing iPSC-derived DA neurons in comparison to control iPSC-derived DA neurons. The same tendency was observed when the DA neurons were plated on postnatal organotypic striatal slices; however, this effect did not reach statistical significance. Next, we examined the neurite outgrowth of the L1CAM- or PSA-NCAM–overexpressing iPSC-derived DA neurons after implantation in the striatum of unilaterally 6-hydroxydopamine (6-OHDA)-lesioned rats, the animal model for PD. Like the outgrowth on the organotypic striatal slices, no significant L1CAM- and PSA-NCAM-enforced neurite outgrowth of the implanted DA neurons was observed. Apparently, induced expression of L1CAM or PSA-NCAM in the iPSC-derived DA neurons cannot completely restore the neurite outgrowth potential that was reduced in these DA neurons as a consequence of epigenetic aberrations resulting from the iPSC reprogramming process.
Introduction
T
Today, induced pluripotent stem cell (iPSC)-derived DA neurons are considered the most promising source for DA neuron grafts for PD patients. iPSCs are obtained by the reprogramming of somatic cells, such as skin fibroblasts (Takahashi and Yamanaka, 2006; Takahashi et al., 2007), which can be collected from the PD patient via a small skin biopsy. The autologous origin of the iPSC-derived DA neuron graft makes immunosuppression superfluous. Because iPSCs have an unrestricted capacity for self-renewal, an unlimited number of cells can be obtained for transplantation purposes.
Various strategies have been developed to differentiate the pluripotent cells toward a ventral midbrain (VM) DA neuron fate (Chambers et al., 2009; Kawasaki et al., 2000; Perrier et al., 2004). DA neurons derived from iPSCs show properties resembling primary VM DA neurons, such as the expression of the specific markers Lmx1A, Nurr1, TH, Pitx3, and Girk2 (Yang et al., 2008), the spontaneous or stimulated release of dopamine, and spontaneous action potential firing with a slow pace (1–10 Hz) (Kriks et al., 2011; Zhang et al., 2010). However, when grafted into the striatum of 6-hydroxydopamine (6-OHDA) unilaterally lesioned rats, the animal model for PD, the neurites from the iPSC-derived DA neurons appear to be restricted within the injection tracks (Yang et al., 2008) or only shortly extending from the site of the graft (Kriks et al., 2011).
Recently, we performed an in-depth comparative analysis of the expression profile of pure mouse iPSC-derived DA neurons with that of pure mouse primary DA neurons (Roessler et al., 2014). We used Pitx3-green fluorescent protein (GFP) mice to enable the isolation of pure iPSC-derived DA neurons and pure primary VM DA neurons with GFP-based fluorescence-activated cell sorting (FACS). Besides comparative global gene expression mapping, we performed comprehensive DNA methylation profiling by reduced representation bisulfite sequencing (RRBS). The mouse iPSC-derived DA neurons largely appeared to adopt characteristics of their in vivo counterparts in morphology, dopamine production, global gene expression, and CpG island (CGI) methylation profiles, although some fibroblast genes still appeared to be expressed (Roessler et al., 2014). However, we also found deviations in CGI methylation for a subpopulation of genes that clearly effected predominantly the expression of genes associated with functional annotations, such as “nervous system development,” “neurogenesis,” “neuron differentiation,” and neurite outgrowth” (Roessler et al., 2014).
It is likely that these iPSC reprogramming “artefacts” are responsible for the limited outgrowth and survival of iPSC-derived DA neurons after implantation in the rat PD model. To achieve significant functional improvement in this animal model, a substantial reinnervation of the affected putamen and caudate is essential. In contrast to pluripotent stem cell-derived DA neurons, primary fetal VM DA neurons have been shown to be able to reinnervate the grafted striatum extensively (Stromberg et al., 1986), providing the proof-of-principle of the beneficial functionality of DA neuron grafts in PD patients (Lindvall et al., 1990; Piccini et al., 2000). Even when 10 times more tyrosine hydroxylase-positive (TH+) cells were implanted, pluripotent stem cell–derived DA neurons gave rise to a much lesser extent of striatal reinnervation than primary fetal VM DA neurons (Kirkeby et al., 2012).
An approach to improve the outgrowth of iPSC-derived DA neurons and to overcome the neurite outgrowth restrictions due to the iPSC-reprogramming could be the forced overexpression of cell adhesion molecules that are highly expressed in the developing fetal brain, in particular L1 cell adhesion molecule (L1CAM) and polysialylated neuronal cell adhesion molecule (PSA-NCAM). L1CAM and PSA-NCAM are crucial cell adhesion molecules for embryonic neuron development and axon guidance. During the fetal development of VM DA neurons, the PSA-NCAM content increases strikingly in both the developing striatum and the mesencephalon, with TH+ cells clearly outlined by PSA-NCAM (Shults et al., 1992). L1CAM is also expressed in fetal VM DA neurons and particularly mostly enriched in the TH+ fibers in the medial forebrain bundle (MFB) (Torre et al., 2010). It has been demonstrated that specific stimulation of the L1CAM signaling pathway in VM tissue grafts by L1 antibody (Neomarkers, Lab Vision, Fremont, CA, USA) resulted in a significantly greater area of graft-derived innervation compared to control grafts (Marchionini et al., 2003).
In the present study, we aimed to examine the effect of L1CAM or PSA-NCAM (by transduction of the gene encoding for STX, the enzyme that adds PSA onto NCAM) overexpression on the neurite outgrowth of mouse iPSC-derived DA neurons in vitro as well as in vivo after transplantation in the rat PD model. By generating iPSCs from the fibroblasts of Pitx3GFP/+ transgenic mouse, we were able to obtain a pure population of DA neurons after their specific differentiation using FACS.
