
Abstract
Abstract
Culture conditions have been established to maintain the pluripotency of mouse naïve and primed embryonic stem cells (ESCs) using human amnion epithelial cells (hAECs) as the feeder layer. In this study, the ultrastructures of mouse primed ESCs grown on hAECs were analyzed by transmission electron microscopy. Consistent with mouse naïve ESCs, the undifferentiated mouse primed ESC line ESD-EpiSC [ESC-derived epiblast stem cells (EpiSCs)] revealed typical characteristics, including large nuclei, reticulated nucleoli, scanty cytoplasm, and low cytoplasm-to-nuclear ratios. Cells had prominent Golgi apparatus and well-developed endoplasmic reticulum. Adjacent cells were tightly in contact with dense junction desmosomes. However, in EpiSC colonies, cell contact was no longer close like naïve ESCs, and differentiated cells existed. The differentiated cells had small nucleoli with large cytoplasm, which represented primitive mesenchyme. Phagosomes or apoptotic cells were also common in the cytoplasm of differentiated cells, which suggests a differentiation potential. When exposed to leukemia inhibitory factor (LIF), ESD-EpiSCs could convert to naïve-like cells. We further analyzed the ultrastructure of converted EpiSCs (cESCs). As compared to ESD-EpiSCs, cESCs showed similar ultrastructural characteristics as naïve ESCs. These findings suggest that ultrastructure could be used to evaluate the pluripotency of ESCs.
Introduction
M
Zhang et al. (2010) first demonstrated that EpiSCs could be achieved from embryoid bodies (EBs) derived from naïve mouse ESCs in the presence of activin A/bFGF. ESC-derived epiblast stem cell (ESD-EpiSC) is an immortal cell line derived from naïve mESCs with the same characteristics as EpiSCs from developing epiblast. Previously, we had reported that ESD-EpiSC could be maintained and propagated on human amnion epithelial cells (hAECs). ESD-EpiSCs could also be reprogrammed to naïve-like pluripotent cells with LIF signaling (Chen et al., 2013).
The characteristics of mouse primed ESCs are well established (Brons et al., 2007; Tesar et al., 2007; Ying et al., 2003). However, their ultrastructure has not yet been reported. In this study, we identified the ultrastructure of mouse primed ESCs and compared it with naïve mouse ESCs. We found ultrastructural differences in the primed ESCs that revealed its pluripotency potential.
Materials and Methods
Cell culture
hAECs were prepared as previously reported (Chen et al., 2013). Human amnion membranes were obtained with written informed consent from pregnant women, and their appropriate use was approved by the institutional ethics committee of the International Peace Maternity and Child Health Hospital. The membranes were cut into pieces and digested by 0.25% trypsin for 30–40 min at 37°C. The resultant cell suspensions were cultured in RPMI-1640 medium supplemented with 10% fetal calf serum (FCS; PAA, Austria), streptomycin (100 U/mL), penicillin (100 U/mL), and glutamine (0.3 mg/mL) and incubated in a humidified tissue culture incubator containing 5% CO2 at 37°C. hAECs at a density of 80–90% were used as feeder layers for mouse ESC culture.
ESD-EpiSCs (Zhang et al., 2010) were maintained on fresh prepared hAECs in Dulbecco's Modified Eagle Medium (DMEM)/F12 medium (Invitrogen) supplemented with 2 mM glutamine (Invitrogen), 0.1 mM minimum essential medium (MEM) and nonessential amino acids (NEAA; Invitrogen), 0.1 mM β-mercaptoethanol (Sigma), and 20% knockout serum replacement (Invitrogen). About 12 ng/mL bFGF (Invitrogen) or 1 × 103 U/mL LIF (Chemicon) was added when converted EpiSCs were cultured.
Naïve mESCs (R1) were maintained as above, besides 1 × 103 U/mL LIF (Chemicon) was added. The medium was changed every day.
Immunofluorescence staining
The mouse ESCs cultured on hAEC feeder layers were fixed with 4% paraformaldehyde for 15–20 min at room temperature. Oct4 was assessed with rabbit monoclonal antibodies (1:400; Santa Cruz); Nanog was detected with rabbit polyclonal antibodies (1:200; Chemicon) and then visualized with fluorescein isothiocyanate (FITC)-conjugated goat anti-rabbit immunoglobulin G (IgG; 1:200, Santa Cruz). Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI). Fluorescence images were visualized and recorded with an inverted fluorescence microscope (Leica, DMI3000).
