
Abstract
Abstract
Increased intracellular cyclic adenosine monophosphate (cAMP) can promote axonal elongation and facilitate neuronal repair, while decreased cAMP is associated with losses in neuronal regenerative capacity. Rolipram, which upregulates intracellular cAMP by blocking phosphodiesterase‐4 (PDE4) enzyme activity, can mitigate diverse neurological disorders. In this study, we investigated whether rolipram induces neuronal differentiation of human bone marrow-mesenchymal stem cells (hBM-MSCs). Rolipram-treated MSCs (Roli-MSCs) had significantly increased expression of the neuroprogenitor proteins Nestin, Musashi, GFAP, and Sox-2. When Roli-MSCs were differentiated with neuronal induction media (Roli-dMSCs), they exhibited cell body and dendritic morphologies similar to those of neurons. The neurite number and length of Roli-dMSCs were significantly increased compared to those of differentiated MSCs (dMSCs). Compared with undifferentiated hBM-MSCs, the Roli-dMSCs and dMSCs showed significantly increased expression of the neuronal-specific marker genes Nestin, Musashi, CD133, GFAP, NF-M, MAP-2, KCNH1, KCNH5, SCN3A, and CACNA1A, and decreased expression of other lineage-specific markers Adiponectin, ALP, FABP4, and MMP13. The Roli-dMSCs also showed a higher expression of the neuronal markers Nestin, Musashi, Sox-2, NF-M, and Tuj-1 compared to those of the undifferentiated hBM-MSCs, measured by immunocytochemistry and immunoblotting assay. Thus, we have shown that rolipram ameliorates neuronal differentiation by the regulation of neuroprogenitor expression in hBM-MSCs, and rolipram treatment of MSCs may improve the therapeutic efficacy of stem cell therapy for neurodegenerative disorders.
Introduction
H
Recent studies have shown that an increased level of intracellular cyclic adenosine monophosphate (cAMP) can promote axonal regeneration (Blesch et al., 2012; Neumann and Woolf, 1999) and facilitate neuronal repair (Neumann et al., 2002; Qiu et al., 2002), while a decreased level of intracellular cAMP is associated with losses in neuronal regenerative capacity (Cai et al., 2001). It has been reported that cAMP promotes neuronal differentiation by upregulating the expression levels of the neuronal markers NSE, Tuj1, and GFAP (Zhang et al., 2011).
A cAMP‐hydrolyzing enzyme, phosphodiesterase‐4 (PDE4), which is a key regulator of the levels of intracellular cAMP, hydrolyzes cAMP to 5′-AMP, resulting in a decrease in the intracellular cAMP level. PDE4 inhibition has been shown to improve depression, cognitive faculty (O'Donnell and Zhang, 2004), and Alzheimer's disease (Garcia-Osta et al., 2012), while PDE4B-deficient animals presented an antipsychotic phenotype (Siuciak et al., 2007).
A selective PDE4 inhibitor, rolipram, has been developed as a potential antidepressant drug (Krause et al., 1990). It has been widely examined in several disease models, including autoimmune diseases (Kumar et al., 2013), Alzheimer's disease (Garcia-Osta et al., 2012; Gong et al., 2004), cognitive enhancement (Normann and Berger, 2008), Huntington's disease (DeMarch et al., 2007), traumatic brain injury (Atkins et al., 2007), spinal cord injury (Hannila and Filbin, 2008), and respiratory diseases (Huang and Mancini, 2006), and has been identified to have beneficial effects in these disease models. Interestingly, recent studies have shown that resveratrol, which ameliorates aging-related metabolic phenotypes and neuronal degeneration, also inhibits PDE4, leading to an increased level of cAMP (Park et al., 2012).
Since modulation of cAMP has been associated with axonal elongation and neuronal regeneration (Blesch et al., 2012; Neumann and Woolf, 1999; Neumann et al., 2002; Qiu et al., 2002), and has shown beneficial effects in neurological diseases (Garcia-Osta et al., 2012; O'Donnell and Zhang, 2004; Siuciak et al., 2007), we investigated whether the PDE4 inhibitor rolipram can promote neuronal differentiation in hBM-MSCs.
Materials and Methods
Characteristics of primary hBM-MSCs and cell culture
The hBM-MSCs were purchased from CEFO (Cell Engineering for Origin, Seoul, Korea) and cultured as described in the previous study (Joe et al., 2015). Cells at passage 7 were used for all the experiments.