Materials and methods
iPSC culture and DA neuron differentiation
Mouse Pitx3GFP/+ iPSC clones CI, CII, and CIII (passages 15–52) were maintained on γ-irradiated mouse embryonic fibroblasts in ES Medium [Knockout DMEM, 15% knockout serum replacement, 1% nonessential amino acids (all Invitrogen, Breda, The Netherlands, www.invitrogen.com); 2 mM
The DA differentiation protocol for mouse iPSCs was based on the MS5 protocol (Barberi et al., 2003). iPSC colonies were dissected manually and plated on MS5 cells in KSR Medium [Dulbecco's modified Eagle medium (DMEM), 15% knockout serum replacement, 100 μM β-mercaptoethanol, 100 U/mL penicillin, 100 μg/mL streptomycin] from day 2 to 5. Sonic hedgehog (SHH; 200 ng/mL, PeproTech, Rocky Hill, NJ, USA) and fibroblast growth factor-8 (FGF8;100 ng/mL, PeproTech) were added to the medium from day 6 to 8 of differentiation. Medium was changed into N2 medium [DMEM/F12 (Invitrogen), 2% N2 supplement (PAA Laboratories), 2 mM
RT-PCR and qPCR
Total RNA was extracted from transduced iPSCs or differentiated iPSCs using a standard RNeasy Kit (QIAGEN, Venlo, The Netherlands). Primers were used for L1, STX, Nestin, Pax6, Map2, β-III-tubulin, Nurr1, Pitx3, TH, and DAT. Primer sequence information is presented in Table 1. Relative gene expression levels were determined by qPCR real-time PCR (LightCycler) using the QuantiTect™ SYBR® Green PCR (QIAGEN) LightCycler® Kit according to the manufacturer's instructions. For each reaction, 0.1 ng of total RNA was used as input. Relative expression levels were normalized to the housekeeping gene hydroxymethylbilane synthase (HMBS).
Quantitative dopamine determination by liquid chromatography-tandem mass spectrometry (LC-MS/MS)
The dopamine production by the iPSC-derived DA neurons was analyzed as described earlier (van de Merbel et al., 2011). We analyzed 100 μL of supernatant from terminally DA-differentiated iPSCs under stimulation by 56 mM KCl.
Immunostaining
Cell cultures of iPSC-derived cells were fixed with 4% paraformaldehyde (PFA) for 15 min and washed three times with phosphate-buffered saline (PBS) before staining. Antigen retrieval was performed on brain cryosections with a 10 mM sodium citrate solution for 10 min heated in a microwave. Immunostaining was performed for the following markers—neuronal markers: Rabbit anti-tyrosine hydroxylase (1:500; cat. no. AB152, Chemicon, Millipore, Amsterdam, The Netherlands), mouse anti-GFP (1:500; cat. no. MAB3580, Millipore, Billerica, MA, USA), mouse anti-L1CAM (1:400; cat. no. ab24345, Abcam), mouse anti-PSA-NCAM, and immunoglobulin M (IgM, 1:400; cat. no. MAB5324, Millipore). Samples were incubated for 24 h with primary antibodies diluted in PBS with 0.05% Triton at 4°C, and primary antibodies were detected with appropriate secondary antibodies coupled to Alexa Fluor 594 or Alexa Fluor 488 (1:800; Jackson ImmunoResearch, West Grove, PA, USA) for 2–3 h at room temperature. Samples were counterstained for 10 min with Hoechst to visualize cell nuclei.
Fluorescence-activated cell sorting
Differentiated iPSC-derived cells were dissociated using the Papain Dissociation Kit (cat. no. LK003150, Worthington Biochemical, Lakewood, NJ, USA). Dissociated cells were collected in colorless DMEM (Gibco) with BDNF, ascorbic acid, GDNF, TGFβ3, and dbcAMP (see above), and sorted on a MoFlo-XDP or MoFlo-Astrios sorter (Beckman-Coulter, Brea, CA, USA) with a 100-μm nozzle at a pressure of 15–17 psi based on GFP expression. Cells were collected in N2 medium with BDNF, ascorbic acid, GDNF, TGFβ3, and dbcAMP. Cells were centrifuged at 300 relative centrifugal force (rcf) for 10 min and resuspended in PBS.
Gene transduction
Sorted GFP+ iPSC-derived DA neurons were transduced with lentiviral vectors containing genes encoding for human L1CAM, human STX (ST8Sia II), and puromycin resistance under the elongation factor-1 alpha (EF-1α) promoter; an empty vector was used as control.
Lentiviral particles were produced by co-transfecting human embryonic kidney (HEK) 293T cells (Invitrogen, Carlsbad, CA, USA) with the lentiviral vector pCMV Δ8.91 and vesicular stomatitis virus G protein (VSV-G) plasmid using Fugene (FuGENE-HD; Roche, Almere, The Netherlands) in a six-well plate following the procedure provided by the manufacturer. The ratio used was 3 μL of FuGENE to 1 μg of DNA for each well. After overnight transfection, culture medium was changed to N2 medium; 24 h later, media containing virus and 4 μg/mL Polybrene (Sigma-Aldrich, Zwijndrecht, The Netherlands) were mixed and filtered through a 0.45-μm filter (Whatman, The Netherlands). The viral particles were further concentrated 100-fold by ultracentrifugation (20,000 × g, 20 min). The pellet was resuspended in N2 medium.
For transduction, N2 medium containing lentiviral particles was added to the resuspended sorted cells in a prelubricated EP tube. After 4 h of transduction, cells were washed twice by adding N2 medium, centrifuged at 300 rcf for 10 min, and resuspended in PBS for grafting in 6-OHDA rats or in N2 medium for culturing on an astrocyte layer (4000 cells/well) or on organotypic striatal slices (400 cells/slice). Three days after transduction, the cells were incubated with 0.5 μg/mL puromycin for 24 h for selective survival of only transduced cells. After 1 week of culturing, samples were taken for qPCR analysis or for immunocytochemistry staining.
Culture of primary astrocytes
Brain tissue was explanted from P1–P3 mice, meninges were peeled off, and the brain tissue was chopped into small pieces and incubated with TE buffer at 37°C for 30 minutes. Glia medium [DMEM, with 10% FBS, 1% sodium pyruvate (Invitrogen), 100 U/mL Pen/Strep] was added to stop the digestion, and tissue was pipetted into cell suspension and cultured at 37°C under 5% CO2. Medium was changed every 3–4 days. The flasks of cell culture were shaken at 150 rpm, 37°C for 1 h, for three times every other day to remove microglia. The remaining astrocytes were passaged into a 24-well culture plate as monolayer for sorted iPSC-derived DA neurons.
Culture of organotypic striatal slices
Organotypic striatal slice cultures were performed with a modification of a previously published method (Cavaliere et al., 2010). Striata were dissected from P1–P3 mice, cut on a vibratome coronally at 250 mm, and collected in ice-cold F12 medium. Slices were cultured at the liquid–air interface on Millicell CM Cell Culture Inserts (Millipore) and maintained in Neurobasal Medium supplemented with 0.5% B27 supplement, 2 mM
Neurite tracing and analysis
Neurite tracing was performed with simple neurite tracer in the software program Fiji (http://fiji.sc/Simple_Neurite_Tracer). The total neurite length represented the sum of the lengths of all neurites of one TH+ neuron. The number of primary neurites per neuron represented the number of neurites directly extending out from a TH+ soma. The number of branches per neuron comprised the total number of neurites traced minus the number of primary neurites. The effective area comprised the area of the polygon created by linking up the far ends of all neurites of one neuron using the polygon selection option in Fiji.