Transmission electron microscopy
Mouse naïve ESCs, ESD-EpiSCs, and converted EpiSCs colonies grown on hAEC feeder layers were picked separately and first fixed with 2.5% glutaraldehyde in phosphate-buffered saline (PBS; 0.1 M, pH 7.4) for 24 h at 4°C. The secondary fixation was performed with 1% osmium tetroxide in the same buffer for 1 h at room temperature. Samples were then dehydrated and embedded in epoxy resin. Ultrathin sections were counterstained with lead citrate and uranyl acetate for 10 min using standard technique. Ultrastructure visualization was performed using an energy-filtered transmission electron microscope (TEM) (Tecnai G2 Spirit TEM, FEI Company, USA).
Results
Both mouse naïve cells and primed ESCs could be maintained on human amnion epithelial cells
Previously, we reported that hAECs could maintain the pluripotency of mouse naïve ESCs and primed ESCs ESD-EpiSCs (Chen et al., 2013). Here, we showed that undifferentiated growth of ESD-EpiSCs could be maintained with bFGF using hAECs as the feeder layer (Fig. 1A). We noticed some differentiated cells along the border of colonies, which is common in ESD-EpiSC cultures (Fig. 1B). Upon exposure to LIF, ESD-EpiSCs underwent a reprogramming process on hAECs and became naïve-like converted ESCs (cESCs) (Fig. 1C).
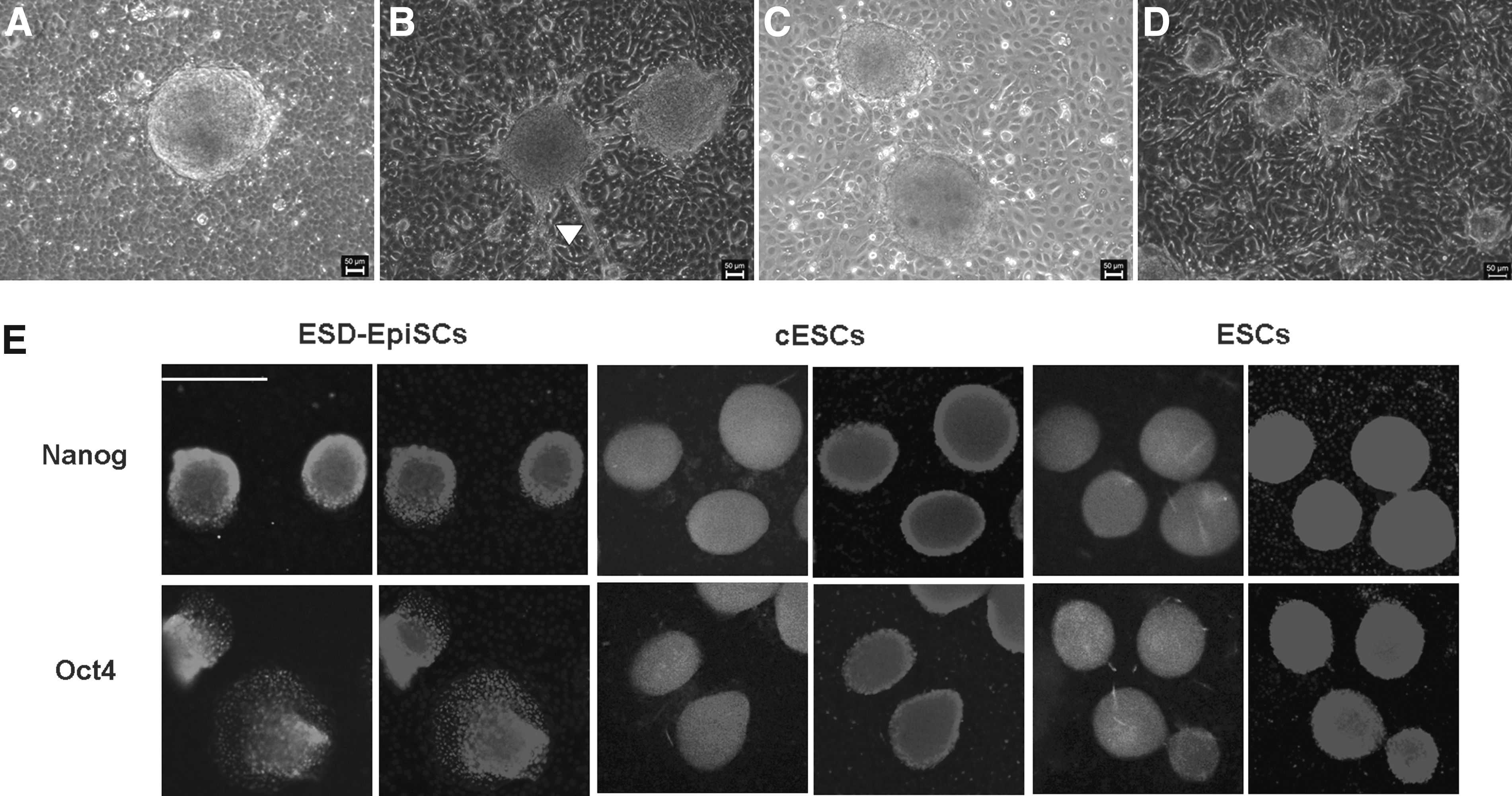
The EDS-EpiSC colonies were maintained on hAECs with either bFGF or LIF. (
From the shape of the colonies, both primed and naïve ESCs appeared dome-like and compact while growing from the center toward the periphery (Fig. 1). The pluripotent stem cell markers, including Nanog and Oct4, were clearly detected by immunostaining in ESD-EpiSCs, cESCs, and naïve ESCs on hAECs (Fig. 1E).