Quantitative polymerase chain reaction (real time polymerase chain reaction)
Total RNAs were extracted from MSCs, rolipram-MSCs, differentiated MSCs (dMSCs) and rolipram-dMSCs using RNAiso reagent (TaKaRa Bio, Shiga, Japan). cDNAs were synthesized using the Primescript™ II 1st strand cDNA synthesis kit (TaKaRa Bio) according to the manufacturer's instructions, and then amplified using Power SYBR® Green PCR master mix (Applied Biosystems, Inc., Waltham, MA) with gene-specific primers for human Nestin, Musashi, CD133, GFAP, BDNF, Syn1, MAP-2, NF-M, KCNH1, KCNH5, SCN3A, CACNA1A, Nanog, Oct-4, ALP, Adiponectin, FABP4, MMP13, or β-actin (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/cell). Used primer pairs were synthesized by GenoTech Corp. (Daejeon, South Korea) and IDT (Integrated DNA Technologies, Inc., Coralville, IA).
(4,5-Dimethylthiazol-2-yl-)2,5-diphenyltetrazolium bromide assay
The cell viabilities of control-MSCs and rolipram-MSCs were measured by the (4,5-dimethylthiazol-2-yl-)2,5-diphenyltetrazolium bromide (MTT) assay (Sigma-Aldrich, St. Louis, MO). Briefly, 2.5 × 103 hBM-MSCs were cultured for a day onto 96-well plates. The cells were then treated with 0–25 μM of rolipram for 12 hours and tested by the MTT assay.
Neuronal differentiation
Control-MSCs and rolipram-treated MSCs were differentiated into neuron-like cells using the neuronal differentiation method, which were established in the previous study (Jeong et al., 2013; Joe et al., 2015; Woodbury et al., 2000). Images were captured under Eclipse TS100 microscope (Nikon, Tokyo, Japan) and i-Solution IMTcam3 digital camera (JENOPTIK, Jena, Germany). Cells were considered differentiated neurons when each cell body had more than two dendrites or axon longer than 60 μm. Neurite length was measured using the ImageJ program (NIH, Bethesda, MD).
Immunocytochemistry
Control- and rolipram-treated MSCs were cultured on poly-
Immunoblotting
Total proteins were extracted from MSCs using 400 μL of protease and phosphatase inhibitors containing the RIPA buffer (Santa Cruz Biotechnology), and the immunoblot analysis was performed using the antibodies against NF-M (1:500), Nestin (1:500), Tuj-1 (1:1000), Musashi (1:500), GFAP (1:500), Sox-2 (1:500), or β-actin (1:5000), and subsequently, the appropriate horseradish peroxidase-conjugated secondary antibodies (1:10,000; Jackson ImmunoResearch Laboratories, West Grove, PA). Band densities were quantified with the ImageJ program (NIH).
Results
Neuroprogenitor gene markers are expressed by rolipram in hBM-MSCs
To evaluate the dose effects of rolipram, hBM-MSCs were incubated with 0, 0.5, 1, 5, or 25 μM of rolipram for 12 hours. The expression of the neuroprogenitor markers Nestin and CD133 was assessed by quantitative polymerase chain reaction (qPCR). As shown in Figure 1A, the expression of these genes was increased by rolipram treatment in a dose-dependent manner at doses ≤1 μM and was decreased at higher doses. (Fig. 1A; t-test, *p < 0.05, **p < 0.005, mean ± SD, n = 4). To determine the optimal time of rolipram treatment in hBM-MSCs, MSCs were incubated with 1 μM rolipram for several different periods of time (0–24 hours). Expression of the tested genes peaked after incubation for 12 hours (Fig. 1B; t-test, *p < 0.05, **p < 0.005, mean ± SD, n = 3). Cellular toxicity was not observed at doses of ≤5 μM rolipram for 12 hours (Fig. 1C; t-test, *p < 0.05, mean ± SD, n = 3). From these data, we determined that treatment with 1 μM rolipram for 12 hours was the optimal condition.