Lesions, rotation behavior, and transplantation surgery
Adult female Sprague-Dawley (Harlan) rats (180–230 grams) were housed under standard conditions with free access to food and water. Rats were anesthetized with ketamine (90 mg/kg) and xylazine (4 mg/kg). Unilateral retrograde destruction of dopaminergic neurons in the substantia nigra was induced by two stereotaxic injections of 2.5 μL of 6-OHDA (3 mg/mL in 0.2% ascorbic acid and 0.9% saline, Sigma) in the MFB of the nigrostriatal pathway (Stereotaxic coordinates 1, AP −4.0, ML −0.8, and V −8.0; toothbar set at +3.4. Coordinates 2, AP −4.4, ML −1.2, and V −7.8; toothbar set at −2.4). The unilateral destruction of the nigrostriatal pathway was confirmed by the recording of
Rotation behavior was evaluated 4 and 8 weeks after grafting. Daily subcutaneous (s.c.) injections of cyclosporine A (15 mg/kg; Sigma-Aldrich) were given, starting 24 h before cell grafting (double dosage) and continued until the rats were sacrificed and perfusion-fixated at 9 weeks after cell grafting.
All animal experiments were carried out according to the Dutch Regulations for Animal Welfare. The protocol was approved by the Institutional Animal Care and Use Committee of the University of Groningen.
Tissue fixation and histology
Under ketamine/metedomidine anesthesia, rats were perfused transcardially with 4% PFA in PBS. Brains were explanted, postfixed in 4% PFA for 24 h, and soaked in a 20% sucrose solution for 1 day. They were sectioned (14-μm sections) on a cryostat after embedding in O.C.T. compound (Sakura Finetek, Torrance, CA, USA).
Statistical analysis
The data presented of neurite outgrowth of DA neurons on astrocyte or organotypic slices were from three independent experiments (n = 3). In each independent experiment, 8–15 cells were analyzed in each group. All tests were performed and analyzed in a blinded manner. Throughout the text and in the figures, all values are expressed as the mean ± standard error of the mean (SEM). SPSS analysis of difference in means between two groups of independent samples was performed using the Student t-test. Differences in means among multiple data sets were analyzed with one-way analysis of variance (ANOVA) by Tukey post hoc analysis. In all analyses, p values of less than 0.05 were considered significant.
Results
Dopaminergic neuron differentiation of iPSC lines
iPSCs were generated from embryonic fibroblasts isolated from Pitx3GFP/+ transgenic mice. The lineage-specific differentiation of three iPSC clones into midbrain DA neurons was performed according to previously described protocols (Barberi et al., 2003) The appearance of Pitx3-expressing DA neurons in the differentiating iPSCs could be detected by their expression of GFP; at day 20 of differentiation, most of the Pitx3-GFP+ cells co-expressed TH (Fig. 1A). GFP expression under control of the Pitx3 promoter allowed purification of the differentiated DA neurons with FACS (Roessler et al., 2014). The highest yield of GFP+ cells (1.78%) after sorting from iPSC clone III (CIII) is shown in Figure 1B. mRNA was collected and analyzed with RT-PCR. The purified Pitx3-GFP+ cells of iPSC clone CIII appeared to express the proper profile of DA neuron markers, i.e., both neuronal markers (Map2 and β-III-tubulin) and midbrain DA neuron markers (Nurr1, Pitx3, TH, and DAT) (Fig. 1C). Measurement of KCl stimulated secretion of dopamine revealed dopamine release in the clone CIII-derived Pitx3-GFP+ cells (Fig. 1D).