Distinct ultrastructural characteristics of mouse primed and naïve ESCs
The mouse naïve ESCs had typical ultrastructural characteristics, including large nuclei, reticulated nucleoli, and scanty cytoplasm (Fig. 2). Likewise, most ESD-EpiSCs (85% of colonies) had scanty cytoplasm with large nuclei indicative of protein-synthesizing cells (Fig. 3A). However, differentiated cells with small nucleoli and large cytoplasm representing primitive mesenchyme were commonly found (15% of colonies). Phagosomes containing cell debris or apoptotic cells were also observed in the cytoplasm of differentiated cells (Fig. 3B, C). Compared to tightly contacted mouse naïve ESCs, the gaps between differentiated cells were obviously increased, whereas dense cell junctions were evident in the ESD-EpiSCs colony (Fig. 3D).
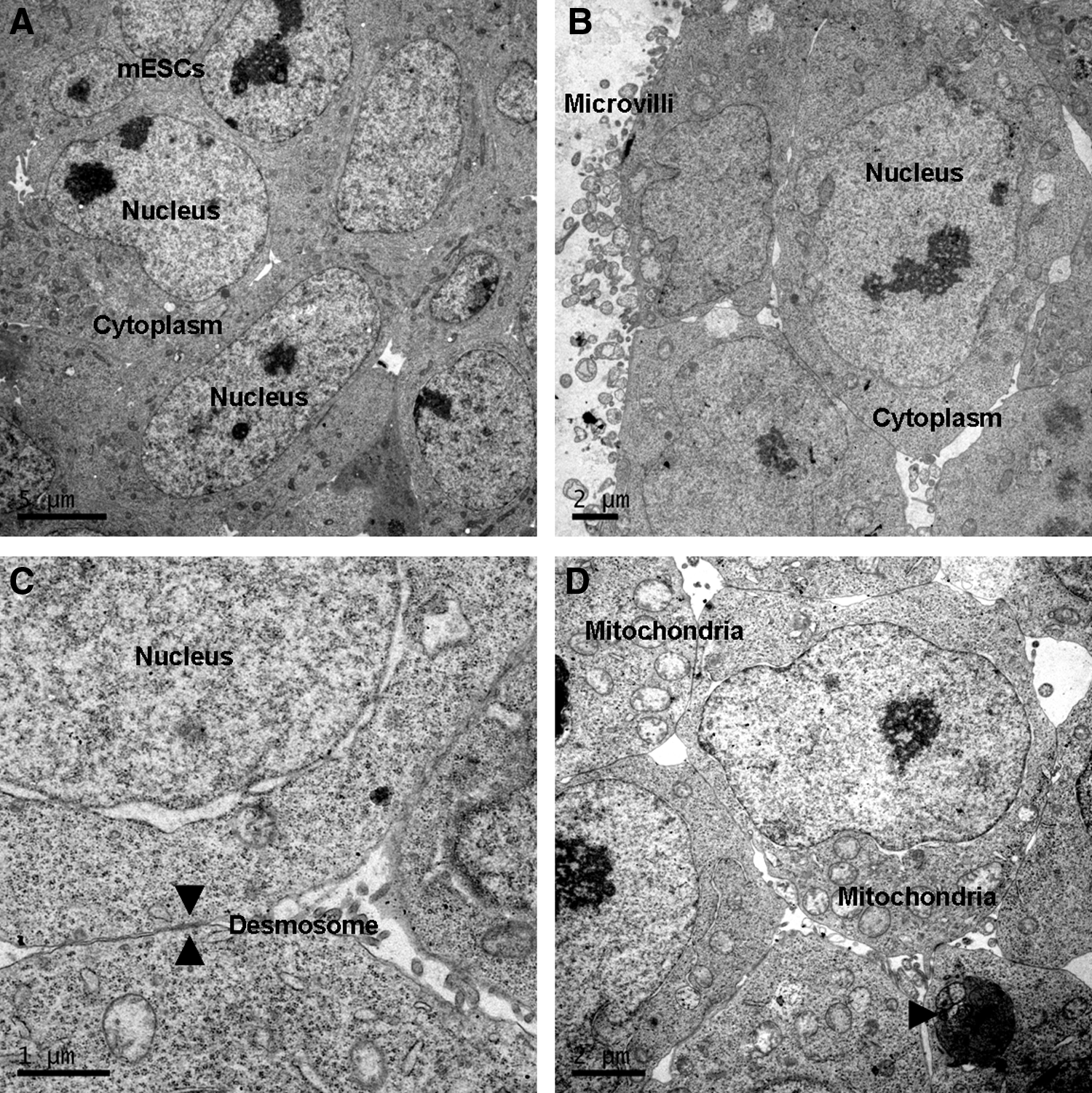
The ultrastructure of mouse ESCs. (
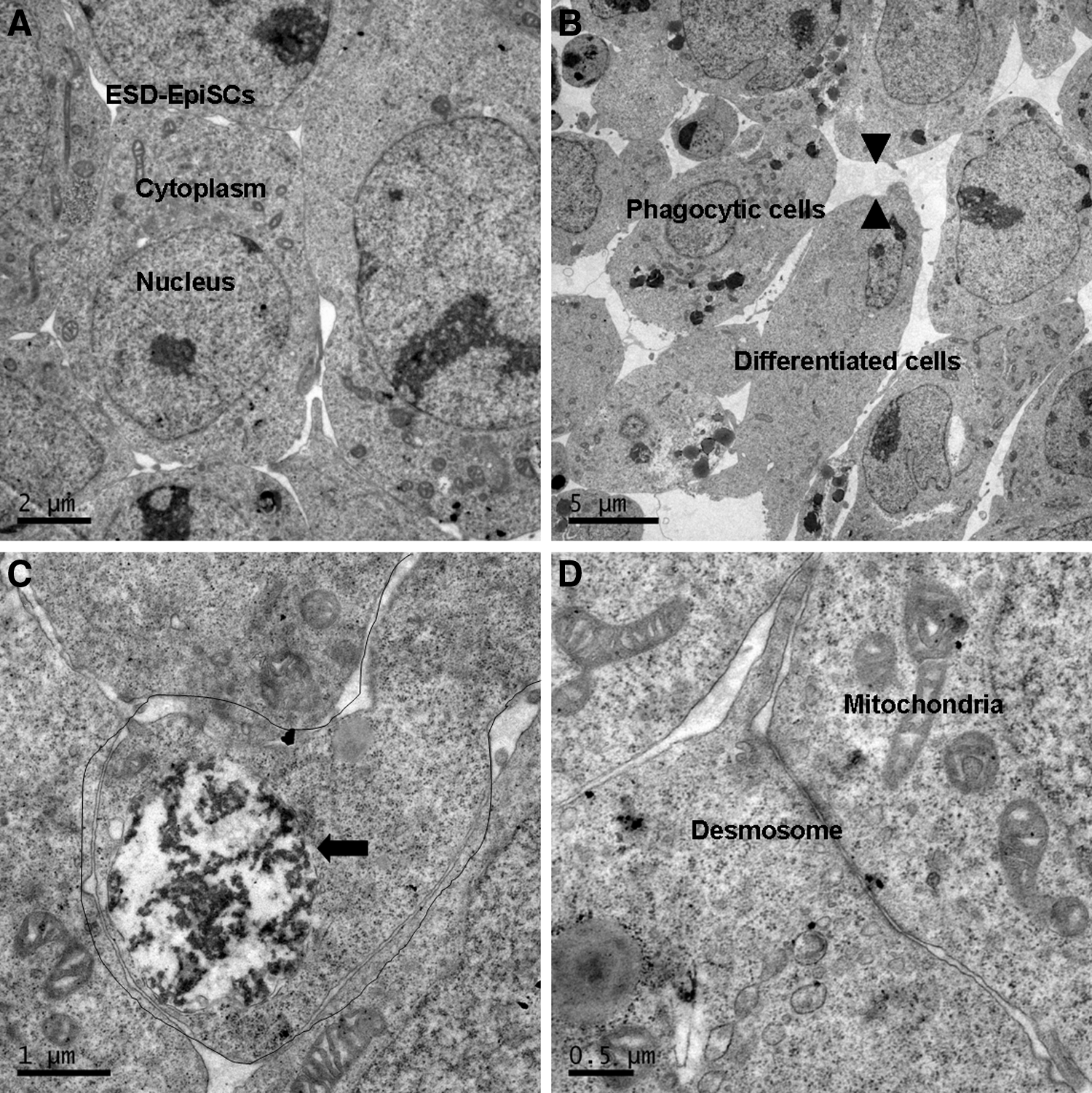
As compared to mouse ESCs, differentiated cells were common in ESD-EpiSC colonies. (
Previously, we reported that ESD-EpiSCs could undergo a reprogramming process when exposed to LIF and were converted to naïve-like cells in which naïve pluripotency markers were activated and primed markers were suppressed (Chen