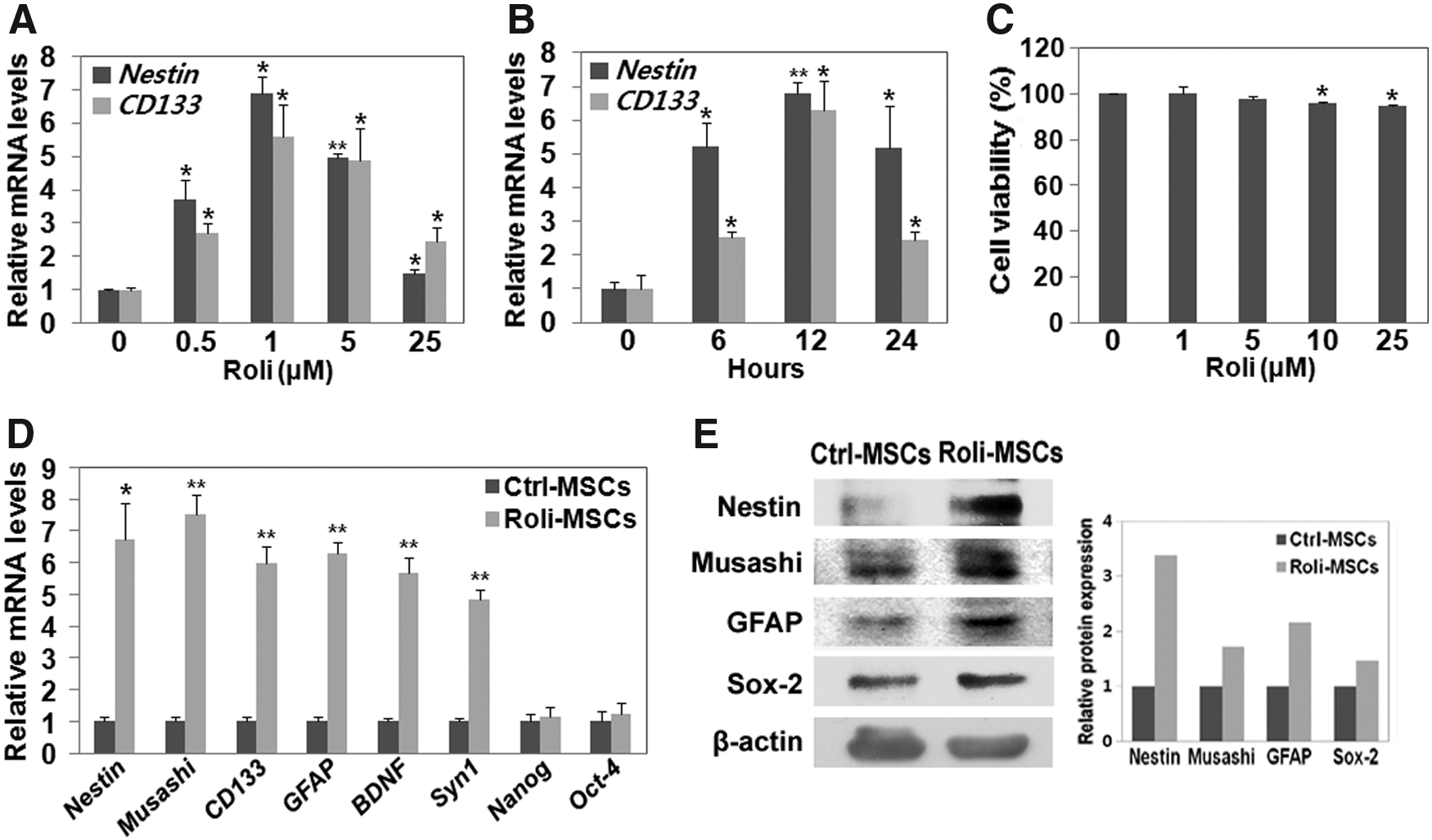
Rolipram induces the expression of neuroprogenitor markers in hBM-MSCs.
To further examine the role of rolipram in regulating cell fate, additional neuroprogenitor markers were tested by qPCR under the optimal condition (1 μM rolipram for 12 hours). The expression of the neuroprogenitor genes Nestin, Musashi, CD133, GFAP, BDNF, and Syn1 was significantly increased in rolipram-treated hBM-MSCs (Roli-MSCs) compared with that of the nontreated hBM-MSCs (Fig. 1D; t-test, *p < 0.05, **p < 0.005, mean ± SD, n = 3). Similarly, the increased protein levels of Nestin, Musashi, GFAP, and Sox-2 were confirmed by immunoblot analysis (Fig. 1E).