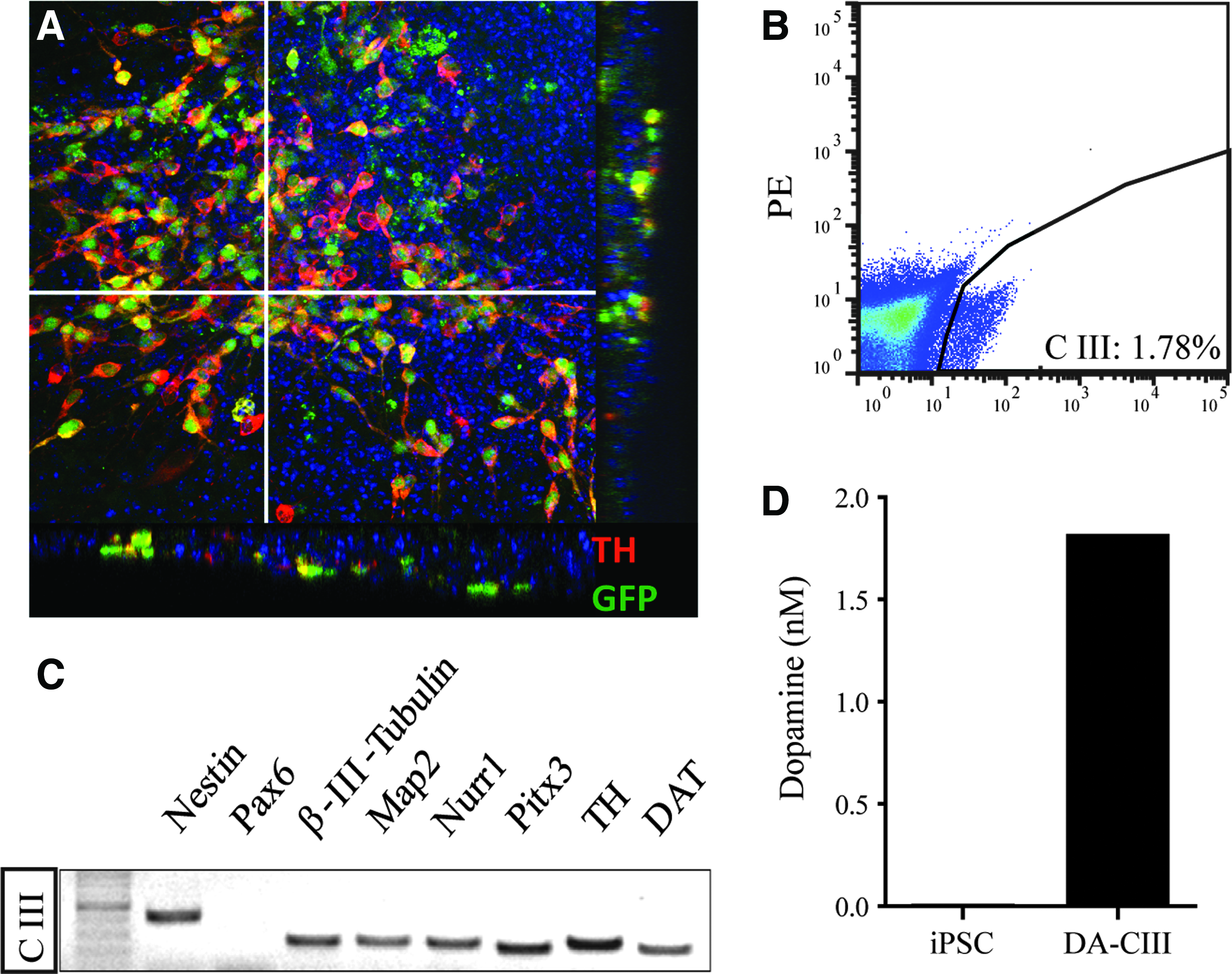
Dopaminergic differentiation of 3 Pitx3GFP/+ iPSC clones. (
Overexpression of L1CAM and PSA-NCAM in iPSC-derived DA neurons
The sorted Pitx3-GFP+ DA neurons were transduced with a lentiviral vector containing the genes encoding human L1CAM or STX; an empty vector was used as a control. The transduced cells were washed thoroughly before they were plated onto primary astrocyte layers. The expression of L1CAM and STX was analyzed using qPCR. There was endogenous expression of mouse L1CAM and mouse STX in DA neurons transduced with control vectors; the level of endogenous L1CAM and STX expression in astrocytes appeared to be very low (Fig. 2A, C). One week after transduction, specific expression of human L1CAM mRNA could be detected at a four times higher level in the L1CAM vector-transduced group (Fig. 2A). L1CAM was also detected by immunofluorescent staining both in the soma and along the neurite of DA neurons in both groups (Fig. 2B). The expression of human STX was about 10 times higher in the STX vector-transduced group in comparison to the control vector group (Fig. 2C). Immunostaining showed the presence of PSA-NCAM both in the soma and the neurite of DA neurons in both groups (Fig. 2D). Both L1CAM and PSA-NCAM were not detectable in the primary astrocyte layer (Fig. 2C, D).
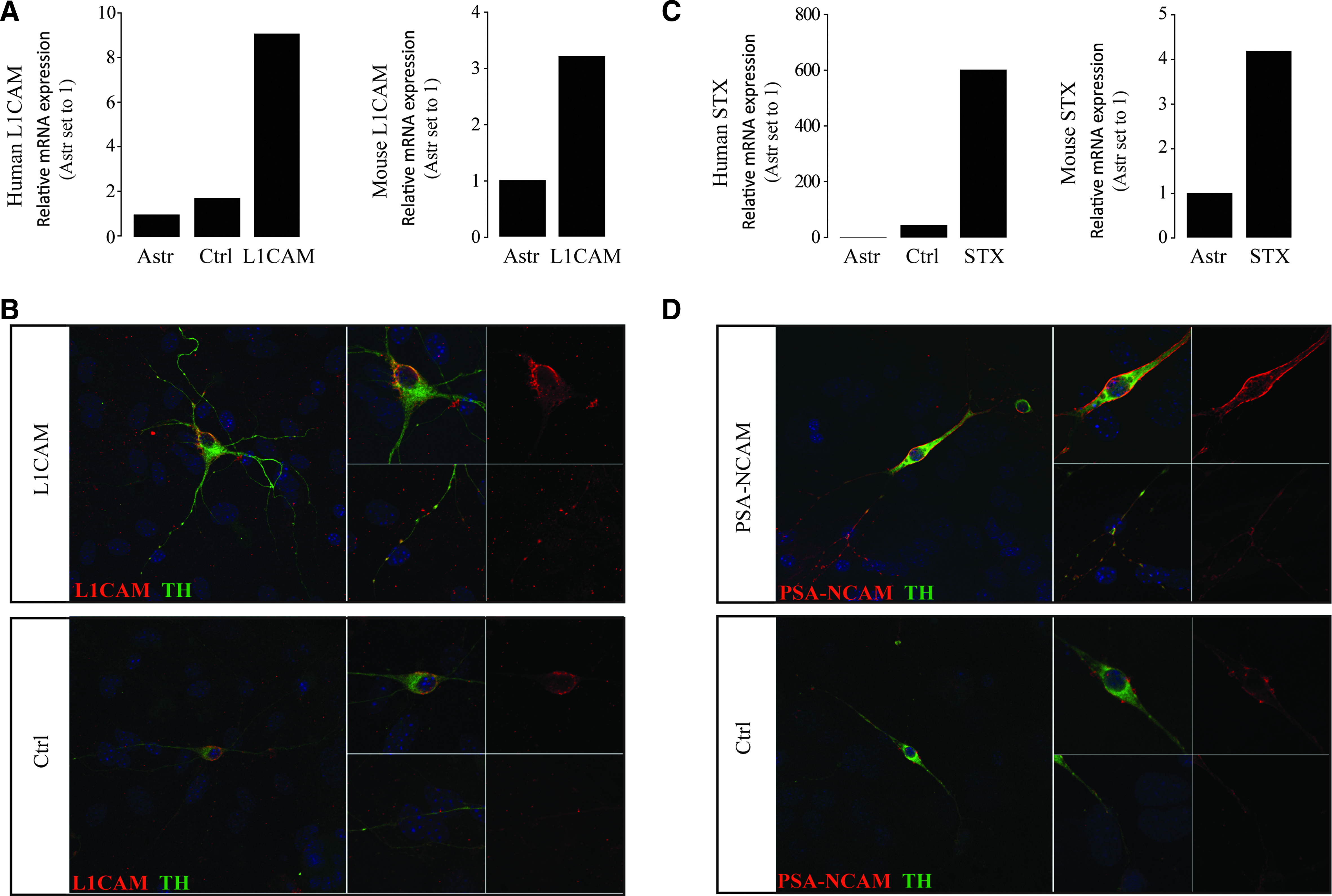
Overexpression of hL1CAM and hSTX in iPSC-derived DA neurons. (
L1CAM and PSA-NCAM overexpression stimulates neurite outgrowth of iPSC-derived DA neurons in vitro
Transduced DA neurons were plated onto a primary astrocyte layer to create an optimal supporting environment for outgrowing neurons, enabling a clear analysis of the effects of L1CAM and PSA-NCAM overexpression on cell morphology. DA neurons co-cultured on top of a primary astrocyte layer had an oval or trigonal soma of approximately similar sizes (Fig. 3A). Tracings made of DA neurons from the control group and the L1CAM overexpressing group revealed bipolar or tripolar neurite formation, whereas DA neurons in the PSA-NCAM group occasionally also had five or six primary neurites (Fig. 3B). However, this difference was not significant among the three groups (control vs. L1CAM, 0.79; control vs. PSA-NCAM, 0.17). The morphology of the neurites in both L1CAM and PSA-NCAM groups was much more complex (Fig. 3A), with more branches than in the control group (Fig. 3C). The longest neurite from each TH+ DA neuron was also measured. Both L1CAM and PSA-NCAM overexpressing cells extended their neurite significantly longer than the control ones (Fig. 3D). With longer neurites and more branches, the total neurite length in these two groups was significantly longer (Fig. 3E) and covered a larger area (Fig. 3F).