et al., 2013). Herein, we further analyzed the ultrastructure of converted ESCs (cESCs). As compared to ESD-EpiSCs, cESCs showed similar ultrastructural characteristics as naïve ESCs (Fig. 4) and had scanty cytoplasm with large nuclei (98% of colonies). Cells were close to each other, leaving less extracellular space between adjacent cells and contact with desmosome. The Golgi apparatus and rough endoplasmic reticulum were both clear and well developed, showing typical morphology. Mitochondria were organized at the periphery of the nucleus. However, less differentiated cells were detected in cESC colonies (2% of colonies) as compared to ESD-EpiSCs (15% of colonies; 2% of colonies versus 15% of colonies, p < 0.01).
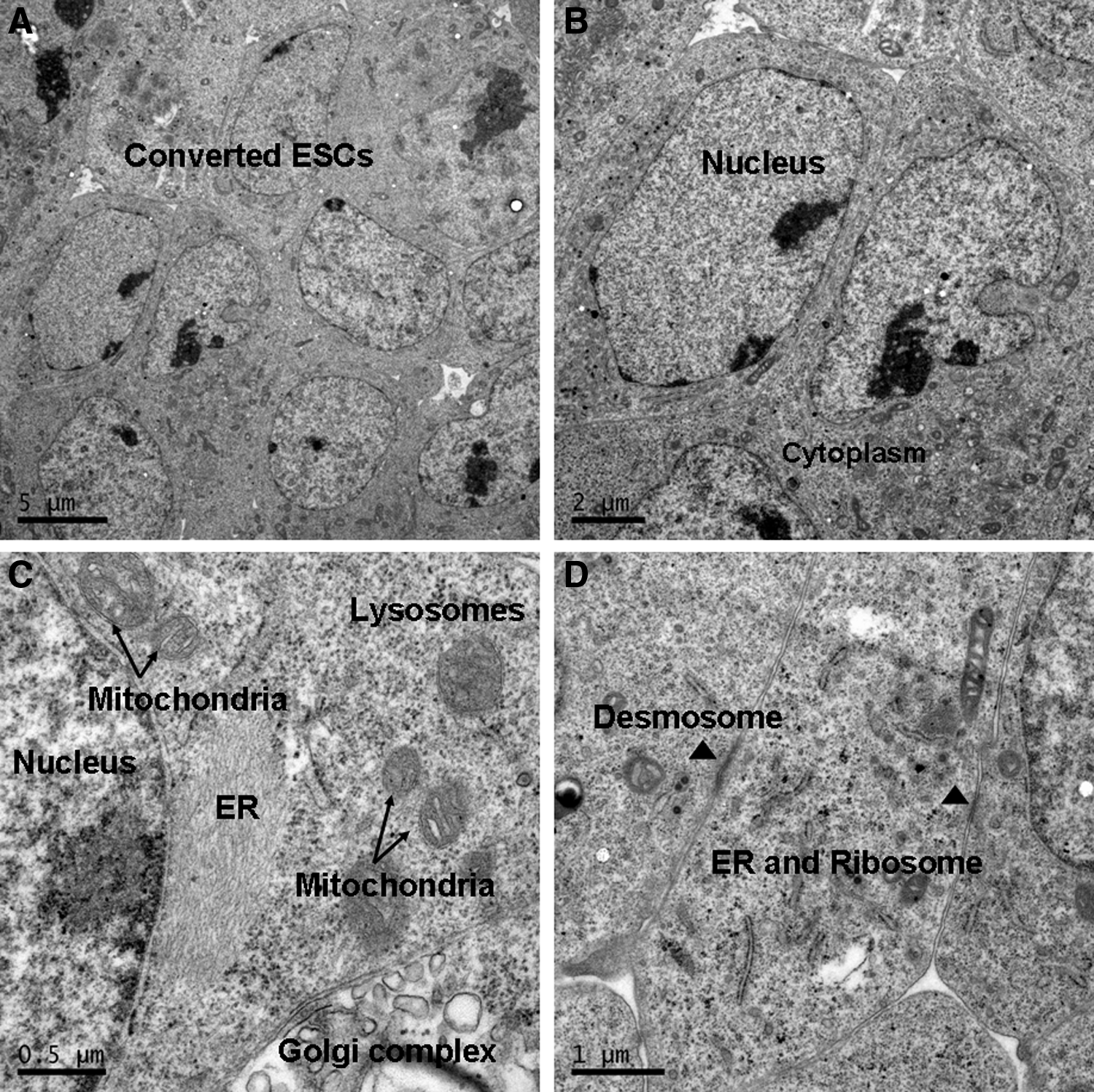
ESD-EpiSCs could revert to ESC-like cells based on LIF signaling. (
Discussion
The ultrastructure of mouse ESCs has been well documented. However, the present study is the first to demonstrate the ultrastructure of mouse primed ESCs. As compared to previously reported mouse ESCs (Baharvand and Matthaei, 2003), our naïve mouse ESC colonies consisted mostly of undifferentiated cells with large nuclei, reticulated nucleoli, and scanty cytoplasm showing a high nuclear-to-cytoplasmic