Rolipram-MSCs have a neuronal appearance in neuronal induction media
After we had identified the induction of the neuronal progenitor genes by rolipram treatment in MSCs, we hypothesized that rolipram-treated MSCs could acquire better differentiation potency into neuronal cells than control MSCs (Ctrl-MSCs) could do. To examine this, both Ctrl-MSCs and Roli-MSCs were differentiated with neuronal induction media (Jeong et al., 2013; Joe et al., 2015; Woodbury et al., 2000). Both neuronally differentiated MSCs (dMSCs and Roli-dMSCs) exhibited neuronal appearance, while undifferentiated BM-MSCs (Ctrl-MSCs) and Roli-MSCs showed a typical appearance similar to that of primary hBM-MSCs (Fig. 2A). Roli-dMSCs showed a significantly higher estimated rate of differentiation into neuron-like cells than dMSCs did (Fig. 2B and Supplementary Table S2; t-test, *p < 0.05, mean ± SD, n = 4). The average neurite length of dMSCs was also significantly greater in Roli-dMSCs than in dMSCs (Fig. 2C, D, and Supplementary Table S3; t-test, *p < 0.05, **p < 0.005, mean ± SD, n = 4), and a significant difference in the number of neurites was observed for dMSCs versus Roli-dMSCs (Fig. 2D; t-test, *p < 0.05, mean ± SD, n = 4).
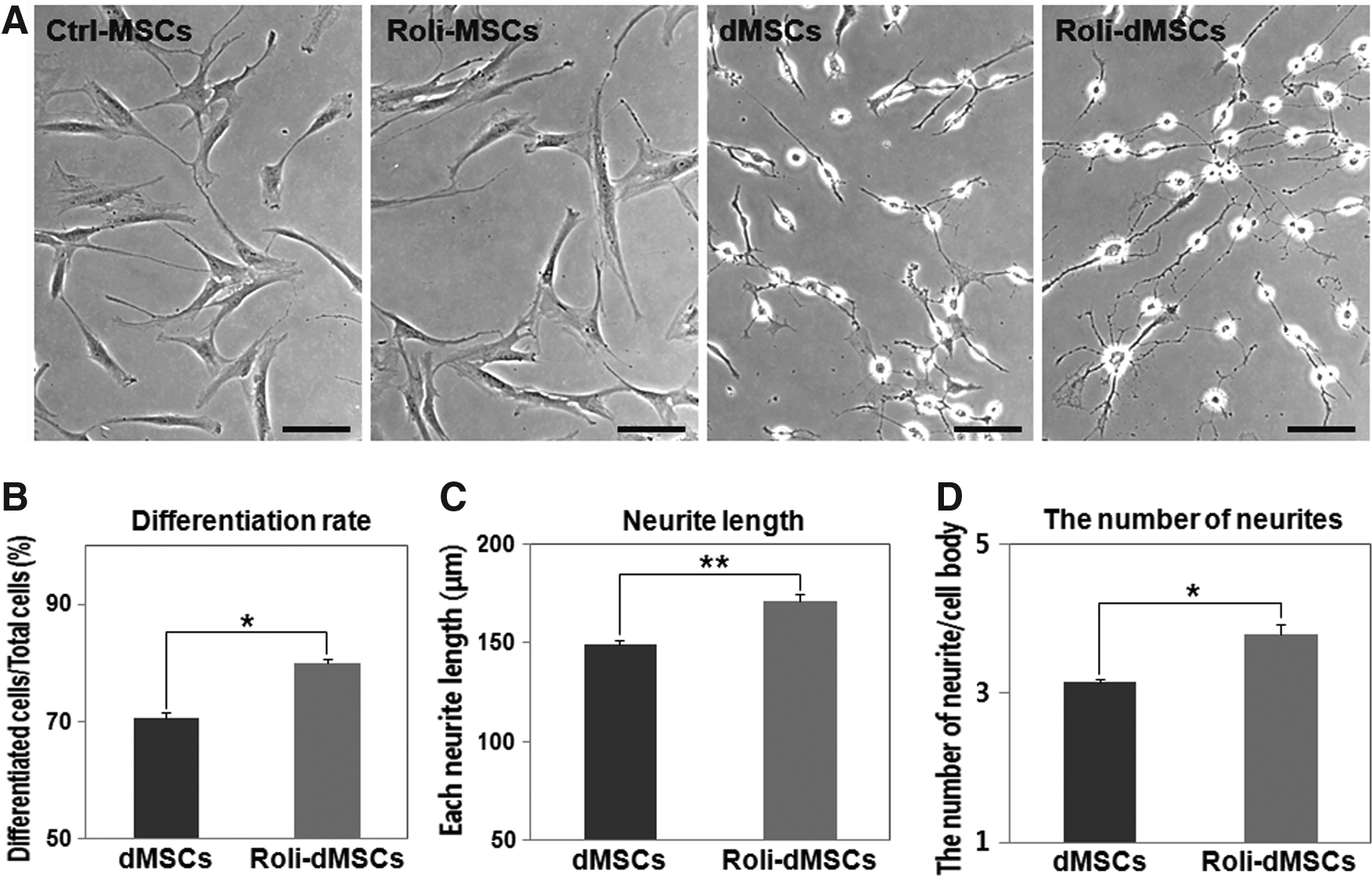
Neuron-like morphological changes in rolipram-pretreated hBM-MSCs followed by neuronal induction.