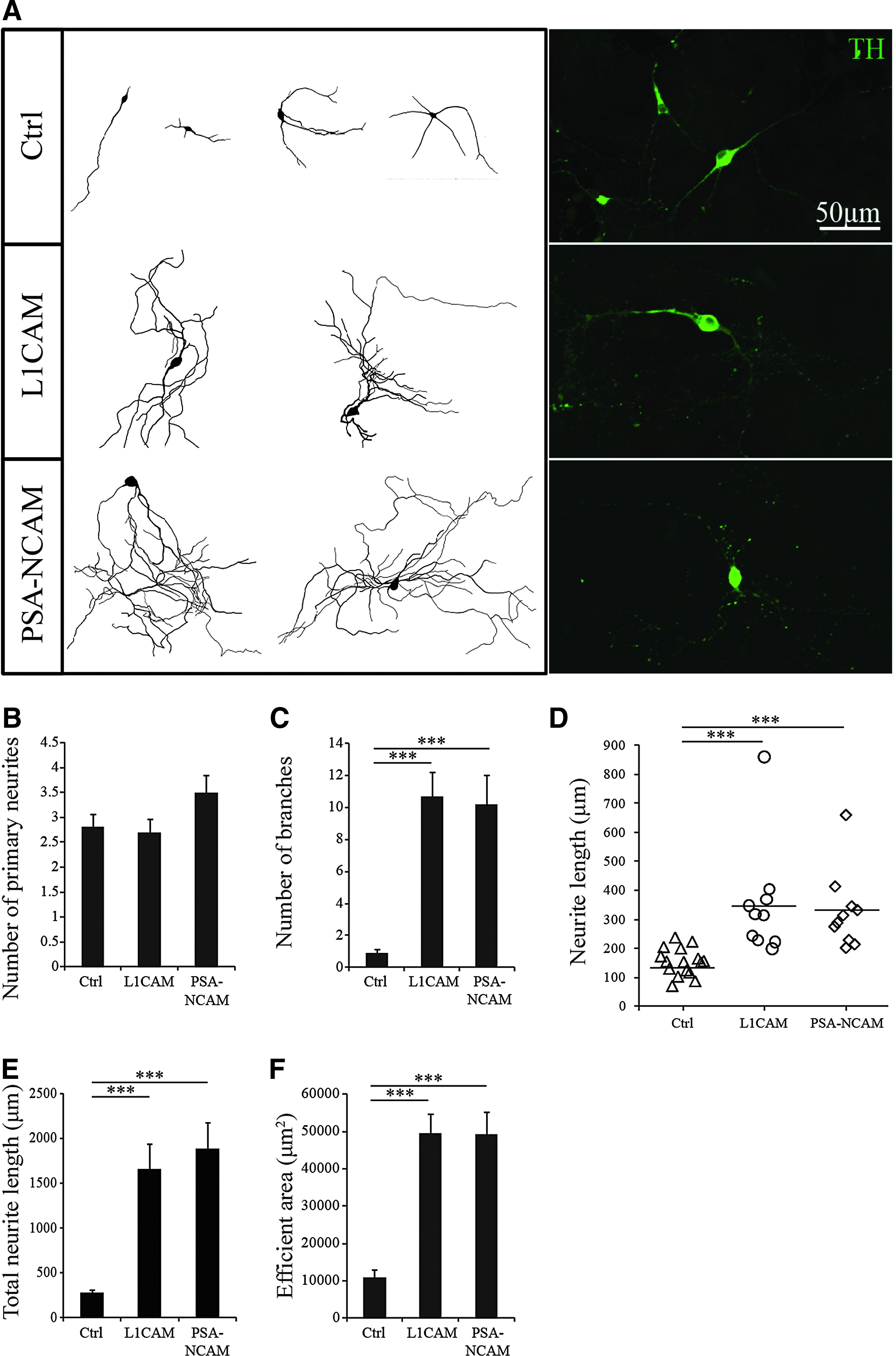
Overexpression of L1CAM and STX stimulated the outgrowth of iPSC-derived DA neurons on an astrocyte layer. (
Effect of L1CAM and PSA-NCAM on neurite outgrowth of iPSC-derived DA neurons in organotypic slices
To provide an environment mimicking the striatal in vivo situation, transduced iPSC-derived DA neurons from all three groups (L1CAM or PSA-NCAM overexpressing and controls) were placed onto mouse organotypic striatal slices and cultured for 1 week. TH+ DA neurons were either clustered as groups or scattered on the slice (Fig. 4A). Analysis of the number of primary neurites revealed no significant differences between the three groups (control vs. L1CAM, 0.48; control vs. PSA-NCAM, 0.43) (Fig. 4C). In general, all DA neurons on the striatal slices grew much longer neurites than those cultured on the astrocyte layers (Fig. 4B vs. Fig. 3D). This may be due to the high level of cell adhesion molecules such as L1CAM and PSA-NCAM already present in postnatal organotypic striatal slices (Fig. 5), and the maximum neurite outgrowth of scattered DA neurons was promoted.