ratio. The cells were tightly contacted with desmosomes. Cell structure included microvilli, mitochondria, lysosomes, Golgi complex, and well-developed rough endoplasmic reticulum (ER). However, ESD-EpiSCs consisted of both undifferentiated and differentiated cells. Differentiated cells and apoptotic cells were commonly seen in mouse primed ESCs.
Human ESCs (hESCs) are believed to lack the potential to be naïve and pluripotent and resemble mouse primed EpiSCsin morphology, developmental potency, gene expression, and epigenetic modification (Buecker et al., 2010; Hanna et al., 2009, 2010). Sathananthan et al. (2002) reported that the ultrastructure of human ESCs consisted of undifferentiated cells resembling inner cell mass (ICM) of blastocysts and differentiated cells, including epithelioid and isolated cells. Herein, we found that differentiated cells were common in the ESD-EpiSCs colony, which was similar to the ultrastructure of human ESCs. Elongated cells representing primitive mesenchyme containing phagocytic and apoptotic cells were observed in ESD-EpiSC colonies. Han et al. demonstrated two subpopulations in EpiSCs, Oct4-GFP positive and negative EpiSCs, which might lead to the restricted pluripotency of EpiSCs. Oct4-GFP positive EpiSCs, which only represent a minor EpiSC fraction, resemble cells of the early epiblast, and can readily contribute to chimeras (Han et al., 2010). Our study of the ultrastructure of EpiSCs suggests that differentiated cells in EpiSCs might compromise the pluripotency potential of mouse primed ESCs.
Notably, when maintained on hAECs with LIF, ESD-EpiSCs could convert to naïve-like ESCs, with ultrastructural characteristics identical to mouse naïve ESCs, and less differentiated cells were observed in converted EpiSCs (cEpiSCs). However, further study of chimeras is necessary to prove the pluripotency potential of cEpiSCs.
In conclusion, this study showed that ultrastructural features of mouse primed ESCs are different from those of mouse naïve ESCs. However, converted primed ESCs were similar to mouse naïve ESCs, indicating that ultrastructure could be used to evaluate the pluripotency of ESCs.
Footnotes
Acknowledgments
We thank Professor Naihe Jing for providing the ESD-EpiSC cell line. This work was supported by grants from National Natural Science Foundation of China (NSFC) (grant nos. 81070533 and 81370678).
Author Disclosure Statement
The authors declare that there are no conflicts of interest.