Increased transcription of various neuronal markers in dMSCs treated with rolipram
Since Roli-dMSCs have a neuronal appearance, the expression of several neuronal-specific genes was examined by real-time PCR (Fig. 3A). The expression levels of all the neuronal marker genes examined in this study were significantly increased in both Roli-dMSCs and dMSCs compared with those of the undifferentiated MSCs (Ctrl-MSCs) (Fig. 3A; t-test, *p < 0.05, **p < 0.005, mean ± SD, n = 5). The expression of neuronal-specific marker genes (CD133, NF-M, MAP-2, KCNH5, SCN3A, and CACNA1A) was significantly increased in Roli-dMSCs compared with that of dMSCs (Fig. 3A; t-test, *p < 0.05, **p < 0.005, mean ± SD, n = 5). We also tested the other lineage-specific genes' expression in dMSCs and Roli-dMSCs, and we identified decreased expression levels of adipogenic (Adiponectin and FABP4)-, chondrogenic (MMP13)-, and osteogenic (ALP)-specific genes in dMSCs and Roli-dMSCs compared with those of the Ctrl-MSCs (Fig. 3B; t-test, **p < 0.005, mean ± SD, n = 4).
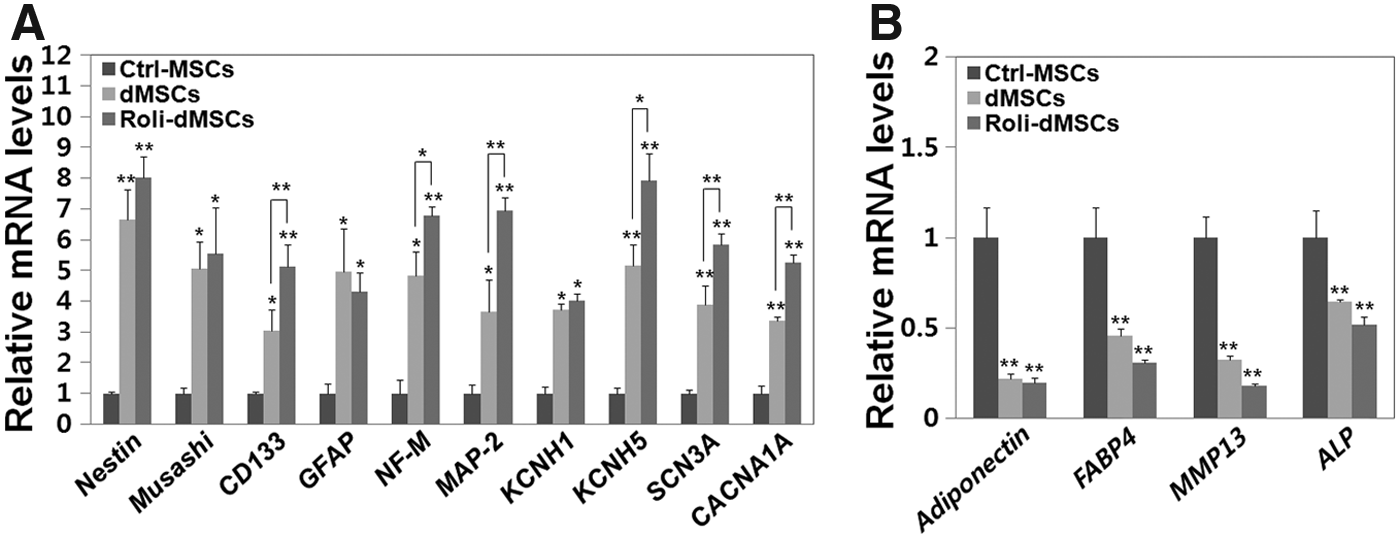
Increased expression of diverse neuronal markers in differentiated rolipram-treated MSCs.