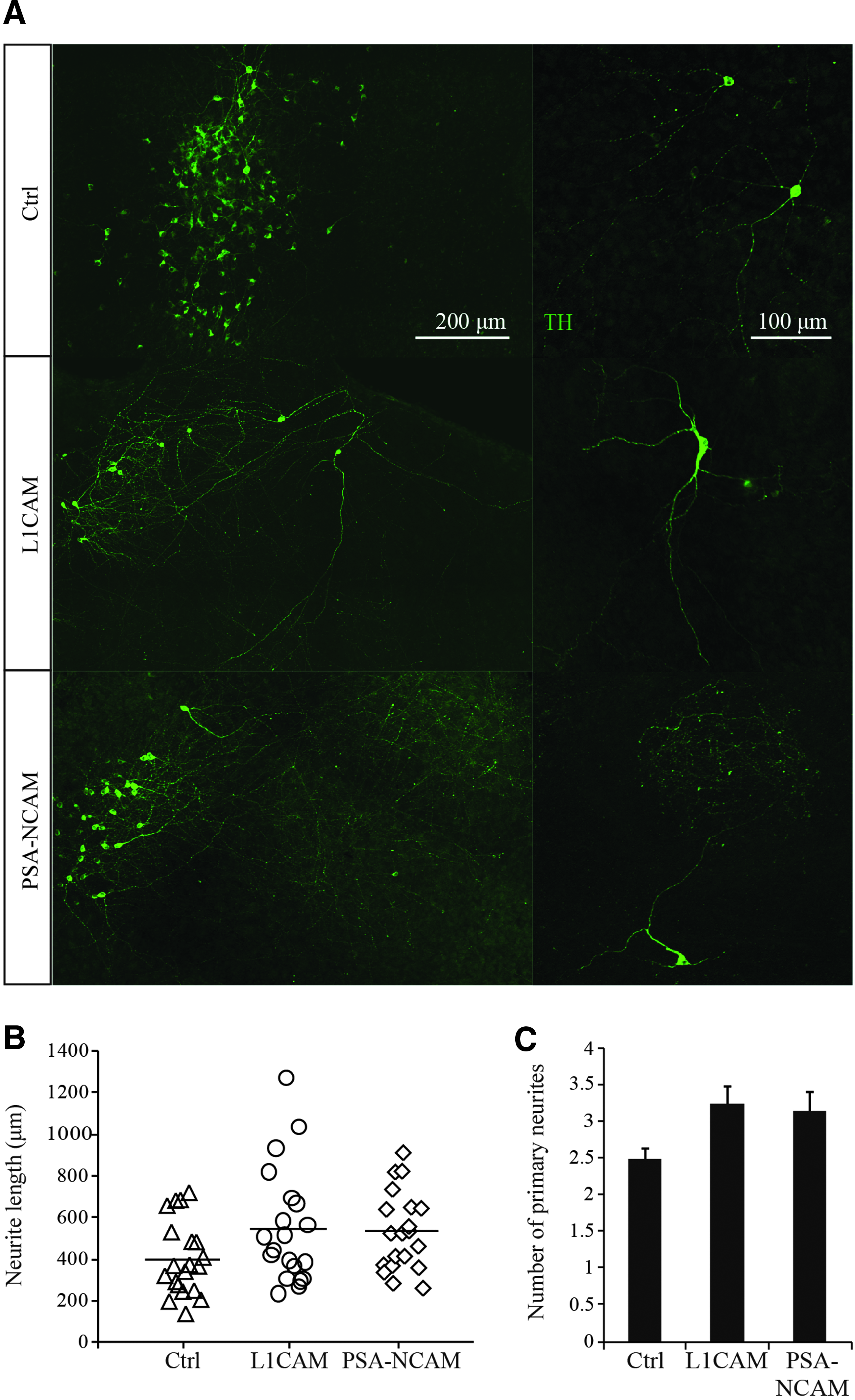
Outgrowth of iPSC-derived DA neurons on organotypic striatal slices. (
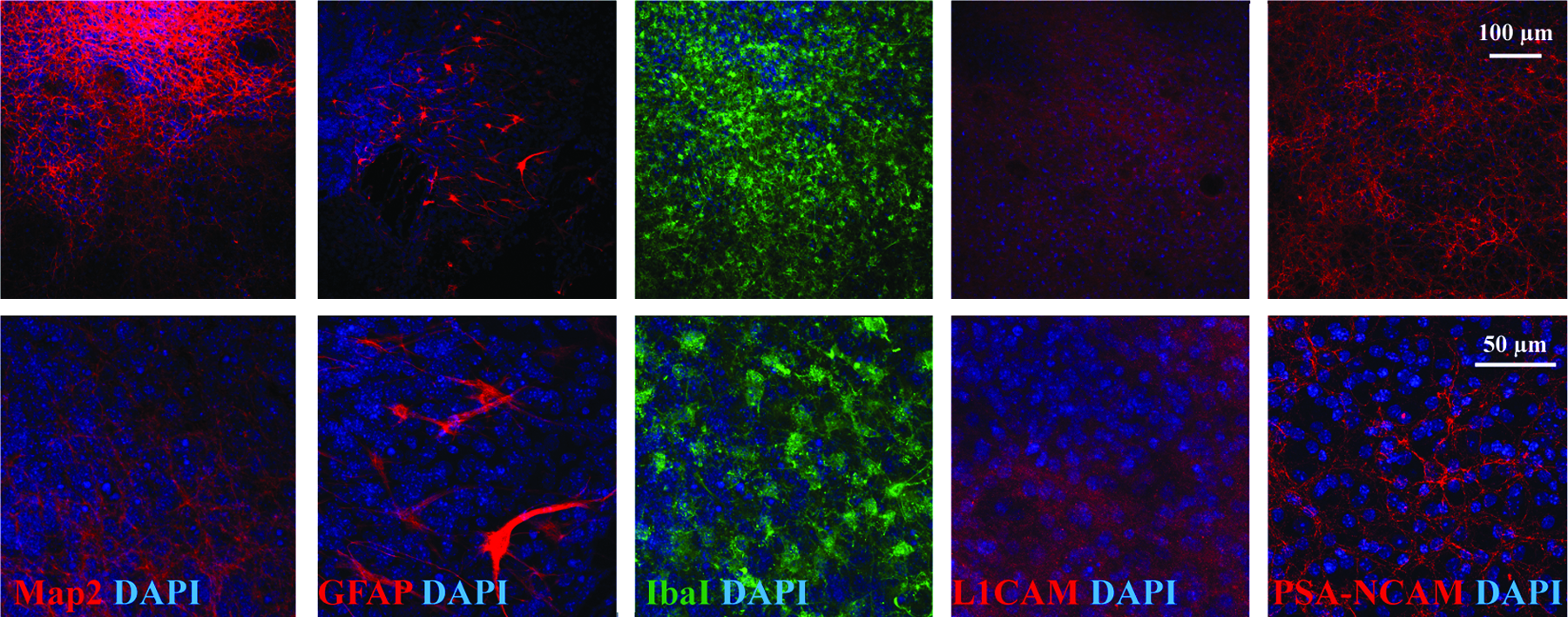
Organotypic striatal slice culture. Striatal slices were cut from the striatum of P1–P3 mice with a thickness of 250 μm. The slices were cultured for 2 weeks before sorted DA neurons were layered on them. Striatal slices were characterized by immunostaining for the neuronal marker β-III-tubulin (
Due to the neuronal clustering and the severe intertwining of the neurites, it was difficult to trace all branches from one neuron. Therefore, we only traced the longest neurite distinguishable from each neuron (mostly scattered DA neurons). On average, the longest neurite from each neuron in the L1CAM and PSA-NCAM group is longer than the control group; however, no significant differences were observed (control vs. L1CAM, 0.45; control vs. PSA-NCAM, 0.31) (Fig. 4B). Remarkably, clustered DA neurons of the L1CAM and PSA-NCAM groups tended to extend their neurites in a parallel direction (Fig. 4A), unlike those of the control group (Fig. 4A). Moreover, the neurites in L1CAM and PSA-NCAM group were much longer than in control group. This phenomenon may point to the fact that L1CAM- and PSA-NCAM–overexpressing DA neurons are more responsive to patterning cues in the striatal slice.