Neuronal-specific proteins are upregulated in rolipram-pretreated dMSCs
To characterize whether the expression levels of markers of neuronal differentiation were changed at the protein level, we used an immunocytochemistry assay. Ctrl-MSCs, dMSCs, and Roli-dMSCs were stained with the neuronal markers'-specific antibodies for Nestin, NF-M, and Tuj-1. Neuronal-induced MSCs (dMSCs) and Roli-dMSCs exhibited positive cytoplasmic staining for Nestin, NF-M, and Tuj-1, while the noninduced MSCs were negative for these markers (Ctrl–MSCs; Fig. 4A). Similar results were obtained by immunoblot analysis using antibodies specific to Nestin, Musashi, Sox-2, NF-M, and Tuj-1 (Fig. 4B).
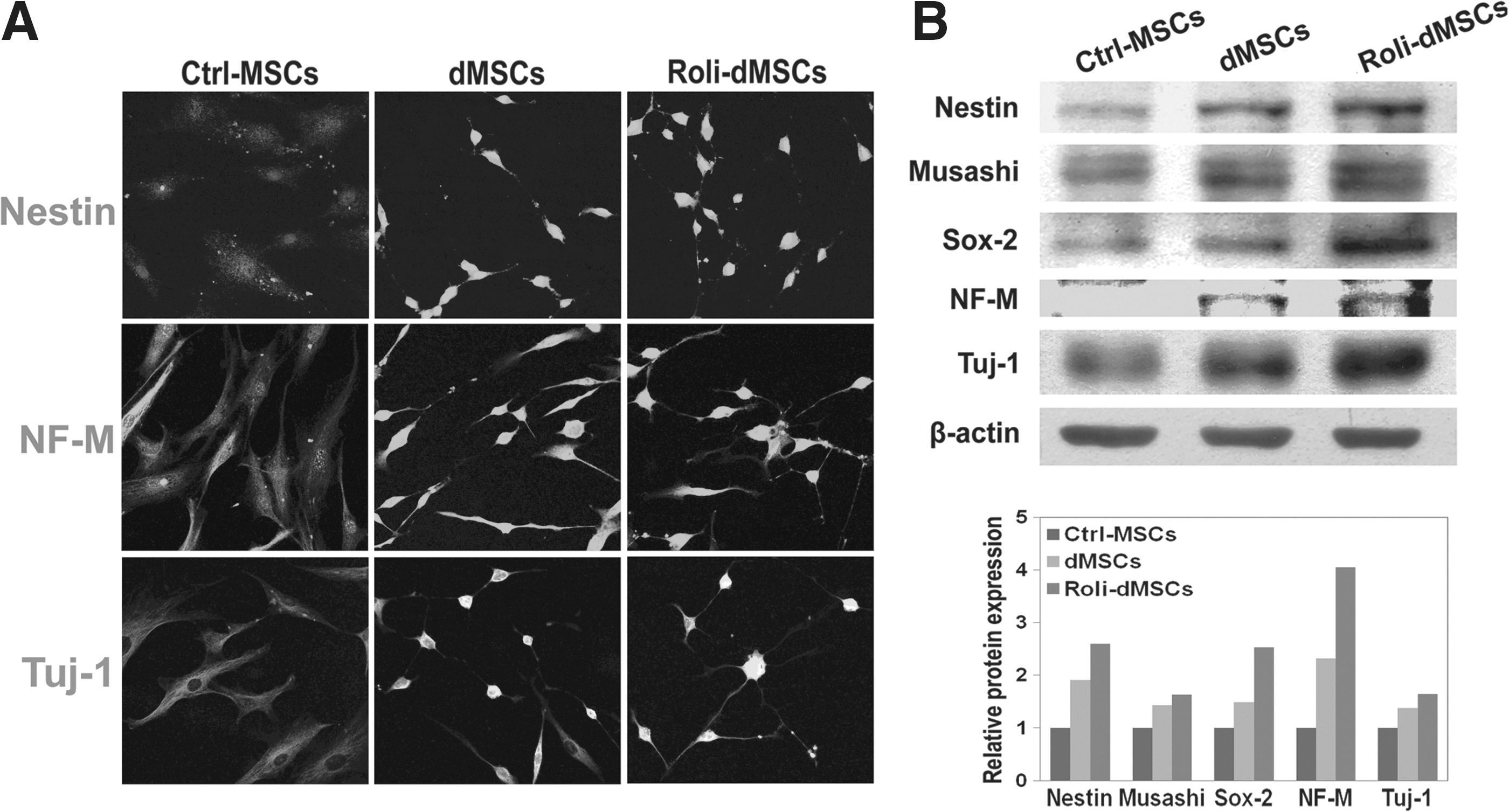
Upregulation of neuronal-specific proteins in rolipram-dMSCs.