Implantation of iPSC-derived DA neurons in 6-OHDA lesion rats
Sorted control iPSC-derived DA neurons and iPSC-derived DA neurons overexpressing L1CAM or PSA-NCAM were transplanted into unilaterally 6-OHDA lesioned rats, according to the procedure described before (Roessler et al., 2014). Two weeks after 6-OHDA injection, rats that rotated more than 5 rpm after amphetamine injection were included in the experiments. At 8 weeks after implantation, a small reduction in the number of rotations was observed in the groups (Fig. 6A). Immunohistochemical examination of the implanted striata revealed the survival of only, on average, 423 (± 108) implanted TH+ neurons in the denervated striatum. The presence of only a very small number of DA neurons may account for the small reduction in rotation behavior.
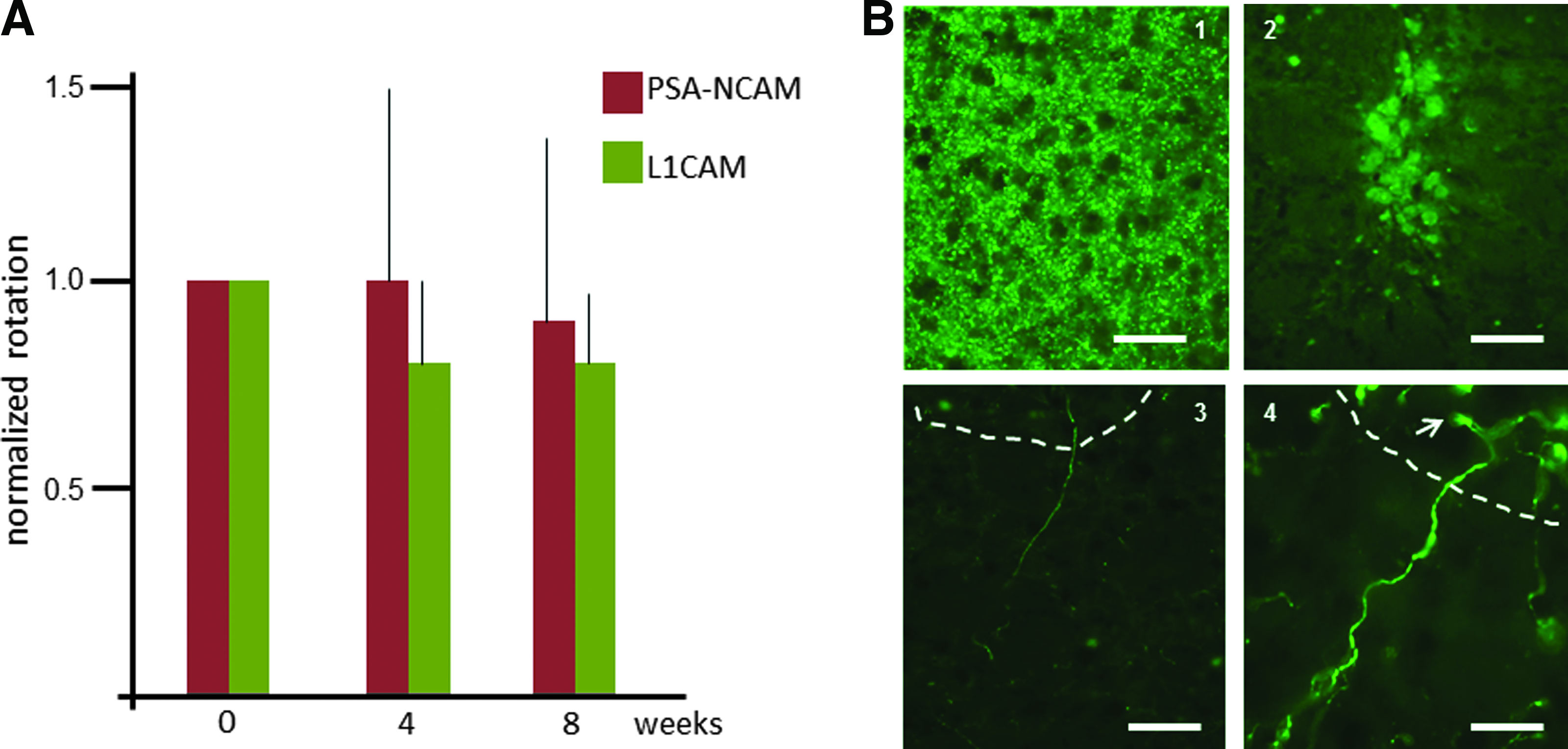
Intrastriatal transplantation of iPSC-derived DA neurons in unilaterally 6-OHDA–lesioned rats. (
Apparently, the subsequent stressful procedures of FACS and viral transduction had made the DA neurons vulnerable for the stress associated with the implantation procedure, resulting in the survival of only about 2% of the injected cells. Accurate analysis of the three-dimensional outgrowth of the newly formed neurites extending from the surviving individual implanted neurons showed that the maximal length of the neurites in each of the groups of implanted iPSC-derived DA neurons did not exceed 500 μm (Fig. 6B). As observed in the cultures on the striatal organotypic slices, no significant differences were found between the neurite outgrowth of the control and the L1CAM- or PSA-NCAM–expressing grafted DA neurons, respectively, 468 ± 108, 492 ± 64, and 477 ± 73 μm.