Discussion
Increasing cAMP involves diverse cellular physiological processes, including neuronal differentiation (Deng et al., 2001; Zhang et al., 2011). Failure of the cellular mechanisms that normally regulate intracellular cAMP homeostasis contributes to diverse neurological diseases (Garcia-Osta et al., 2012; O'Donnell and Zhang, 2004; Siuciak et al., 2007). Inhibition of the cAMP-hydrolyzing enzyme PDE4 improves these diseases (Atkins et al., 2007; DeMarch et al., 2007; Garcia-Osta et al., 2012; Gong et al., 2004; Hannila and Filbin, 2008; Huang and Mancini, 2006; Kumar et al., 2013; Normann and Berger, 2008), suggesting that PDE4 may be an important therapeutic target for these neurological diseases. In this study, we investigated whether the PDE4-selective inhibitor, rolipram, could facilitate neuronal differentiation in hBM-MSCs. Rolipram-treated MSCs showed an increased expression of neuronal progenitor genes under optimized treatment conditions (1 μM for 12 hours; Fig. 1). When cultured with neuronal induction media (Jeong et al., 2013; Joe et al., 2015; Woodbury et al., 2000), Roli-dMSCs showed significant increases in the expression of neuronal-specific markers and the rate of neuronal differentiation, compared with those of dMSCs (Figs. 2–4).
Increasing the levels of intracellular cAMP has been shown to sustain neuronal architecture by stimulating axon elongation and neurite outgrowth (Blesch et al., 2012; Neumann and Woolf, 1999). When using rolipram as a PDE4 inhibitor in this study, both neurite lengths and the number of neurites were significantly increased in Roli-dMSCs. The expression levels of the dendritic-specific gene MAP-2 and the axonal marker Tuj-1 (Moskowitz and Oblinger, 1995) were both elevated in Roli-pretreated dMSCs compared with those of dMSCs (Figs. 3 and 4). This suggests that Roli-MSCs had greater neurite outgrowth and a higher rate of differentiation into neuron-like cells than did the Ctrl-MSCs in the presence of neuronal induction media.
We have shown that rolipram treatment lead to neuronal differentiation in hBM-MSCs. However, the signaling pathways associated with this process have not been examined. Rolipram treatment increases intracellular cAMP by inhibiting the PDE4 enzyme activity. The elevated cAMP may activate protein kinase A (PKA) and cAMP-responsive element-binding protein (CREB), subsequently leading to the increased expression of several genes, potentially including genes associated with neuronal induction (Gong et al., 2004). Previous studies have shown that PDE4 inhibition improves symptoms in neurodegenerative disease models (Atkins et al., 2007; DeMarch et al., 2007; Gong et al., 2004; Hannila and Filbin, 2008; Normann and Berger, 2008) and enhances the differentiation capacity of stem cells (Deng et al., 2001; Kim et al., 2005; Zhang et al., 2011), which supports our results.
hBM-MSCs, which have the capacity to differentiate into neurons (Francischi et al., 2000; Jeong et al., 2013; Joe et al., 2015; Woodbury et al., 2000), have been used for cell therapy in neurodegenerative diseases (Dantuma et al., 2010). However, only a few cells were successfully differentiated into neurons when this approach was attempted in an in vivo animal study (Fan et al., 2013), suggesting that the differentiation potency of donor stem cells should be improved before cell therapy. In this study, we have shown that rolipram-treated MSCs endogenously induced the expression of neuroprogenitor proteins that simultaneously facilitated the differentiation into neuron-like cells in the presence of neuronal induction media. Thus, rolipram-treated MSCs may enhance neuronal differentiation and thereby improve the efficacy of stem cell therapy for neurodegenerative diseases.
Footnotes
Acknowledgment
This work was supported by research fund from Chosun University, 2015.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.