Discussion
Considering the option of a cell replacement therapy for PD, iPSCs are presently the most promising source for an unlimited number of autologous DA neurons to be grafted intracerebrally. The in vitro–differentiated, iPSC-derived DA neurons have been demonstrated to adopt the characteristics of true (VM) DA neurons as assessed by cell type–specific marker expression, electrophysiology, and dopamine release (Hargus et al., 2010; Kriks et al., 2011). However, comparison of the genome-wide expression profile of iPSC-derived DA neurons to primary DA neurons showed a subset of hypermethylated genes that were downregulated in iPSC-derived VM DA neurons. Gene annotation analysis showed that these genes were involved in nervous system development, neuron differentiation, neurogenesis, and neurite outgrowth (Roessler et al., 2014). This may explain the fact that iPSC-derived DA neurons are not as efficient as primary DA neurons in reinnervating the DA depleted striatum in PD 6-OHDA rats and in giving rise to functional recovery.
Extensive striatum-wide reinnervation is an absolute prerequisite for a DA neuron replacement therapy for PD to be successful. Yet the short neurites extending from grafted iPSC-derived DA neurons appeared to be restricted within a short range around the injection track (Hargus et al., 2010; Wernig et al., 2008). In the present study, we attempted to restore this hampered neurite outgrowth potential of iPSC-derived DA neurons by forcing their overexpression of L1CAM and PSA-NCAM, both crucial adhesion molecules for axon guidance during embryonic neuronal development. It should be noted that our comparative genome-wide expression profiling did not reveal differences in the expression of genes encoding the formation of L1CAM and PSA-NCAM between the primary and the iPSC-derived DA neurons. iPSCs were reprogrammed from fibroblasts of Pitx3GFP/+ mice (Maxwell et al., 2005) and subsequently differentiated into DA neurons. Inducing overexpression of L1CAM and PSA-NCAM in DA neurons derived from iPSCs appeared to be cumbersome.
Our first approach was to transduce the relevant genes in the iPSCs and to establish stably transduced iPSC lines that were subsequently differentiated into DA neurons. However, L1CAM expression was lost during differentiation and could only be detected in the undifferentiated cells. This is in accordance with previous observations on transgene silencing when transduced stem cells differentiate into specialized cell types (Hamaguchi et al., 2000; Vroemen et al., 2005). We decided to perform lentiviral hL1CAM and hSTX gene transduction on a suspension of Pitx3-GFP DA neurons sorted by FACS and were able to show stable upregulation of both hL1CAM and hSTX in the transduced iPSC-derived DA neurons.
These L1CAM and PSA-NCAM overexpressing iPSC-derived DA neurons allowed us to evaluate the effect of these molecules on neurite outgrowth in vitro on top of a feeder layer of astrocytes (which did not express these molecules). We demonstrated that, indeed, the neurites in both THE L1CAM and PSA-NCAM groups showed a much more complex, branched morphology and overall length than those in the control group. When the iPSC-derived DA neurons were cultured on top of organotypic slices of postnatal striatum, similar differences were observed, but these differences were less pronounced and not statistically significant. Because the postnatal striatal slices appeared to be already rich in L1CAM and PSA-NCAM content, we speculate that this may have obscured the effects of growth promotion of iPSC-derived DA neurons by the transduced adhesion factors. Only the pattern of outgrowth of the L1CAM or PSA-NCAM overexpressing iPSC-derived DA neurons appeared different from the control ones, indicating a different response of these cells to the cues in the substrate.
We intended to examine the effect of L1CAM and PSA-NCAM on the neurite outgrowth of iPSC-derived DA neurons in the denervated striatum of 6-OHDA– lesioned PD rats. In our previous studies (Roessler et al., 2014), we already had demonstrated that iPSC-derived DA neurons transplanted in this rat model do survive and show some outgrowth. The subsequent stressful procedures of FACS and lentiviral transduction made the DA neurons vulnerable to the stress associated with the implantation procedure, resulting in the survival of only about 2% of the injected cells. Low survival (<3%) of implanted iPSC-derived DA neurons have been described before (Doi et al., 2014; Rhee et al., 2011), and sorting before transduction has been shown to further reduce the survival rate to less than 0.06% (Hargus et al., 2010).
Although our in vitro experiments clearly revealed the outgrowth-stimulating ability of L1CAM and PSA-NCAM, pronounced outgrowth stimulation could not be detected in the few surviving L1CAM and PSA-NCAM overexpressing iPSC-derived DA neurons in the denervated striatum of the unilaterally 6-OHDA–lesioned rats. This is in contrast to recent findings by Battista et al. (2014), who demonstrated that PSA-NCAM overexpression was able to enhance neurite outgrowth of stably transduced mESC-derived DA neurons in vivo. Moreover, experimental studies with the spinal cord injury model clearly demonstrated that L1CAM (Chen et al., 2007) and PSA-NCAM (Zhang et al., 2007) were able to enhance axon regeneration and extension. It is possible that the sorting, lentiviral transduction of the sorted DA neurons, and implantation protocol affected the viability and health of the surviving DA neurons, interfering with the potential growth-promoting effects of the adhesion factors.
In conclusion, we have demonstrated that L1CAM and PSA-NCAM significantly stimulate the neurite outgrowth of iPSC-derived DA neurons, both neurite extension and branching in vitro. However, we failed to demonstrate significant neurite outgrowth stimulation by L1CAM and PSA-NCAM in iPSC-derived DA neurons co-cultured on striatal organotypic slices and after implantation in the 6-OHDA-denervated striatum. Apparently, under these conditions, induced expression of L1CAM- or PSA-NCAM in the iPSC-derived DA neurons cannot completely restore the neurite outgrowth potential that was reduced in these DA neurons as a consequence of epigenetic aberrations resulting from the iPSC-reprogramming process.
Footnotes
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.